祁连山北坡中段祁连圆柏群落学特征的研究
2015-01-02田晓萍占玉芳
田晓萍,马 力,占玉芳
(张掖市林业科学研究院,甘肃 张掖734000)
祁连圆柏(Sabina przewalskii)是我国特有常绿针叶树,适应幅度广,喜光,耐寒、干旱和瘠薄,根系发达,抗风力强[1]。在祁连山林区,以它为建群种的天然林主要分布在海拔2 500~3 500m的阳坡、半阳坡上,伴生树种少,天然更新十分困难[2]。祁连圆柏作为祁连山林区亚高山暗针叶林带中惟一分布在阳坡或半阳坡上的森林类型[3],在寒区和旱区水土保持和水源涵养方面起重要的作用[4],具有其他针叶树种不可替代的生态价值,对构成祁连山森林复合群体的作用十分突出[1,3]。目前对祁连圆柏的研究多集中在该乔木树种本身的化学成分[5-7]、生物量及营养元素积累量[8]、育苗技术[9-11]、造林技术[12-13],以及年轮和气候等方面[14-16]。在群落学特征方面,对岷江上游祁连圆柏群落的研究较多[17-20],而对祁连山北坡中段祁连圆柏群落学特征方面的研究还尚未见报道。以祁连山北坡中段寺大隆林区的样地调查为基础,分析祁连圆柏群落的区系成分、层次结构、多样性和种群结构,为该群落的保护及祁连山林区特别是阳坡的植被恢复提供基础数据和科学依据。
1 研究区自然概况
研究地点位于祁连山北坡中段寺大隆林场向阳台护林站,38°26′304″N、99°55′707″E,海拔高度2 780m,温带大陆性气候,年日照时数2 200h,≥10℃的积温1 630.9℃;年平均气温3.6℃,一月平均气温-10.4℃,七月平均气温15.9℃,年均降水量约600mm,年蒸发量1 828.5mm;土壤主要为草甸土、山地棕壤土。
2 研究方法
2.1 样地设置与调查
在祁连圆柏自然分布较集中的祁连山寺大隆林区,选择分布较多的地段作为调查样地,分别在2 800、3 000m 和3 200m的海拔梯度各设置样地3个,共设置9个,样地面积20m×20m,每个样地取4个4m×4m灌木样方,并在每个灌木样方中各取1个1m×1m的草本样方。对样地内的乔木进行每木调查,详细记录种名、株数、胸径、树高等。记录灌木及草本植物种类、高度、株数(多度)、盖度等。同时,记录样地的群落总盖度、乔木层盖度、灌木层盖度、海拔、坡度、坡向、地理位置、经纬度、土壤类型及人类活动的干扰情况(表1)。在对9个样地作全面调查的基础上,根据海拔高度把样地1、2、3作为一个样地组,记为样地组1,同样把样地4、5、6记为样地组2,把样地7、8、9记为样地组3,分别对其指标进行平均和综和,便于比较。

表1 9个样地的基本情况Table 1 Basic conditions of 9sampling polts
2.2 区系组成与地理成分分析
将从样地采集的植物标本带回进行鉴定,分析其区系组成。按照吴征镒先生对中国种子植物分布区类型的划分方法[21-22],对群落种子植物进行地理成分分析。
2.3 多样性分析
2.3.1 重要值计算 重要值=(相对密度+相对频度+相对盖度)×100/3
2.3.2 物种多样性指数[23-24]采用的物种多样性测度指标为:物种多样性指数、物种丰富度指数、物种均匀度指数。
1)多样性指数
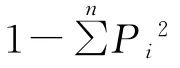
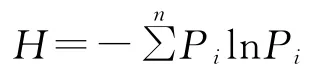
式中:Pi为ni/N ,是第i个种的株数占所有种株数的比例;N为所有种的个体总数。
2)Margalef丰富度指数(Ma):Ma=(S-1)/lnN
式中:S为群落中物种的数量。
2.4 种群结构划分
由于祁连圆柏生长缓慢,木材坚硬,加上调查区内株数较多,无法完整获取其年龄数据,鉴于多数乔木树种的胸径与年龄具有较好的相关性[25-26],并根据陈文年[18]等对祁连圆柏径级结构的划分情况,结合调查数据的特点,将径级共划分10级。划分方式:1)把胸径<2.5cm、树高<1.5m的幼树和幼苗,统一按高度分为3级:Ⅰ级高1~30cm,Ⅱ级高30~60cm,Ⅲ级高60~150cm;2)胸径2.5cm以上的个体按胸径级进行划分,以5cm为径阶划分出7个径级,Ⅳ级为2.5~7.5cm、Ⅴ级为7.5~12.5 cm、Ⅵ级为12.5~17.5cm、Ⅶ级为17.5~22.5 cm、Ⅷ级为22.5~27.5cm,Ⅸ级为27.5~32.5cm,Ⅹ级为32.5cm以上的大树。
3 结果与分析
3.1 群落区系组成与地理成分
经调查统计,祁连圆柏群落共有种子植物18科35属44种。其中裸子植物1科1属1种;双子叶植物15科28属36种;单子叶植物2科6属7种,无蕨类植物。群落植物含种数最多的是菊科(As-teraceae)6种,其次是禾本科(Gramineae)、唇形科(Labiatae)和毛茛科(Ranunculaceae)均4种,莎草科(Cyperaceae)、豆科(Leguminosae)、蔷薇科(Rosaceae)、玄参科(Scrophulariaceae)均3种,以上8科占总科数的44%;其余组成中含2种的有2科,占总科数的12%;仅占1种的有8科,占总科数的44%。在属的组成中仅含1种的有29属,占总属数的83%,显示出含1种的属比例较高的区系特点。由此可见,祁连圆柏群落植物的科、属组成分散,与种的数目相比,科、属的数目相对较多,尤其是属的组成最突出,这一现象说明祁连圆柏群落属于比较古老的植物区系。
对样地内18科35属种子植物进行地理成分分析(表2)。在科的分布类型中,世界分布的有11科;北温带分布的有2科,占总科数的34.58%;北温带和南温带间断分布“全温带”的有2科,占总科数的10.53%。在35属植物中,温带性质的属有22个,占总属数的62.86%,其中主要以北温带分布及其变型的属为主,共有17属,占总属数的48.57%,旧世界温带分布及温带亚洲分布的属均2个,其余分布类型的属各有1个。扣除世界分布属,属的分布类型表明祁连圆柏群落具有明显的温带性质。

表2 祁连圆柏群落种子植物分布区类型Table 2 Distribution types of seed plants in S.przewalskii community
3.2 群落的层次结构
森林群落的垂直结构是指群落的成层现象。层次结构是群落垂直结构的重要标志,其成因决定于生态环境,特别是群落生长的水热条件和土壤条件。该区祁连圆柏林是一个稀疏的植物群落,群落结构单一,垂直结构可分为乔木层、灌木层和草本层,无藤本植物及附生植物等层间植物。乔木层构成了群落外貌的主体部分。
表3为祁连山北坡中段祁连圆柏群落中各层物种重要值。可以看出:乔木层由单种祁连圆柏构成,该种是整个群落的建群种,在整个群落中其重要值最大,为35.30,占绝对优势。灌木层树种组成较简单,在调查的9个样地中仅有灌木3种,分别为银露梅(Potentilla glabra)、金露梅(Potentilla fruticosa)和怀腺柳(Salix cupularis)。其中银露梅的重要值最大,达到3.31,是祁连圆柏群落灌木层的优势种。金露梅的重要值为1.46,是该层的次优势种。怀腺柳的重要值最小,为0.28。草本层种类40种,其中重要值较大的前18种列于表3。从表3可以看到,矮生嵩草(Kobresia humilis)的重要值最大,达16.91,是该群落草本层的优势种,圆穗蓼(Polygonum macrophyllum)、牛 毛 毡 (Eleocharis yokoscensis)、鹅绒委陵菜(Potentilla anserina)、滨麦(Leymus mollis)草玉梅(Anemone rivularis )5种的重要值都在2.0以上,是次优势种。另外,在调查中还发现某些样方中密生苔草(Carex crebra)也可以形成次优势种。
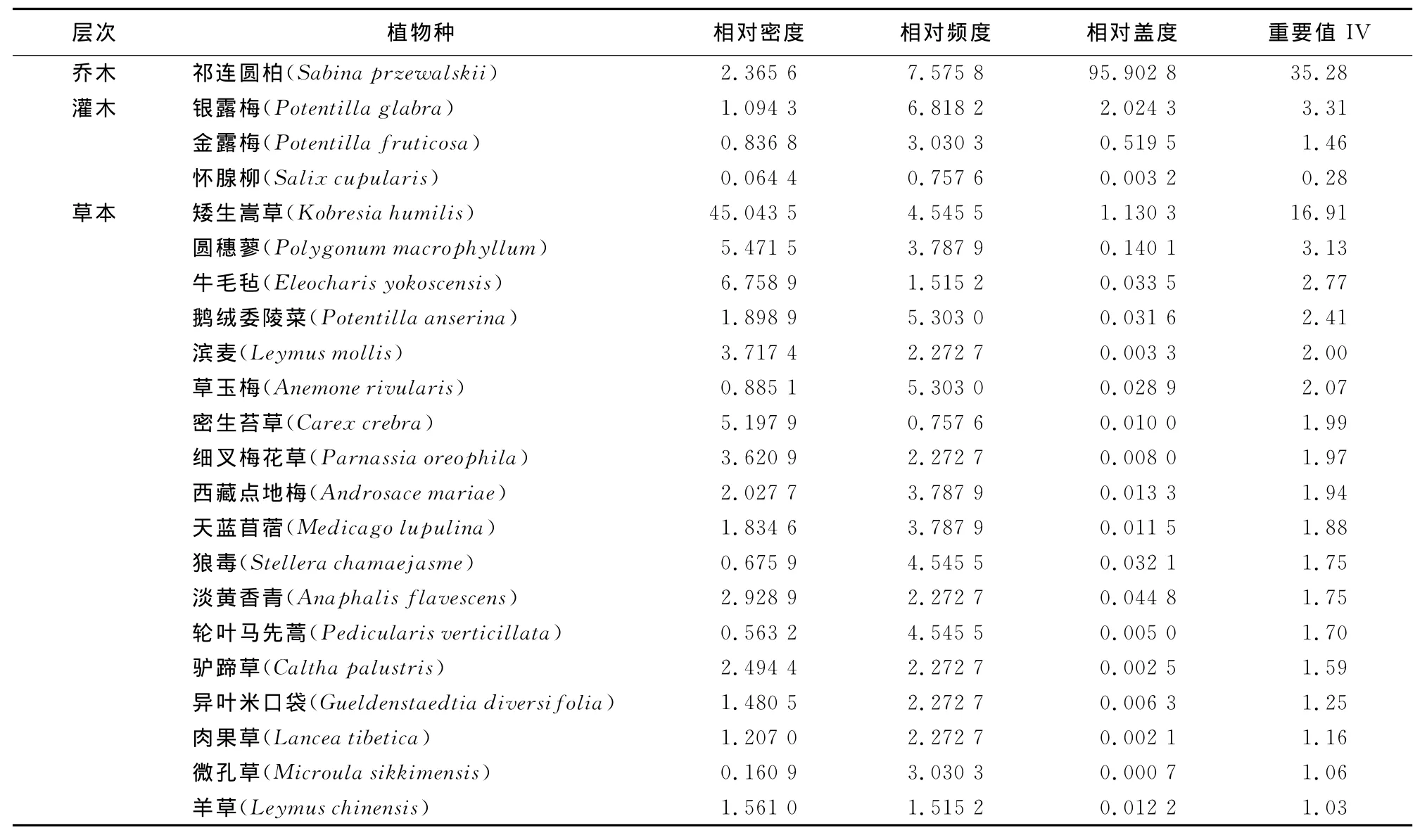
表3 祁连圆柏群落乔灌草层物种的重要值Table 3 Tree,shrub and herb layer species in S.przewalskii community and their importance value
从3个样地组的情况来看,乔木层均由单种祁连圆柏构成,是整个群落的建群种;灌木层物种组成不同,样地组1中有金露梅、银露梅和怀腺柳3种,样地组2中有金露梅、银露梅2种,而样地组3只有银露梅1种。草本层物种种类组成相差不大,但某些物种的重要值变化较大,在样地组1和2中矮生嵩草的重要值较大,特别是在样地组1中,达到了27.2,而在样地组3中只有7.7;对牲畜适口性差的轮叶马先蒿在样地组1中为0.77,而在样地组3中为2.75。3个样地组中草本层的物种组成及某些物种重要值的差异主要与3个样地的海拔高度不同导致人为干扰强度差异有关。
3.3 群落的多样性特征
祁连圆柏群落多位于祁连山自然保护区的阳坡地带,特殊的环境条件使该群落形成了自己独特的物种多样性特征。祁连山圆柏群落结构比较简单,乔木层只有祁连圆柏1种,盖度小,无层间植物,因此计算物种多样性时只考虑灌木层与草本层。

表4 祁连圆柏群落物种多样性指数Table 4 The species diversity indices in S.przewalskii community
祁连圆柏群落灌木层的物种数、Margalef物种丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数均小于草本层,说明草本层具有物种多样性、均匀度高的特点,但草本层的优势度指数(0.21)小于灌木层的(0.46)(表4)。这是由于在一个植物群落中不同的植物之间会出现以食物、空间等资源为核心的种间、种内竞争及密度制约因素,导致竞争性弱的物种比例减少,从而引起群落的均匀度降低。当群落由于密度增加引起优势度的增加而成为单优势种群落时群落多样性降低,而优势度增加[27]。从3个样地组的情况来看,由于乔木层、灌木层物种组成简单,因此对各样地组群落草本层的物种多样性进行了比较,样地组1草木层的物种数、Margalef物种丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数均最大,其次是样地组2,样地组3的最小,但不同样地组草本层物种多样性指数之间的差异不显著。
3.4 种群结构特征
种群结构是群落结构的基础,它在很大程度上影响着群落结构。对9个样地的数据进行综合,得到祁连圆柏种群径级结构图(图1)。从图1可以看出,各径级中Ⅵ、Ⅶ级树的个体较多,占总株数的40.1%,Ⅴ级以上的各级株数之和占总株数的72.8%,这说明种群中成年个体占有较大比重;Ⅴ级以下幼苗数急剧减少,Ⅰ、Ⅱ、Ⅲ级幼苗总数仅占总株数的10.2%,种群中幼树和幼苗稀缺,幼苗贮备严重不足,说明种群自身更新不良。从图1还可以看出,Ⅲ级幼苗所占的比例最小,说明Ⅰ、Ⅱ级幼苗的死亡率很高,这对于种群的发展十分不利。从整个径级结构图来看,祁连圆柏种群呈倒金字塔形,更新受阻导致种群结构严重失调,属衰退型种群。
图2表明,3个样地组中祁连圆柏种群的个体数随着径级的变化其变化趋势一致,均呈倒金字塔形。Ⅰ~Ⅲ级幼苗在3个样地组中所占的比例均不同,样地组1最高,为14.3%;样地组3最低,仅有5.1%,这可能与样地之间的海拔高度有较大的关系,由于样地组1海拔最高,放牧、砍薪等人为干扰作用较弱,较多幼苗得以保存下来。样地组3中祁连圆柏种群的径级结构不完整,未见Ⅰ径级的个体分布,主要是因为该样地组海拔最低,遭受的放牧、砍薪等人为干扰作用最强,牲畜的践踏、啃食对幼苗小树具有极大的破坏作用。
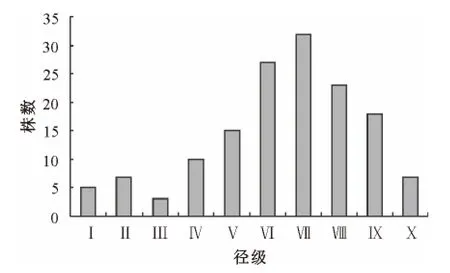
图1 祁连圆柏种群径级结构Fig.1 Diameter class structure of Sabina przewalskii populations

图2 样地组1(A)、2(B)、3(C)的径级结构Fig.2 Diameter class structure in frist,second and third plot groups
4 结论与讨论
祁连山北坡中段祁连圆柏群落组成种类较丰富,共记载有种子植物18科35属44种,温带性分布的科、属所占的比例较大,反映出该植物区系具有明显的温带性质,说明祁连圆柏群落属于1个比较古老的植物区系。群落垂直结构明显,可分为乔木层、灌木层、草本层,无藤本植物及附生植物等层间植物,乔木层由单种祁连圆柏构成,灌木及草本层的优势种分别是银露梅和矮生嵩草。群落中乔木层、灌木层物种多样性低于草本层,主要是因为祁连山林区特定的水、热条件的限制,尤其是春夏持续少雨干旱的状况影响乔灌木的生长,并可导致抗性弱的物种死亡。同时,人为采樵、砍伐和牛羊的啃食、践踏也抑制了乔灌木的更新与生长。另外,祁连圆柏群落草本层的多样性程度高,跟乔木层和灌木层的稀疏有关,上层乔灌木稀疏使阳光可以直接照射到草本层,从而促进了草本植物的生长。
种群结构分析表明,祁连圆柏种群的径级基本连续,结构呈倒金字塔形,该种群中主要分布中等偏大径级树木,小径级的幼苗和幼树比较少,并且Ⅰ、Ⅱ级幼苗的死亡率很高,属衰退型种群。另外,从不同海拔高度的样地来看,海拔低而放牧、砍薪等干扰作用较强的样地,其幼苗数量相对更少甚至缺失,更新更加困难。因为祁连圆柏种子产量低、质量差,种仁率只有10%~20%,并且后熟期长,一般播种后需2a才能出苗,繁殖困难,从而导致祁连圆柏种群更新不良。同时,由于柏木具有特殊的香味,材质坚硬,不易腐烂,是当地人们烧烤食品和制作农具、模具及棺椁的首选木材,畜牧业又是当地的支柱产业,林下植被成为家畜饲料的主要来源,当地牧民在林区放牧和砍伐,祁连圆柏实生幼苗连同周围的杂草被刈割、践踏,导致祁连圆柏固有的繁育体系遭到破坏,因此,林牧矛盾问题是导致祁连山林区祁连圆柏种群内幼苗稀缺、种群更新困难的又一原因。另外,祁连山林区特定的水、热条件限制,尤其是春夏持续少雨干旱的状况影响了祁连圆柏幼苗的萌发和生长。整个祁连圆柏更新进程和更新效果主要受立地条件、林分状况、植被状况等多种因子的综合作用和制约[1],其具体衰退机理和过程还需要进一步分析和研究。
祁连圆柏作为祁连山林区亚高山暗针叶林带中惟一分布在阳坡或半阳坡上的森林类型,是祁连山重要的水源涵养林,对保护阳坡水土流失具有重要作用。目前对祁连圆柏种群的保护力度还不够,为有效地保护该种群,应采取就地保护与迁地保护相结合的综合保护对策。如采取天然林人工促进母树生长发育、结实,通过生境因素调控幼苗定居更新等,减少外界干扰,促进种群的自我更新;从不同的种群分布地收集繁殖材料,利用扦插、组培、细胞培养等现代生物技术手段培育具有遗传差别的个体,增加祁连圆柏的遗传变异水平,促进其进化潜能;开发建立规模化人工苗快速繁育体系,在祁连圆柏分布区和适宜区营造人工林,促进种群更新。
[1] 刘贤德,王清忠,孟好军.祁连山圆柏[M].北京:中国科学技术出版社,2006:25.
[2] 《中国森林》编辑委员会.中国森林[M].北京:中国林业出版社,1999:1107-1140.
[3] 马永俊,王金叶,刘兴明.祁连山保护区森林生态系统现状与保护对策[J].西北林学院学报,2005,20(4):5-8.MA Y J,WANG J Y,LIU X M.Status of forestry ecosystem and protection countermeasure in the protection areas in Qilian Mountains[J].Journal of Northwest Forestry University,2005,20(4):5-8.(in Chinese)
[4] 赵维俊,敬文茂,刘贤德,等.祁连圆柏生长特性研究[J].安徽农业科学,2011,39(10):5847-5849.
[5] 文陇英,陈拓,张满效,等.不同生境下祁连圆柏叶片色素和稳定碳同位素组成的变化[J].冰川冻土,2010,32(4):823-828.WEN L Y,CHEN T,ZHANG M X,et al.Variations of pigments and stable-carbon isotope ratios in Sabina przewalskii under different environments[J].Journal of Glaciology and Geocryology,2010,32(4):823-828.(in Chinese)
[6] 周宝石.祁连圆柏化学成分研究[J].陕西师范大学学报:自然科学版,2005,33(3):68-70.ZHOU B S.Chemical constituents of Sabina przewalskii[J].Journal of Shanxi Normai Universtiy:Nat.Sci.,2005,33(3):68-70.(in Chinese)
[7] 简启亮,文陇英,陈拓,等.祁连圆柏和圆柏色素含量及其花青苷合成酶活性的季节性变化[J].植物学报,2010,45(6):698-704.JIAN Q L,WEN L Y,CHEN T,et al.Seasonal changes in the contents of pigments and anthocyanins synthetase activity of Sabina przewalskii and Sabina chinensis[J].Chinese Bulletin of Botany,2010,45(6):698-704.(in Chinese)
[8] 常学向,车克钧.祁连圆柏群落生物量及营养元素积累量[J].西北林学院学报,1997,12(1):23-28.
[9] 李得庆,高国雄.生根粉处理祁连圆柏扦插育苗技术研究[J].林业机械与木工设备,2008,36(4):62-63.
[10] 薛长福.祁连圆柏在玛可河林区引种育苗造林技术[J].防护林科技,2010,96(3):120-122.
[11] 沈海岗,张宏斌,翟玉新.祁连山区祁连圆柏种子育苗技术研究[J].防护林科技,2011,101(2):5-8.
[12] 孟好军,潘爱华,姜梅.干旱山地祁连圆柏营造技术[J].甘肃科技,2004,20(11):162-163.
[13] 徐生旺.祁连圆柏育苗造林技术[M].杨陵:西北农林科技大学出版社,2007.
[14] 邵雪梅,方修琦,刘洪滨.柴达木东缘山地千年祁连圆柏年轮定年分析[J].地理学报,2003,58(1):90-100.SHAO X M,FANG Q,LIU H B.Dating the 1 000-year-old Qilian Juniper in mountains along the eastern margin of the Qaidam basin[J].Acta Geographical Sinica,2003,58(1):90-100.(in Chinese)
[15] 黄磊,邵雪梅,梁尔源.青海沙利克山祁连圆柏千年树轮宽度序列的变化特征[J].地理研究,2004,23(3):365-373.HUANG L,SHAO X M,LIANG E Y.Characteristics of millennial tree-ring width variations of Qilian juniper in Shalike Mountain,Qinghai[J].Geographical Research,2004,23(3):365-373.(in Chinese)
[16] 郑永宏,梁尔源,朱海峰,等.不同生境祁连圆柏径向生长对气候变化的响应[J].北京林业大学学报,2008,30(3):7-12.ZHENG Y H,LIANG E Y,ZHU H F,et al.Response of radial growth of Qilian juniper to climatic change under diferent habitats[J].Journal of Beijing Forestry Universtiy,2008,30(3):7-12.(in Chinese)
[17] 陈文年,吴宁,罗鹏,等.岷江上游林草交错带祁连山圆柏群落的物种多样性及乔木种群的分布格局[J].应用与环境生物学报,2003,9(3):221-225.CHEN W N,WU N,LUO P,et al.Species diversity and arbor population distribution pattern of Sabina przewalskii community in the forest-grassland ecotone in the watershed of upper Minjiang River[J].Chinese Journal of Applied Environmental Biology,2003,9(3):221-225.(in Chinese)
[18] 陈文年,吴宁,罗鹏,等.岷江上游祁连山圆柏群落优势种群间的联结性[J].应用与环境生物学报,2004,10(6):712-717.CHEN W N,WU N,LUO P,et al.Interspecific association among dominant populations in Sabina przewalskii community in the upper reaches of the Min Rever[J].Chinese Journal of Applied Environmental Biology,2004,10(6):721-217.(in Chinese)
[19] 陈文年,吴宁,罗鹏.岷江上游祁连山圆柏群落结构研究[J].应用生态学报,2005,16(2):197-202.CHEN W N,WU N,LUO P.Sabina przewalskii community structure in the upper reaches of Miniiang River[J].Chinese Journal of Applied Ecology,2005,16(2):197-202.(in Chinese)
[20] 陈文年.岷江源区祁连圆柏林线交错带的物种多样性[J].四川教育学院学报,2012,28(10):122-124.CHEN W N.Species diversity of Sabina przewalskii timberline ecotone in the source area of the Min River[J].Journal of Sichuan college of Education,2012,28(10):122-124.(in Chinese)
[21] 吴征镒,周浙昆,李德铢,等.世界种子植物科的分布区类型系统[J].云南植物研究,2003,25(3):245-257.WU Z Y,ZHOU Z K,LI D Z,et al.The areal-types of the world families of seed plants[J].Aeta Botaniea Yunnaniea,2003,25(3):245-257.(in Chinese)
[22] 吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991(Supp.Ⅵ):1-139.WU Z Y.The areal-types of the chinese genera of seed plants[J].Aeta Botaniea Yunnaniea,1991(Supp.Ⅵ):1-139.(in Chinese)
[23] 毛志宏,朱教君,谭辉.辽东山区次生林植物物种组成及多样性分析[J].林业科学,2007,43(10):1-7.MAO Z H,ZHU J J,TAN H.Plant species composition and diversity of secondary forests in montane regions of eastern Liaoning Province,China[J].Scientia Silvae Sinicae,2007,43(10):1-7.(in Chinese)
[24] 方凯,王得祥,保积存.秦岭中段典型群落多样性分析[J].西北林学院学报,2012,27(4):50-53.FANG K,AWNG D X,BAO J C.Analysis typical community diversity in the middle of Qinling Mountains[J].Journal of Northwest Forestry University,2012,27(4):50-53.(in Chinese)
[25] 郑维列,薛会英,罗大庆,等.巨柏种群的生态地理分布与群落学特征[J].林业科学,2007,43(12):8-15.ZHENG W L,XUE H Y,LUO D Q,et al.Eco-geographic distribution and coenology characteristics of Cupressus gigantea[J].Scientia Silvae Sinicae,2007,43(12):8-15.(in Chinese)
[26] KNOWLES P,GRANT M C.Age and size structure snslysis of engelmann spruce,pondrosapine,loadgepole pine,and limber pine in cororado[J].Ecology,1983,64:1-9.
[27] 杨军,傅德平,杨晓艾.艾比湖湿地自然保护区典型群落物种多样性分析[J].干旱区资源与环境,2007,24(2):145-149.YANG J,FU D P,YANG X A.Study on the species diversity of typical plant communities in Ebinur Lake Wetland Nature Reserve[J].Journal of Arid Land Resources and Environment,2007,24(2):145-149.(in Chinese)
