高寒灌丛土壤温室气体释放对添加不同形态氮素的响应
2015-01-02马钢王平王冬雪徐世权
马钢,王平,王冬雪,徐世权
(1.甘肃农业大学资源与环境学院,甘肃 兰州730070;2.中国科学院成都山地灾害与环境研究所,四川 成都610041)
随着工业的发展,人类向大气中排放的含氮物质越来越多,导致大气氮沉降不断增加。通过长期氮沉降输入生态系统的氮素会改变土壤的碳氮比以及土壤理化环境,从而影响到土壤微生物群落结构组成以及土壤微生物活性,进而影响到生态系统土壤呼吸[1-3]。有研究指出,全球每年沉降到陆地的活性氮有43.7Tg N/a[4],人工施肥也导致各生态系统氮素输入量增加,这些已经对生态系统造成了不同程度的影响[5]。世界三大氮沉降集中区是中国、美国和欧洲,中国作为三大氮沉降集中区之一,氮沉降现象越来越严重,氮的排放量已由1961年的1.4×1010kg/a,升至2000年的6.8×1010kg/a,预计2030年活性氮排放量将达到1.05×1011kg/a[6]。当前,氮素输入引起的土壤碳氮生物地球循环的改变已受到广泛关注,国内外学者也相继展开了一系列有关氮沉降对土壤影响的研究[7-10],在这些研究中,氮沉降与土壤CO2、CH4和N2O等温室气体排放的关系研究是一项重要的主题,而且认识到氮沉降对不同生态系统温室气体释放的影响不同,有促进[11]、抑制[12]和无变化[13]等响应方式,因此还需要深入研究氮素输入对土壤呼吸的影响。我国有关氮沉降对土壤温室气体排放的研究开始较晚,目前研究对象主要集中在农田,森林和温带草原生态系统土壤上[14-16],而氮沉降对青藏高原高寒生态系统土壤温室气体释放影响的研究还比较薄弱。
陆地生态系统是全球碳循环重要组成部分,在全球碳收支平衡中占有主导地位,是预测气候变化的基础[17]。而青藏高原是世界上面积最大的高海拔陆地,高寒草地是其独特的植被类型,因其土壤碳储量较高,而且对气候变化敏感,在亚洲甚至全球气候变化中有着重要作用,是对全球变化最为敏感的地区之一[18]。青藏高原广泛分布着高寒草甸,高寒草原和高寒灌丛等高寒草地生态系统类型,其土壤是一个巨大的碳氮库,在区域乃至全球碳氮循环中有着重要的作用[19]。同时,由于高寒草地生态系统比较脆弱,其碳循环极易受到人类活动的干扰,并随着气候的变化而产生相应变化[20]。有研究表明,青藏高原东缘目前的氮沉降速度为8.7~13.8kg/(hm2·a)[21],已经对高寒草地生态系统产生了明显的影响。并且由于大气沉降加剧和其他人为活动,青藏高原正在经历氮素输入增加的影响,在未来几十年中氮素输入将呈现不断增加的趋势[22]。由此可见,未来高寒生态系统氮素输入对其碳氮循环可能产生显著影响,而认识和揭示高寒生态系统土壤温室气体对氮素添加的响应可以更好地预测全球变化条件下高寒生态系统土壤碳氮通量和存储量的变化。然而,目前有关氮素输入对高寒生态系统土壤呼吸的影响主要集中在高寒草甸和高寒草原上[23-25],且主要研究单一氮源(如NH4+)对土壤温室气体的排放的影响,截至目前,鲜有针对氮素输入对高寒灌丛生态系统土壤影响的报道。本文针对目前外源氮素输入对青藏高原高寒灌丛土壤影响研究不足的现状,对比分析了高寒灌丛土壤温室气体对不同形态氮素输入的响应差异,以期为全球变化背景下高寒生态系统土壤碳氮循环管理和评价提供科学依据。
1 材料与方法
1.1 研究区概况
研究地点位于四川省西北部,距离成都市约400km的青藏高原东部的若尔盖高原-红原县(33°54′N,102°36′E),东南部地势较高,西北部地势较低,研究区是一个相当平坦的宽谷景观,周围被高度50~150m的小山丘环绕,主要地貌类型为浅丘山地和丘间低地。当地海拔3500m以上,是严格的大陆性高原气候,日温差较大。年平均降水量752mm,5-9月占到全年降水量的86%左右,年均蒸发量1263mm,年平均湿度60%~70%,年均日照时间2159h,年均太阳辐射6194mJ/m2。年平均气温1.1℃,没有绝对无霜期。月平均气温最高的月份是7月,最高月平均气温约10.9℃;月平均气温最低的月份是1月,最低月平均气温约为-10.3℃。
试验样地植被为金露梅(Potentillafruticosa)高寒灌丛。植被盖度为60%~75%,金露梅灌丛高50~70 cm,丛间草本优势植物种为紫羊茅(Festucarubra)、矮嵩草(Kobresiahumilis)和垂穗披碱草(Elymsnutans)。土壤为暗沃寒冻雏形土,土层发育年轻,有机质含量高,其土壤基本性状见表1。

表1 研究区高寒灌丛土壤理化性质Table 1 Chemical and physical properties of the soils studied(alpine shrub)
1.2 样品采集和处理
2012年,选择典型的高寒灌丛生态系统,随机设置10m×10m样方5个,在每个样方内,用直径为5cm的土钻随机采集0~15cm表层土壤,每个样方取5~10个样点,每个样方土壤样品混合作为一个复合样品,得到20个复合土壤样品。采集土样前,用剪刀齐地面剪除植物地上部分,采集的土壤样品在室温下风干并剔除土样中残留的根茬等,然后过2mm筛,过筛后的土样充分混匀,储存在4℃下备用。
1.3 样品实验室培养
使用275mL玻璃广口瓶进行培养,广口瓶的橡胶塞上接三通阀。培养实验设置3个氮素添加处理和1个对照。3个添加处理的试剂分别为:NH4Cl,NH4NO3,KNO3,添加量为0.1g N/kg干土,对照即不做任何添加处理,每个处理设5个重复。培养时把30g风干土装入培养瓶中,分别添加试剂并添加去离子水调节土壤含水量至土壤持水能力(WHC)的60%,在15℃培养箱中黑暗条件下培养29d,培养过程中使用称重法差额补水,保持土壤含水量恒定。在培养第2,5,9,13,17,21,25,29天定时进行8次采气,采气样时塞上橡胶塞立刻使用气密注射器依次从每个培养瓶中抽取10mL气体样品,每个培养瓶采完气立刻关闭三通阀,密闭2h后再使用另一批气密注射器依次从每个培养瓶中抽取10mL气体样品,采样结束取下橡胶塞。所取气体样品均在24h内使用气相色谱(Agilent 7890A)测定温室气体浓度。在培养第2天和第29天,分别使用0.5mol/L K2SO4溶液提取土壤 NH4+-N、NO3--N、可溶性有机碳(DOC),并用流动分析仪(Auto Analyzer 3)分析其含量。
1.4 计算与统计分析
净硝化速率公式[26]:Nnit=(N2’-N1’)/d;其中,Nnit为净硝化速率(mg/L·d),N1’为培养前的硝态氮量(mg/L);N2’为培养后的硝态氮量(mg/L);d为培养天数。若为净铵化速率,则Nnit为净铵化率(mg/L·d),N1’为培养前的铵态氮量(mg/L);N2’为培养后的铵态氮量(mg/L)。
温室气体累积排放量计算公式参考王义祥等[27]测定生物呼吸累积量的方法,公式如下:
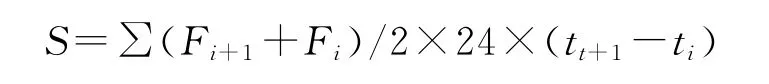
式中,S为温室气体(CO2,CH4和N2O)累积排放量,表示CH4时为累积吸收量;i为测定次数;t为测定时间,即培养天数。F是每次采样分析得出的温室气体排放速率(mg/kg·h),连续2次排放速率之和除以2,作为这2次采气间隔时间内的温室气体平均排放速率,平均值乘以每天的24h,再乘以2次采气的日期天数差,就得到这段时间内的温室气体累积排放量(mg/kg),连续求和得到整个实验过程中的温室气体总累积排放量。
N2O和CH4不同尺度上的增温效应采用2001年的IPCC标准,公式如下[28]:

式中,TH是积分时间范围,一般取20年,100年,500年;RFx和RFCO2分别表示温室气体x和参考气体CO2的辐射强迫;ax和aCO2分别表示温室气体x和参考气体CO2的辐射效率;x(t)和r(t)分别表示温室气体x和参考气体CO2的时间响应函数。计算得到在20年时间尺度上,单位质量N2O和CH4的全球增温潜能(GWP)分别为CO2的275和62倍;在100年时间尺度上,单位质量N2O和CH4的GWP分别为CO2的298和25倍;在500年时间尺度上,单位质量N2O和CH4的GWP分别为CO2的156和7倍。
数据运算和作图使用Microsoft Excel 2013。应用SPSS 17.0软件进行了数据的统计分析:采用方差分析和最小显著差异法(LSD)分析比较了不同处理间的差异,利用单变量分析确定不同处理的标准偏差(SD),运用SPSS相关分析进行了相关性分析。
2 结果与分析
2.1 土壤矿化态氮和DOC的变化
净铵化速率为正值表示培养期间土壤铵态氮增加,负值表示铵态氮减少(表2)。本研究中,CK和KNO3处理在培养期间净铵化速率均为0.67mg/(L·d),只添加铵态氮的NH4Cl处理净铵化速率最大,为1.07mg/(L·d),NH4NO3处理的净铵化速率是负值,为-0.21mg/(L·d)。NH4NO3处理铵态氮含量是减少的,培养后减少了5.76mg/L,与其他处理差异均达到显著水平(P<0.05),其他处理铵态氮含量都是增加的,且两两间差异不显著(P>0.05)。
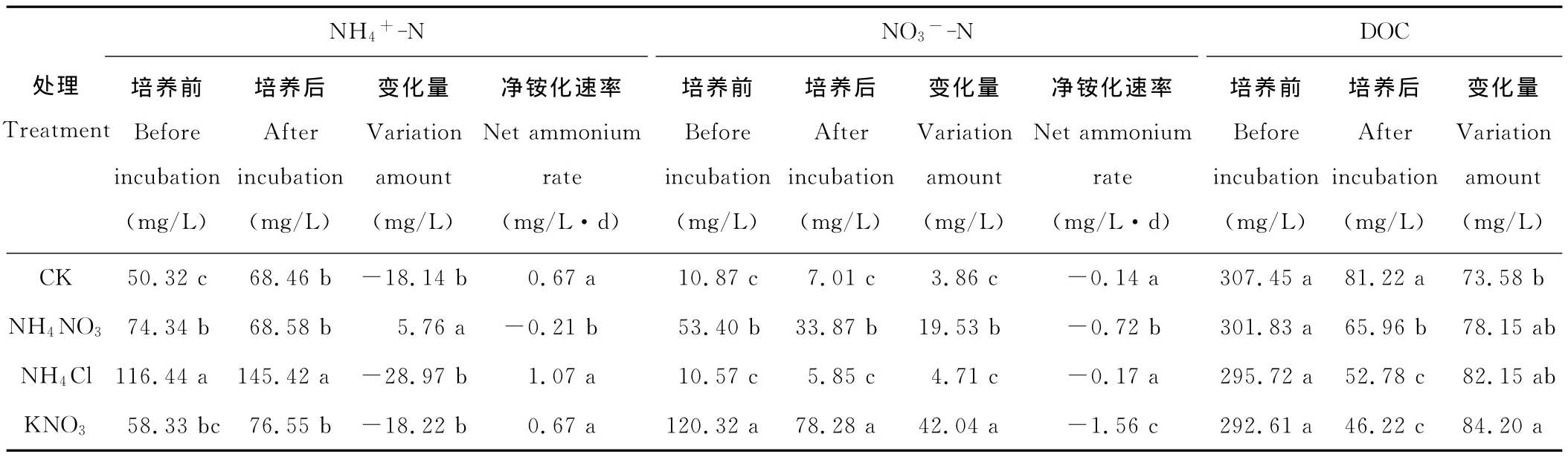
表2 培养前后土壤矿化态氮和可溶性有机碳变化Table 2 The variation of the mineral nitrogen and dissolved organic carbon(DOC)
净硝化速率为负值,表示土壤硝态氮减少。培养过程中,所有处理硝态氮含量均减少,其中CK净硝态氮减少速率为0.14mg/(L·d),NH4NO3处理为0.72mg/(L·d),NH4Cl处理为0.17mg/(L·d),KNO3处理为1.56mg/(L·d)。研究发现土壤添加硝态氮越多,硝态氮含量减少越快。CK和NH4Cl处理间的硝态氮减少速率差异不显著,其他两两处理间差异均显著(P<0.05)。各处理硝态氮减少速率大小趋势:KNO3>NH4NO3>NH4Cl>CK,与各处理N2O累积排放量一致。
DOC作为土壤微生物呼吸的底物,被土壤微生物呼吸作用分解,释放出CO2,经过培养之后各处理的土壤DOC含量均降低。CK和KNO3两个处理土壤DOC变化差异显著(P<0.05),其他两两处理间差异均不显著。经过培养,CK的土壤DOC含量减少了73.58%,与CK相比,施氮处理的土壤DOC减少更多,NH4NO3、NH4Cl和KNO3三个处理的土壤DOC分别减少了78.15%,82.15%和84.20%。
2.2 添加不同形态氮素对土壤CO2释放的影响
2.2.1 添加不同形态氮素对CO2排放速率和累积排放量的影响 研究发现,高寒灌丛土壤输入氮素,CO2释放被抑制,施氮处理的CO2累积排放量各不相同且均低于CK(图1)。培养期间,CO2排放速率一直呈减小趋势,第一次测定(培养第2天)时CO2排放速率最大,培养第2天到第5天CO2排放速率减小较快,培养5d后CO2排放速率减小较慢(图1)。
经过培养,施氮处理CO2累积排放量均显著低于CK(P<0.05)。CK的CO2累积排放量是1673.98 mg/kg,NH4NO3处理是1429.87mg/kg,KNO3处理是1379.87mg/kg,NH4Cl处理是1328.08mg/kg。与CK相比,NH4NO3处理的CO2累积排放量减少了14.58%,KNO3处理减少了17.57%,NH4Cl处理减少了20.66%。施氮处理中的NH4Cl处理与NH4NO3处理的CO2累积排放量差异达到显著水平(P<0.05),而KNO3处理和NH4Cl、NH4NO3处理间差异均不显著。

图1 不同处理CO2通量及累积排放量Fig.1 CO2fluxes and emission under different treatments
2.2.2 土壤DOC与CO2排放的关系 作为土壤微生物呼吸作用的底物,土壤DOC含量不同,土壤微生物呼吸释放的CO2量也各不相同。
高寒灌丛土壤的CO2累积排放量与土壤DOC浓度之间相关系数为0.7237,二者具有极显著的正相关关系(P<0.01)(图2)。说明土壤中DOC含量可能是衡量高寒灌丛土壤释放CO2能力的一个重要指标。本研究中添加氮素之后土壤DOC含量发生了一定程度的变化,可在一定程度上解释土壤释放CO2对氮素添加的响应模式。本研究表明至少在短期内氮素的添加会抑制高寒灌丛土壤CO2的释放,从而减少CO2排放量。推测无论未来氮沉降加剧还是人工施氮肥增多,青藏高原高寒灌丛土壤排向大气的CO2量都将减少。

图2 CO2累积排放量和土壤DOC含量的关系Fig.2 The relationship between CO2emission and the concentration of DOC
2.3 添加不同形态氮素对土壤CH4释放的影响
湿地是大气CH4的主要源,其他陆地生态系统则大多是大气CH4的汇,本研究也显示高寒灌丛土壤是大气CH4的汇。图3显示,只有在培养第2天测得的CH4通量是正值,培养第5天开始至培养结束,CH4通量均为负值。培养期间,CH4通量变化是单峰曲线,吸收峰出现在培养的第9天。土壤中CH4产生和CH4氧化作用共存,刚开始培养时,土壤理化性质在短时间内适合甲烷微生物生长,且底物充足,甲烷微生物释放的CH4量大于甲烷氧化量,使土壤短时间内成为CH4的源。培养第2天到第9天,随着底物浓度减少以及土壤理化性质变化,甲烷微生物活性受到抑制,CH4释放量越来越少,土壤吸收CH4不断增多。但表2显示培养过程中铵态氮是增加趋势,土壤中的铵态氮与土壤CH4氧化作用会产生竞争,培养第9天之后土壤中增加的铵态氮开始强烈抑制CH4的氧化作用,且培养瓶中甲烷减少也成为甲烷氧化作用的制约因素,因此土壤对CH4的吸收越来越少。
培养期间 CK 的 CH4累积吸收量为207.53μg/kg;NH4Cl处理为191.63μg/kg,比 CK 少7.66%;NH4NO3处理为196.40μg/kg,比CK少5.37%;KNO3处理为199.48μg/kg,比CK少3.88%。图3显示土壤对CH4的吸收有随着添加的铵态氮量增加而减小的趋势,但显著性分析显示不同处理间土壤对CH4的吸收差异不显著(图3)。

图3 土壤CH4排放速率及累积排放量Fig.3 CH4fluxes and emission under different treatments
2.4 添加不同形态氮素对土壤N2O释放的影响
2.4.1 添加不同形态氮素对N2O排放速率和累积排放量的影响 所有施氮处理的土壤N2O排放速率都经历了一个先增大后减小的过程(图4),N2O排放高峰出现在培养的第5天到第9天,且在N2O排放高峰期间施氮处理的土壤N2O排放速率显著高于CK(P<0.05)。KNO3处理的土壤N2O排放速率高于其他处理,添加有铵态氮的两个处理(NH4Cl和NH4NO3)的土壤N2O排放速率差异不显著。培养刚开始一段时间内N2O排放速率增大可能与土壤中作为微生物分解底物的铵态氮硝态氮含量较丰富有关,随着分解底物的减少和土壤理化性质的逐渐变化,从培养第9天开始,所有处理的N2O排放速率开始急剧减小,并在培养第13天后逐渐趋于稳定(图4)。

图4 培养土壤N2O排放速率及累积排放量Fig.4 N2O fluxes and emission under different treatments
图4显示所有施氮处理的N2O累积排放量均显著高于CK(P<0.05),且KNO3处理的N2O累积排放量显著高于NH4Cl处理和NH4NO3处理(P<0.05),但NH4Cl处理和NH4NO3处理间N2O累积排放量差异不显著。CK的 N2O累积排放量为2.64mg/kg,NH4Cl处理为6.39mg/kg,NH4NO3处理为7.66mg/kg,KNO3处理为10.16mg/kg。与CK相比,土壤添加NH4Cl,N2O累积排放量增加了1.42倍;添加NH4NO3,增加了1.90倍;添加KNO3,增加了2.85倍。本研究显示添加硝态氮越多,土壤排放N2O越多,可能与高寒灌丛土壤反硝化作用大于硝化作用有关,这与其他的研究结果一致[29]。
2.4.2 土壤硝态氮和铵态氮与N2O排放的关系 硝化作用和反硝化作用过程中均有N2O产生,为了确定高寒灌丛土壤释放出的N2O的主要来源,研究了N2O累积排放量和土壤硝态氮、铵态氮含量的关系(图5)。
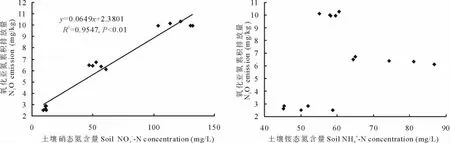
图5 N2O累积排放量和土壤硝态氮、铵态氮含量的关系Fig.5 The relationship between N2O emission and the concentration of NH4+-N or NO3- -N
图5显示,高寒灌丛土壤N2O累积排放量与土壤硝态氮含量的相关系数是0.9547,二者之间有极显著正相关关系(P<0.01),N2O累积排放量与土壤铵态氮含量无明显关系(P>0.05)。说明高寒灌丛土壤反硝化作用大于硝化作用,该土壤排放到大气中的N2O主要来自反硝化细菌对土壤中硝态氮的分解过程,在土壤硝态氮含量达到微生物分解极限之前,添加硝态氮越多,土壤释放的N2O将越多。
2.5 添加不同形态氮素释放温室气体的综合温室效应
若以CO2累积排放量1mg/kg土壤的GWP为1,可计算出添加不同形态氮素后土壤释放的温室气体在不同时间尺度上的综合增温潜能GWP(表3)。
表3显示,只有在500年时间尺度上的NH4Cl处理的GWP与CK差异不显著,其他施氮处理在不同时间尺度上的GWP均显著高于CK(P<0.05),同一时间尺度上,KNO3处理的GWP显著高于其他两个施氮处理(P<0.05),但NH4Cl处理和 NH4NO3处理的GWP在不同时间尺度上的差异均不显著。所有处理在100年时间尺度上的GWP最大,在500年时间尺度上的GWP最小。CK,NH4Cl,NH4NO3和KNO3各处理在100年时间尺度上的GWP较在20年时间尺度上的GWP分别增加了2.86%,5.01%,5.20%和5.79%,在500年时间尺度上的GWP较在100年时间尺度上的GWP则分别减少了15.10%,28.01%,29.23%和32.69%。在20年时间尺度上,NH4Cl,NH4NO3和KNO3各施氮处理的GWP分别是CK的1.29,1.48和1.74倍;在100年时间尺度上,分别是CK的1.32,1.51和1.79倍;在500年时间尺度上,分别是CK的1.12,1.26和1.42倍。在500年时间尺度上,NH4Cl处理的GWP与CK差异不显著,这可以为高寒灌丛土壤施肥提供一定的参考。

表3 不同处理释放温室气体的综合温室效应Table 3 Global warming potential(GWP)of different treatments CO2-eq mg/kg
3 讨论
氮素输入可能是导致土壤作为温室气体源和汇功能发生变化的一个重要原因[30]。现有研究表明,添加氮素可使温带和寒带土壤CO2释放量增加、减少或不变[11-13],本研究发现氮素添加抑制了高寒灌丛土壤CO2的排放。氮素对土壤释放CO2的抑制作用可能有以下解释:1)氮素添加抑制了与有机质分解有关的酶的数量和活性,降低了土壤微生物分解有机物释放CO2的能力和速率[31];2)土壤有效氮增加之后,土壤中的微生物可能转变呼吸方式以响应土壤有效氮含量的变化,从而减少了土壤CO2的排放[32]。DOC是土壤有机质物理、化学和生物分解的产物[33],DOC的分解与土壤理化性质和土壤微生物数量及活性有关。添加铵态氮和硝态氮都能抑制DOC分解酶的数量和活性[34],而添加铵态氮更可使土壤酸化,进而改变土壤微生物群落组成和微生物多样性指数[35],从而抑制土壤DOC的分解,因此单施铵态氮比单施硝态氮对DOC分解的抑制作用更大,导致NH4Cl处理的土壤释放出的CO2最少。同时添加两种形态氮素可能引起土壤理化性质和土壤微生物更复杂的变化,这可能是NH4NO3处理的土壤CO2排放量高于其他两个施氮处理的原因。有人推测土壤释放出的CO2的主要来源是土壤中的DOC[36],也有报道指出土壤呼吸和DOC浓度具有正相关性[33]。本研究发现,培养期间CO2累积排放量与土壤DOC含量有显著正相关关系,与已有研究一致[33]。总体上来看,高寒灌丛土壤CO2释放对外源氮素的输入表现为负反馈效应。
外源氮素输入也影响土壤对CH4的排放和吸收。氮素添加对土壤CH4排放的影响,不同研究得出的结论不一致。有研究认为土壤中添加铵态氮可抑制甲烷产生菌的活性或促进CH4的氧化[37],也有观测发现,施氮处理比不施氮处理的土壤CH4排放量会增加10%~20%[38]。一般情况下,只有在极端还原条件下(氧化还原电位低于-150~-160mV),土壤中甲烷微生物才能利用CO2和H2合成或分解乙酸等生成甲烷[30],因此湿地是大气CH4的一个很重要的源。除湿地外,大多陆地生态系统为好气的土壤环境,有利于甲烷营养微生物的生长,土壤吸收和氧化甲烷的作用强烈,使之成为大气甲烷的一个重要汇。本研究也证明了高寒灌丛土壤是大气CH4的汇。目前,施用氮肥抑制土壤对CH4吸收的机理还不十分清楚,但有研究表明,NH4+可作为CH4替代物而被许多甲烷营养微生物利用[30],可能是施氮肥增加了土壤NH4+含量,NH4+与CH4产生了竞争,从而减少了甲烷营养微生物对CH4的吸收。本研究虽然发现所有施氮处理对CH4的吸收低于CK,且随着添加的铵态氮的增多,土壤对CH4的吸收有减少趋势,但各施氮处理对CH4的累积吸收量较CK的差异均不显著,即高寒灌丛土壤对CH4的吸收与添加不同形态氮素无显著关系。
土壤N2O是微生物参与分解的硝化、反硝化作用的产物,土壤的硝化作用与反硝化作用是大气中N2O的主要产生机制[39]。硝化作用是指在通气条件下,土壤中硝化微生物将铵盐转化为硝酸盐的过程,可释放N2O[40]。反硝化过程是指在通气不良条件下,土壤微生物将硝酸盐或硝态氮还原成氮气(N2)或氧化氮(N2O,NO)的过程[40]。由于不同土壤类型中硝化细菌、亚硝化细菌和反硝化细菌群落结构组成和数量不同,硝化反硝化作用也各不相同。有研究表明,铵化速率受土壤pH影响很大[41],从化学角度来看,KNO3作为中性肥料,不会改变土壤pH值,因此KNO3处理的净铵化速率与CK一致。NH4Cl和NH4NO3可不同程度改变土壤理化性质,从而影响到土壤微生物,使二者净铵化速率不一致。有研究表明,高寒灌丛土壤中反硝化细菌比硝化细菌多,反硝化作用大于硝化作用[29],因此高寒灌丛土壤中硝态氮越多,释放出的N2O越多,二者有正相关关系。
本研究发现所有处理在100年时间尺度上的GWP最大,在500年时间尺度上的GWP最小,这是因为CO2在大气中可平均存留200年左右,而N2O可平均存留150年左右,只有在长期条件下,由于自然分解破坏机制的存在,GWP才会被削弱。本研究发现在同一时间尺度上,添加硝态氮越多,GWP越大,可能也与高寒灌丛土壤微生物反硝化作用大于硝化作用有关。
4 结论
研究表明,氮素添加会抑制高寒灌丛土壤CO2排放。CO2累积排放量与土壤DOC浓度显著正相关。施氮处理的土壤对CH4的吸收低于CK,但较CK差异不显著,不同形态氮素间差异也不显著。高寒灌丛土壤微生物反硝化作用大于硝化作用,释放的N2O主要来自土壤微生物反硝化作用过程。土壤硝态氮越多,N2O排放量越高,二者呈显著正相关关系。在不同时间尺度上,施氮处理的GWP均显著高于CK,但NH4Cl处理和NH4NO3处理的GWP差异不显著。在同一时间尺度上,KNO3处理的GWP显著高于其他处理。不同处理温室气体全球增温潜能大小:KNO3>NH4NO3>NH4Cl>CK。
青藏高原高寒生态系统经历着氮沉降的影响,本研究证明了高寒灌丛土壤温室气体排放对不同形态氮素沉降有不同的响应,在评价氮沉降对这一土壤温室气体排放的影响时,应充分考虑不同形态氮素输入所产生的差异性。过度放牧、鼠虫灾害以及全球变暖可使草原退化、产草量下降,因此在未来,施氮可能将是高寒生态系统管理的主要措施之一,制定氮肥使用措施时,应考虑施肥后土壤释放的温室气体的综合温室效应。从减少温室气体排放的角度来考虑,在高寒灌丛生态系统氮肥的选择上,应该推荐使用铵态氮肥。
Reference:
[1]Peng Q,Dong Y S,Qi Y C.Influence of external nitrogen input on key processes of carbon cycle in terrestrial ecosystem.Advances in Earth Science,2008,23(8):874-883.
[2]Liu D Y,Song C C.Influence of external nitrogen input on soil organic carbon mineralization and litter decomposition.Chinese Journal of Soil Science,2008,39(3):675-680.
[3]Zhang Y L,Zhang J,Shen Q R,etal.Effect of combined application of bioorganic manure and inorganic nitrogen fertilizer on soil nitrogen supplying characteristics.Chinese Journal of Applied Ecology,2002,13(12):1575-1578.
[4]James N,Galloway,Ellis B,etal.Reactive nitrogen and the World:200years of change.AMBIO-Journal of Human Environment,2002,(2):64-71.
[5]Galloway J N,Aber J D,Erisman J W,etal.The nitrogen cascade.Bioscience,2003,53(4):341-356.
[6]Zheng X H,Fu C B,Xu X K,etal.The asian nitrogen cycle case study.AMBIO-Journal of Human Environment,2002,(2):79-87.
[7]Tietema A,Wright R F,Blank K,etal.NITREX:The timing of response of coniferous forest ecosystems to experimentally changed deposition of nitrogen.Water,Air and Soil Pollution,1995,85:1623-1628.
[8]Wright R F,Roelofs J G M,Bredemeier M,etal.NITREX:Responses of coniferous forest ecosystems to experimentally changed deposition of nitrogen.Forest Ecology and Management,1995,71:163-169.
[9]Aber J D,McDowell W H,Nadelhoffer K J,etal.Nitrogen saturation in temperate forest ecosystems:hypotheses revisited.Bio Science,1998,48:921-934.
[10]Flanagan P W,Van C K.Nutrient cycling in relation to decomposition and organic-matter quality in taiga ecosystems.Canadian Journal of Forest Research,1983,13(5):795-817.
[11]Song X G,Hu T X,Xian J R,etal.Soil respiration and its response to simulated nitrogen deposition in evergreen broad-leaved forest,southwest Sichuan.Journal of Soil and Water Conservation,2007,(4):168-172.
[12]Bowden R D,Davidson E,Savage K,etal.Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest.Forest Ecology and Management,2004,196(1):43-56.
[13]Micks P,Aber J D,Boone R D,etal.Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests.Forest Ecology and Management,2004,196(1):57-70.
[14]Mo J M,Fang Y T,Xu G L,etal.The short-term responses of soil CO2emission and CH4uptake to simulated N deposition in nursery and forests of Dinghushan in subtropical China.Acta Ecologica Sinica,2005,(4):682-690.
[15]Hu Z H,Li H M,Yang Y P,etal.Effects of simulated nitrogen deposition impact on soil respiration in the northern subtropical deciduous forest.Chinese Journal of Environmental Science,2010,(8):1726-1732.
[16]Li R H,Tu L H,Hu T X,etal.Effects of simulated nitrogen deposition on soil respiration in a Neosinocalamus affinis plantation in Rainy Area of West China.Chinese Journal of Applied Ecology,2010,(7):1649-1655.
[17]Ren J Z,Lin H L.Grassland soil organic carbon storage technology research.Acta Prataculturae Sinica,2013,(6):280-294.
[18]Wang G X,Cheng G D,Shen Y P.Soil organic carbon pool of grasslands on the Tibetan Plateau and its global implication.Journal of Glaciology and Geocryology,2002,(6):693-700.
[19]Wang G,Qian J,Cheng G,etal.Soil organic carbon pool of grassland soils on the Qinghai-Tibetan Plateau and its global implication.Science of the Total Environment,2002,91:207-217.
[20]Chen W Y,Zhang J,Qi D C,etal.Desertification dynamic change trend and quantitative analysis of driving factors of alpine meadow in Maqu County in the First Meander of the Yellow River.Acta Prataculturae Sinica,2013,(2):11-21.
[21]LüC Q,Tian H Q.Spatial and temporal patterns of nitrogen deposition in china:Synthesis of observational data.Journal of Geophysical Research,2007,112(D22):1-10.
[22]Galloway J N,Dentener F J,Capone D G,etal.Nitrogen cycles:past,present,and future.Biogeochemistry,2004,70(2):153-226.
[23]Zhu T H,Cheng S L,Fang H J,etal.Early responses of soil CO2emission to simulating atmospheric nitrogen deposition in an alpine meadow on the Qinghai Tibetan Plateau.Acta Ecologica Sinica,2011,10:2687-2696.
[24]Bai J B,Xu X L,Song M H,etal.Effect of temperature and nitrogen input on soil carbon mineralization in three alpine grassland on the Tibetan Plateau.Ecology and Environmental Sciences,2011,(5):855-859.
[25]Zhang P L,Fang H J,Cheng S L,etal.The early effects of nitrogen addition on CH4uptake in an alpine meadow soil on the Eastern Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2013,13:4101-4110.
[26]Gao J Q,Ou Y H,Zhang F,etal.The response of soil nitrogen mineralization to temperature and water moisture in Zoige alpine wetland.Wetland Science,2008,(2):229-234.
[27]Wang Y X,Wang F,Wong B Q,etal.The effect of grass cultivation on soil organic carbon mineralization in Amygdalus persica garden.Acta Prataculturae Sinica,2013,(6):86-92.
[28]Houghton J T,Meirafilho L G,Callender B A,etal.IPCC,Climate change 1995:The scientific of climate change[R].Cambridge,UK:Cambridge University Press,1996.
[29]Wang Z M,Le Y Z,Chen W M.The research of alpine meadow soil ammonia volatilization.Alpine Meadow Ecosystem,1993,(3):219-226.
[30]Sun W T,Xiao Q M,Lou C R,etal.The formation,emission and influencing factors of CH4in the soil.Rain Fed Crops,2000,(5):44-47.
[31]Lee K,Jose S.Soil respiration,fine root production,and microbial biomass in cottonwood and loblolly pine plantations along a nitrogen fertilization gradient.Forest Ecology and Management,2003,185:263-273.
[32]Moscatelli M C,Lagomarsino A,De Angelis P,etal.Seasonality of soil biological properties in a poplar plantation growing under elevated atmospheric CO2.Applied Soil Ecology,2005,30:162-173.
[33]Fang C M,Moncrieff J B.The variation of soil microbial respiration with depth in relation to soil carbon composition.Plant and Soil,2005,268:247-253.
[34]Compton J,Watrud L S,Porteus L A,etal.Response of soil microbial biomass and community composition to chronic nitrogen additions at Harvard forest.Forest Ecology and Management,2004,196:143-158.
[35]Xiao H L.Effects of atmospheric nitrogen deposition on forest soil acidification.Scientia Silvae Sinicae,2001,37(4):111-116.
[36]Bengtson P,Bengtsson G.Rapid turnover of DOC in temperate forests accounts for increased CO2production at elevated temperatures.Ecology Letters,2007,10:783-790.
[37]Cai Z C,Xing G X,Yan X Y,etal.Methane and nitrous oxide emissions from rice paddy fields as affected by nitrogen fertilizers and water management.Plant and Soil,1997,196(1):7-14.
[38]Li L,Hu L F,Chen F,etal.Effects of different long-term fertilization on emission of CH4and N2O from paddy soil.Journal of Agro-Environment Science,2006,25(Supplement):707-710.
[39]Qi Y C,Dong Y S.The formation,emission and influencing factors of N2O in the soil.Acta Geographica Sinica,1999,(6):534-542.
[40]Song W Z,Wang S B,Su W H,etal.The research of main greenhouse gases(CO2,CH4and N2O )emissions in Chinese farmland.Chinese Journal of Environmental Science,1996,(1):85-88.
[41]Zheng X Q.The preliminary of ammonification and nitrification changes in three kinds of farmland soil under different hydrothermal conditions[D].Nanjing:Nanjing Agricultural University,2008.
[1]彭琴,董云社,齐玉春.氮输入对陆地生态系统碳循环关键过程的影响.地球科学进展,2008,23(8):874-883.
[2]刘德燕,宋长春.外源氮输入对土壤有机碳矿化和调落物分解的影响.土壤通报,2008,39(3):675-680.
[3]张亚丽,张娟,沈其荣,等.秸秆生物有机肥的施用对土壤供氮能力的影响.应用生态学报,2002,13(12):1575-1578.
[4]James N,Galloway,Ellis B,etal.活性氮和世界:200年的变化.AMBIO-人类环境杂志,2002,(2):64-71.
[6]郑循华,符聪斌,徐星凯,等.亚洲氮循环案例研究.AMBIO-人类环境杂志,2002,(2):79-87.
[11]宋学贵,胡庭兴,鲜骏仁,等.川西南常绿阔叶林土壤呼吸及其对氮沉降的响应.水土保持学报,2007,(4):168-172.
[14]莫江明,方运霆,徐国良,等.鼎湖山苗圃和主要森林土壤CO2排放和CH4吸收对模拟N沉降的短期响应.生态学报,2005,(4):682-690.
[15]胡正华,李涵茂,杨艳萍,等.模拟氮沉降对北亚热带落叶阔叶林土壤呼吸的影响.环境科学,2010,(8):1726-1732.
[16]李仁洪,涂利华,胡庭兴,等.模拟氮沉降对华西雨屏区慈竹林土壤呼吸的影响.应用生态学报,2010,(7):1649-1655.
[17]任继周,林慧龙.草地土壤有机碳储量模拟技术研究.草业学报,2013,(6):280-294.
[18]王根绪,程国栋,沈永平.青藏高原草地土壤有机碳库及其全球意义.冰川冻土,2002,(6):693-700.
[20]陈文业,张瑾,戚登臣,等.黄河首曲-玛曲县高寒草甸沙化动态演变趋势及其驱动因子定量分析.草业学报,2013,(2):11-21.
[23]朱天鸿,程淑兰,方华军,等.青藏高原高寒草甸土壤CO2排放对模拟氮沉降的早期响应.生态学报,2011,10:2687-2696.
[24]白洁冰,徐兴良,宋明华,等.温度和氮素输入对青藏高原三种高寒草地土壤碳矿化的影响.生态环境学报,2011,(5):855-859.
[25]张裴雷,方华军,程淑兰,等.增氮对青藏高原东缘高寒草甸土壤甲烷吸收的早期影响.生态学报,2013,13:4101-4110.
[26]高俊琴,欧阳华,张锋,等.若尔盖高寒湿地土壤氮矿化对温度和湿度的响应.湿地科学,2008,(2):229-234.
[27]王义祥,王峰,翁伯琦,等.生草栽培对油桃园土壤有机碳矿化的影响.草业学报,2013,22(6):86-92.
[29]王在摸,乐炎舟,陈伟民.高山草甸土氨挥发的研究.高寒草甸生态系统,1993,(3):219-226.
[30]孙文涛,肖千明,娄春荣,等.土壤中甲烷的形成、排放及影响因素.杂粮作物,2000,(5):44-47.
[35]肖辉林.大气氮沉降对森林土壤酸化的影响.林业科学,2001,37(4):111-116.
[38]李琳,胡立峰,陈阜,等.长期不同施肥类型对稻田甲烷和氧化亚氮排放速率的影响.农业环境科学学报,2006,25(增刊):707-710.
[39]齐玉春,董云社.土壤氧化亚氮产生、排放及其影响因素.地理学报,1999,(6):534-542.
[40]宋文质,王少彬,苏维瀚,等.我国农田土壤的主要温室气体CO2、CH4和N2O排放研究.环境科学,1996,(1):85-88.
[41]郑宪清.不同水热条件下三种农田土壤中氨化和硝化作用的变化初探[D].南京:南京农业大学,2008.
