百合属野生种和园艺品种的染色体核型
2014-12-24张艺萍王继华吴丽芳王祥宁郭树华张世梅瞿素萍
张艺萍,王继华,吴丽芳,王祥宁,郭树华,张世梅,周 瑜,瞿素萍
(国家观赏园艺工程技术研究中心/云南省花卉育种重点实验室/云南省花卉工程技术研究中心/云南省农业科学院花卉研究所,云南昆明650200)
百合是全球较重要的花卉之一.目前市场上的百合品种来自几大系列,分别是东方百合、亚洲百合、铁炮百合、新铁炮百合、东方—铁炮百合和铁炮—亚州百合.东方百合(Lilium“Oriental Hybrids”)包括天香百合、鹿子百合、日本百合和红花百合等品种及其与湖北百合的杂种中选育出来的栽培杂种系[1-2].亚州百合(Lilium“Asiatic Hybrids”)是从亚州原产的百合,如卷丹、垂花百合、川百合、大花卷丹、朝鲜百合、山丹和鳞茎百合等品种或杂种群中选育出来的[3-4].铁炮百合(L.longiflorum Thumb)为百合科球根植物,又名麝香百合.铁炮百合花筒平展成喇叭状,香浓,花粉黄色,株高50-120 cm,最流行的顏色为白色,荷兰已育出不同颜色.铁炮—亚州百合是铁炮百合与亚州百合的杂交种,具有花色丰富的特点[5-6].
中国是世界百合遗传资源最重要的产地之一,百合野生资源非常丰富,但在百合育种工作中,还没有培育出比较知名的品系或品种.近年来,前人在百合属植物的核型研究方面已作了大量的工作[7-12],但这些工作多集中于百合野生种,对百合栽培品种的核型报道较少.为此,本试验挑选东方百合、亚州百合和LA系列百合等品(种)系的代表品种及野生种,对其染色体进行观察和核型分析,找出几者间在核型上的差异,了解其遗传特点,旨在为百合的遗传育种、分类、杂交后代真实性鉴定等研究提供一定的细胞学依据.
1 材料与方法
1.1 材料
材料有东方百合品种“Berlin”和“Parasol”、亚州百合品种“Saniciro”、LA系列百合品种“Royal sunset”、百合野生种“大花卷丹”[L.leichtlinii Hook.f.var.maximowiczii(Regel)Baker]、湖北百合(L.henryi Baker).
1.2 方法
采用常规压片法,参照王红霞等[13]和荣立苹等[14]的染色体制备方法,并稍作改动.选取生长适宜的根尖用0.1%秋水仙素预处理;用卡诺固定液在0℃的冰水中固定30 min;用95%乙醇清洗3 s后用蒸馏水洗30 min;用解离液于60℃的水浴锅中酸解6 min;用卡宝品红染色30 min后压片镜检,对分散较好的根尖进行拍照.按李懋学等[15]的方法对获得的根尖染色体进行核型分析.核型类型参照Stebbins[16]的标准,按核型中最长与最短染色体的比值及臂比值大于2的染色体所占比例划分.核型不对称系数/%=长臂总长/全组染色体总长×100,采用Arano[17]的方法计算.
2 结果与分析
Berlin、Parasol、Saniciro、Royal sunset、大花卷丹和湖北百合的染色体数均为二倍体,2n=2x=24,染色体参数和核型特征分别见表1和2,中期染色体形态和核型分别见图1.
2.1 Berlin和Parasol的染色体核型
Berlin的核型公式为2n=2x=24=4m+18st+2sm,第1和2对为中部着丝粒染色体,第12对为近中部着丝粒染色体,第3-11对为近端部着丝粒染色体.最长与最短染色体比值为2.22,臂比值大于2∶1的染色体占全部染色体的83.33%,属于3B型.核型不对称系数为75.19%.
Parasol的核型公式为2n=2x=24=4m+10sm+8st+2t,第1和6对为中部着丝粒染色体,第2、7、9-11对为近中部着丝粒染色体,第3、4、8和12对为近端部着丝粒染色体,第5对为端部着丝粒染色体.最长与最短染色体比值为2.11,臂比值大于2∶1的染色体占全部染色体的66.67%,属于3B型.核型不对称系数为72.40%.
2.2 Saniciro的染色体核型
Saniciro的核型公式为2n=2x=24=2m+2sm+10st+10t,第1对为中部着丝粒染色体,第8对为近中部着丝粒染色体,第2、6、9-11对为近端部着丝粒染色体,第3-5、7和12对为端部着丝粒染色体.最长与最短染色体比值为1.94,臂比值大于2∶1的染色体占全部染色体的91.67%,属于3A型.核型不对称系数为83.15%.
2.3 Royal sunset的染色体核型
Royal sunset的核型公式为2n=2x=24=8sm+10st+6t,第1、3、7和10对为近中部着丝粒染色体,第4、6、8、9和11对为近端部着丝粒染色体,第2、5和12对为端部着丝粒染色体.最长与最短染色体比值为1.58,全部染色体均为臂比值大于2∶1的染色体,属于4A型.核型不对称系数为80.98%.
2.4 大花卷丹和湖北百合的染色体核型
大花卷丹的核型公式为2n=2x=24=2sm(SAT)+10st(SAT)+12t(2SAT),第1对为近中部着丝粒染色体,第6、9-12对为近端部着丝粒染色体,第2-5、7和8对为端部着丝粒染色体.其中,第1对染色体上有1个居间随体,第3、4和6对染色体上各有1个近端随体(图1E).最长与最短染色体比值为1.88,全部染色体均为臂比值大于2∶1的染色体,属于4A型.核型不对称系数为85.78%.湖北百合的核型公式为2n=2x=24=10sm+2st+12t,第5对为近中部着丝粒染色体,第7、9-12对为近端部着丝粒染色体,第1-4、6和8对为端部着丝粒染色体.最长与最短染色体比值为1.71,全部染色体均为臂比值大于2∶1的染色体,属于4A型.核型不对称系数为86.75%.
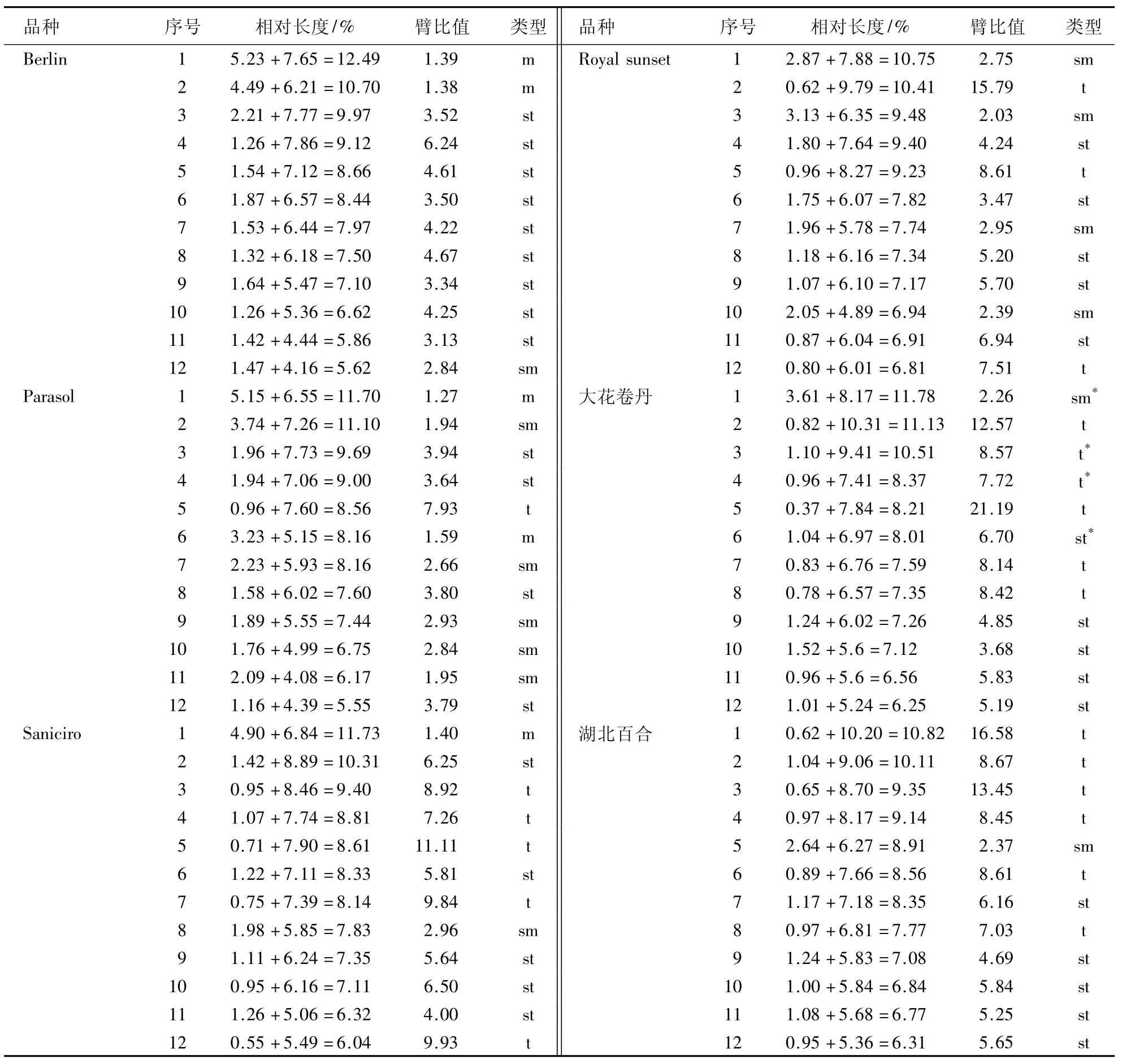
表1 百合的染色体参数Table 1 The parameters of the chromosomes of lily

表2 百合的核型特征Table 2 The parameters of the karyotypes of lily

图1 百合品种的染色体形态和核型图Fig.1 Chromosome and karyotype of lily
3 讨论
本试验中,Berlin、Parasol、Saniciro、Royal sunset、大花卷丹和湖北百合的染色体均为24条,且都有1对中部着丝点染色体或近中部着丝点染色体,这与百合属植物的核型共有特征[18]一致.根据有花植物核型进化中的对称—原始、不对称—进化的观点[19]以及核型不对称的分类标准,百合属植物所有核型均为极不对称型,且最长与最短染色体比值较大,是比较进化的类型.本试验中,Berlin的最长与最短染色体比值为2.22,核型不对称系数为75.19%;Parasol的最长与最短染色体比值为2.11,核型不对称系数为72.40%;Saniciro的最长与最短染色体比值为1.94,核型不对称系数为83.15%;Royal sunset的最长与最短染色体比值为1.58,核型不对称系数为80.98%;大花卷丹的最长与最短染色体比值为1.88,核型不对称系数为85.78%;湖北百合的最长与最短染色体比值为1.71,核型不对称系数为86.75%.这些数据都证实了上述结论.
本试验中,2个东方百合品种为3B型,1个亚州百合品种为3A型,与戴晓红等[4]的研究结果基本相似.戴晓红等[4]采用的东方百合品种“Siberia”为3B型,亚洲百合品种“Pollyanna”为3A型,表明本试验的结果具有较好的参考价值.岳玲等[2]得出大花卷丹的核型公式为2n=4m(2SAT)+10st+10t,核型为3A型,而本试验大花卷丹的核型为4A型,并在第1、3、4和6对染色体上发现随体,可能与大花卷丹在云南的栽培环境有关.戴小红等[10]发现湖北百合的核型为3B型,而本试验湖北百合的核型为4A,可能也与云南的栽培环境有关.
[1]JONG S L.Influence of frozen storage duration and forcing temperature on flowering of oriental hybrid lilies[J].Hortscience,2001,36(6):1053-1056.
[2]岳玲,雷家军,胡新颖.大花卷丹和卷丹的核型研究[J].黑龙江农业科学,2006(6):41-43.
[3]MASUMI Y.A genetic model for a pollenless trait in Asiatic hybrid lily and its utilization for breeding[J].Scientia Horticulturae,2003,98:293-297.
[4]戴小红,牛立新,张延龙.百合三品系代表品种的核型分析[J].西北林学院学报,2006,21(4):58-61.
[5]ARZATE FERNÁNDEZ A M,NAKAZAKI T,TANISAKA T.Production of diploid and triploid interspecific hybrids between Lilium concolor and L.longiflorum by in vitro ovary slice culture[J].Plant Breeding,1998,117(5):479-484.
[6]孙晓梅,贾莲,杨宏光,等.新铁炮百合3个品种的核型分析[J].园艺学报,2010,37(6):1003-1008.
[7]杨雪珍,张克中,贾月慧,等.21个百合栽培品种的核型分析[J].中国农业大学学报,2011,16(3):100-106.
[8]荣立苹,雷家军,毕晓颖,等.8种野生百合染色体数目的变异[J].东北林业大学学报,2009,37(9):48-50.
[9]赵西宁,王文和,郝培.百合6个品种的染色体核型分析[C]∥张启翔.中国观赏园艺研究进展.北京:中国林业出版社,2010:183-187.
[10]戴小红,张延龙,牛立新.百合属4种植物的核型研究[J].西北植物学报,2006,26(1):50-56.
[11]刘冬云,张晓曼,杨楠,等.兰州百合和有斑百合的核型研究[J].西北农林科技大学学报:自然科学版,2009,37(11):102-106.
[12]王树红,李巧峡,赵庆芳,等.四种观赏百合的核型分析[J].西北师范大学学报:自然科学版,2007,43(4):93-97.
[13]王红霞,王文锋,丰慧根.两种百合染色体变异比较研究[J].生物学杂志,2006,23(2):42-44.
[14]荣立苹,雷家军,王志刚.百合根尖染色体制片技术研究[J].安徽农业科学,2007,35(7):2006-2009.
[15]李懋学,张赞平.作物染色体及其研究技术[M].北京:中国农业出版社,1996:1-37.
[16]STEBBINS G L.Chromosomal Evolution in Higher Plants[M].London:Edward Arnoold Ltd,1971:72-123.
[17]ARANO H.The karyotypes and the speciations in subfamily Carduoideae of Japan[J].Jap Journ Bot,1965,19(3):31-67.
[18]STEWART R N.The morphology of somatic chromosomes in Lilium[J].Amer J Bot,1947,34:9-26.
[19]洪德元.植物细胞分类学[M].北京:科学出版社,1990:91-96.
