利用分子标记辅助选择培育转Bt 基因抗虫水稻
2014-12-14魏志刚裴婷婷胡颂平罗利军叶水峰刘国兰
张 琳,魏志刚,裴婷婷,胡颂平,余 霞,张 静,罗利军,叶水峰,刘国兰
(1.江西农业大学 理学院,江西 南昌 330045;2.江西农业大学 生物科学与工程学院,江西 南昌 330045;3.上海市农业生物基因中心,上海 201106)
水稻是最重要的粮食作物之一,是世界1/3 人口的主要粮食来源。而虫害是造成水稻减产的主要原因之一,其中水稻鳞翅目害虫如二化螟、三化螟、稻纵卷叶螟等是我国水稻生产中的主要害虫。据统计,全世界每年因虫害造成的经济损失就达近百亿美元。因此,培育抗螟虫的水稻新品种具有十分重要的现实意义和应用前景。苏云金芽胞杆菌(Bacillus thuringiensis,简称Bt)是一种革兰氏阳性菌,属于原核生物细菌纲芽胞杆菌科芽胞杆菌属,广泛存在于自然界[1]。1901 年日本学者首次分离出Bt 菌,并证明它对部分鳞翅目昆虫有毒性作用。1953 年发现Bt 菌的杀虫活性与伴胞晶体有关[2],并证实这种伴胞晶体由蛋白质组成[3]。这种蛋白通常被称作δ-内毒素(δ-endotoxins)或杀虫晶体蛋白(insecticidal crystal protein,ICP)。随着人们对Bt 菌及其所产生的杀虫晶体蛋白研究的逐步深入,人们已从不同的Bt 菌的亚种中分离出对不同昆虫(如鳞翅目、鞘翅目、双翅目等)和无脊椎动物(如螨类、寄生线虫、原生动物等)有特异毒杀作用的杀虫晶体蛋白。
Schnepf 和Whiteley[4]将Bt 库斯塔克亚种菌株HD-1 中的杀虫晶体蛋白基因克隆到大肠杆菌的质粒中,克隆了第1 个Bt 杀虫晶体蛋白基因[4]。Zambryskpi[5]则在1983 年成功的获得了第1 株转基因烟草,这两个试验的成功促使了转Bt 抗虫植物的出现。随后,有学者将Bt 基因转入烟草[6-8]、西红柿[9]、棉花[10]等获得了相应的转Bt 基因植物。但获得的这些早期的转Bt 基因植株的抗虫性都很弱,难以检测出mRNA 的转录,蛋白质表达量很低。可能和野生Bt 基因的AT 含量高和(或)密码子使用偏爱有关。因此,要使得Bt 基因在转基因植物中高效表达,必须对野生Bt 基因进行有效的改造。1990 年,Perlak 等[10]通过使用人工修饰过的cryIA(b)和cryIA(c)基因获得了转Bt 基因的棉花。cryIA(b)和cry-IA(c)基因在植物中的表达水平大大提高,其蛋白含量占到了植株总可溶蛋白的0.05%~0.1%,其抗虫效果可以满足农业上的应用[10]。郭三堆等[11]人工合成了全长1 824 bp 的cry1Ab 和cry1Ac 融合的GFM 杀虫基因,结果Bt 毒蛋白在植物中的表达量提高了约100 倍[11]。第1 例Bt 转基因水稻是由Wünn 等[12]获得。而Maqbool[13]则报道了在1999 年首次获得了双价的Bt(cry1Ac+cry2A)水稻。Tu[14]则在2000 年首次报道了转Bt 水稻的田间试验。到目前为止,已经有大量的转Bt 基因水稻的报道。林拥军[15-16]经人工改造获得具有自主知识产权的两个抗虫基因:cry1C*和cry2A*,并培育出高抗的转基因品系T1C-19 和T2A-1。这两个转基因品系及其配制的杂种对二化螟、三化螟和稻纵卷叶螟等鳞翅目害虫的抗虫效率大于95%。为了更有利于推动抗虫转基因水稻的商品化,Ye 等[17]采用胚乳特异不表达的rbcS 启动子来驱动cry1C*基因,得到的转基因株系具有高抗虫性,Cry1C 蛋白在叶片和茎秆中高表达,而在胚乳的表达量极低[17]。
目前,水稻生产主要采取喷洒杀虫剂来控制虫害。虽然化学杀虫剂在减少虫害损失起到了巨大作用,但是它的使用不可避免的造成环境污染以及威胁人类健康,而且提高了农业生产的成本。通过大规模的筛选,目前在水稻及其近缘种中还没有发现二化螟、三化螟、稻纵卷叶螟等鳞翅目害虫的抗性种质资源,因此无法通过常规育种及分子标记辅助选择技术方式来培育抗虫水稻新品种。通过转基因育种的手段增强水稻自身的抗虫性是解决上述问题的有效途径。Bt 基因是来自于苏云金芽胞杆菌的一种杀虫基因,其表达的蛋白产物可特异性毒杀不同的害虫,而且对人畜和环境无害,是目前转基因育种中主要使用的抗虫基因。应用转基因培育抗虫水稻是现阶段及未来解决水稻虫害问题的主要策略。
本研究的目的是通过传统杂交的方法将3 个Bt 明恢63 中的Bt 基因,转育到水稻品种秀水123、湘晴及节水抗旱稻恢复系旱恢3 号和保持系沪旱1B 中。利用分子标记辅助选择、田间除草剂筛选或者试纸条检测方法,以获得新型转Bt 基因株系,并筛选出抗虫性良好、农艺性状优良的株系,为后续抗虫育种提供材料。
1 材料与方法
1.1 材料
1.1.1 供体植物材料 本试验研究的水稻材料为Bt 明恢63 株系:TT51(cry1Ac/Ab)、T1C-19(cry1C*)、T2A-1(cry2A*),以上材料来自华中农业大学林拥军教授。
1.1.2 受体材料 受体亲本为常规粳稻品种秀水123,恢复系湘晴以及节水抗旱稻恢复系旱恢3 号和保持系沪旱1B。
1.1.3 供试昆虫 中进行大田抗虫鉴定试虫为水稻主要鳞翅目害虫之一:稻纵卷叶螟。
1.1.4 试剂 1.5×CTAB;氯仿/异戊醇(24∶1);引物(序列见表1,由Invitrogen 公司合成)Taq 酶;Buffer;dNTP;琼脂糖;溴酚蓝;EB;Basta 试剂(除草剂)等。

表1 Bt 基因扩增引物Tab.1 Amplification primer of Bt gene
1.2 实验方法
1.2.1 研究技术路线 利用抗虫转基因新材料T1C-19(携带cry1C*基因)、T2A-1(携带cry2A*基因)和TT5-1(携带cry1Ab/Ac 融合基因)为供体材料,以秀水123、湘晴、旱恢3 号和沪旱1B 为受体材料,进行有性杂交和回交,并通过自交来获得纯合转Bt 基因株系(图1)。本研究主要是对回交二代自交三代(BC2F3)进行分子标记阳性检测,得到纯合转Bt 基因株系。同时,采用田间除草剂筛选、试纸条检测方法及田间抗虫鉴定等方法来鉴定是否纯合。

图1 研究技术路线Fig.1 The route of study technique
1.2.2 分子标记阳性检测 (1)水稻叶片总DNA 提取:采用小量DNA 抽提法(CTAB 法)。
(2)PCR 扩增:采用常规PCR 扩增。
(3)电泳检测。1)琼脂糖凝胶电泳:①制胶。a.6 g 琼脂糖加到盛有275 mL 0.5×TBE 的锥形瓶中煮沸10 min 至琼脂糖充分溶解;b.当锥形瓶温度将为60 ℃左右加入12 μL 的EB 混合均匀;c.将琼脂糖溶液倒入制胶模中,在适当的位置插上梳子;d.在室温下使胶凝固,之后拔出梳子将胶放入电泳槽中。②点样。模板DNA 8 μL,Maker 4 μL。③0.5×TBE 缓冲液中120V 电压跑胶至溴酚蓝跑到底。④将胶置于紫外光下观察,拍照记录。2)聚丙烯酰胺凝胶电泳:采用常规聚丙烯酰胺凝胶电泳。
1.2.3 转基因植株的除草剂抗性检测 由于转基因植物具有抗除草剂的特性,可以利用喷洒除草剂的方法来检测转基因植株。转基因植株的除草剂抗性检测方法主要有两种:Basta 溶液涂抹叶片和Basta 溶液直接喷洒。本研究采用后一种,配制终浓度为0.3%的Basta 直接在田间喷施水稻植株,7 d 后进行观察。
1.2.4 转基因植株的试纸条检测 取标记检测阳性植株叶片,加自来水于碾钵中磨碎,取汁液于离心管中,将胶体金试纸置于汁液中,观察结果。
1.2.5 转基因植株的田间抗虫鉴定 全生育期实行除不喷施农药外的正常田间管理,观察转基因植株对卷叶螟的抗性。田间种植材料整个生育期不打药,通过自然发虫进行材料选择,由于近年螟虫危害较重,田间筛选结果较好。在田间共种植有BC2F3材料,根据检测的阳性植株以及田间发虫情况来进行选种,选择了植株表现上与轮回亲本接近的,且抗虫的材料共270 份材料。
2 结果
2.1 分子标记阳性检测结果
对所获得2A 材料93 份,1C 材料112 份,1Ab/Ac 材料65 份进行水稻植株提取DNA,通过PCR 及琼脂糖电泳检测,因为Bt 基因是显性的,因此在BCnF1 代有扩出对应条带的为阳性植株(图2、3、4)。而自交后的植株中如果全部的单株都能扩增出Bt 基因,则为纯合株系。BC2F3代PCR 检测的结果显示分别获得cry1C*和cry2A*基因的一些纯合株系(表2),而cry1Ac/Ab 暂还未获得纯合株系,需下一代自交挑选。
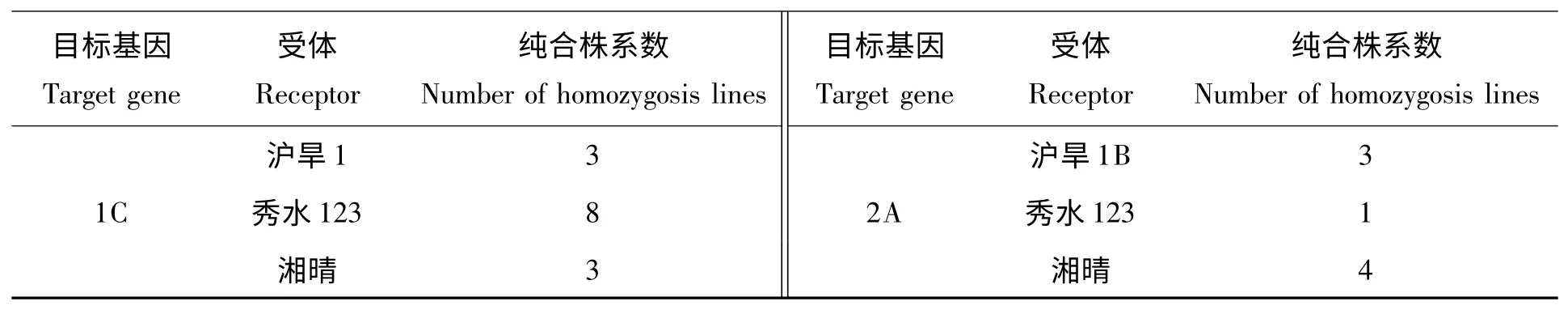
表2 BC2F3代纯合株系Tab.2 Homozygous lines of BC2F3

图2 Cry2A* 琼脂糖凝胶电泳结果Fig.2 Agarose gel electrophoretogram of Cry2A*

图3 Cry1C* 琼脂糖凝胶电泳结果Fig.3 Agarose gel electrophoretogram of Cry1C*
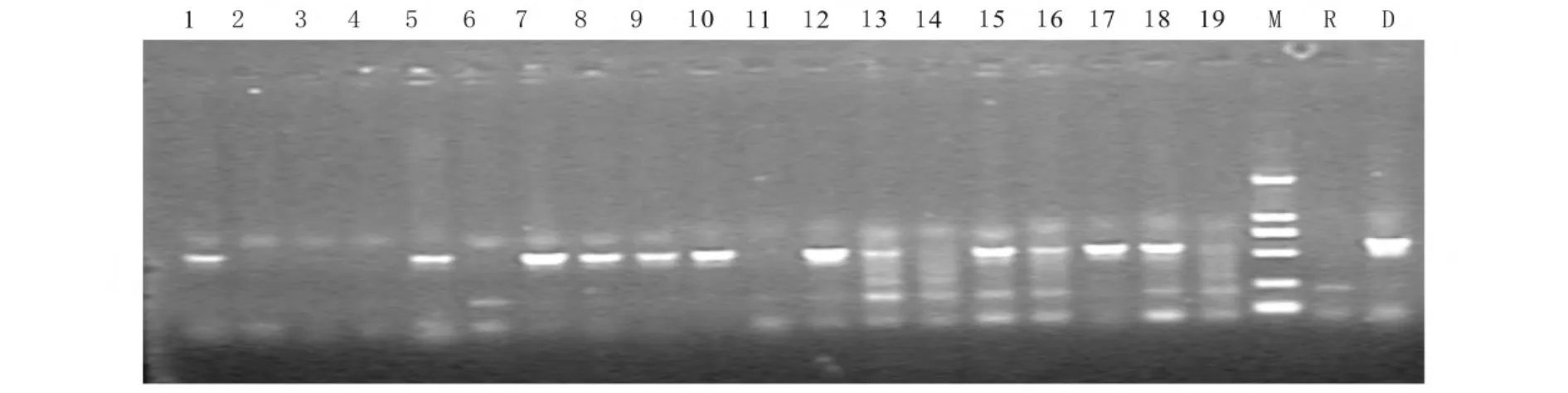
图4 Cry1Ab/Ac 琼脂糖凝胶电泳结果Fig.4 Agarose gel electrophoretogram of Cry1Ab/Ac

图5 田间喷施Basta 试验.Fig.5 Experiment of spraying Basta in field
2.2 转基因植株的Basta 喷施试验结果
除常规的PCR 检测外,对cry1C*和cry2A*植株进行0.3%的Basta 喷施检测,进一步确认阳性植株。在BC2F3代进行的纯合株系挑选,若为纯合株系,则所有单株均不受Basta 损害;而杂合的则出现3∶1 的分离比(阳性:阴性),本次实验杂合植株总株数为192 株,其中表现阳性的为144 株,表现阴性的为48 株;全部死亡的则为阴性株系(图5)。
2.3 试纸条检测结果
除常规的PCR 检测外,对转基因植株进行胶体金试纸进行蛋白的阳性鉴定。进一步确保筛选结果的可靠性(图6)。其中cry1C*和cry2A*标记结果和试纸条检测结果相吻合。而对于Cry1Ab/Ac,PCR 结果阳性,但试纸检测却为假。

图6 试纸条检测结果Fig.6 Results of test paper detection
2.4 田间稻纵卷叶螟抗性鉴定结果
2011 年夏在上海白鹤转基因试验基地进行材料的稻纵卷叶螟抗性鉴定。已转入Bt 基因的植株对稻纵卷叶螟表现出高抗性,卷叶率为0;而阴性植株则被侵食,卷叶率平均达37.8%(图7)。此外,含cry2A*植株的抗性相对较差,有个别植株受轻微影响,有1~2 片叶片被侵害。

图7 田间自然感虫试验结果Fig.7 Results of natural affection of rice leaf roller in field
3 讨论与结论
在转基因植株检测中普遍PCR 检测的假阳性结果较多,给进一步的检测造成麻烦。Cry1Ab/Ac 的问题,PCR 结果阳性,但试纸检测却为假。所以Cry1Ab/Ac 暂未获得纯合株系,需进一步自交。
在选择琼脂糖凝胶电泳检测供体-受体间差异标记时,产物不清晰,即在3%琼脂糖凝胶电泳中,供体与受体条带一致,需用5%聚丙烯凝胶电泳才能检测出差异。而聚丙烯酰胺凝胶在制备过程中会受到很多因素的影响:首先是催化剂AP(过硫酸铵)与加速剂TEMED(二甲基乙二胺)浓度。适量的AP与TEMED 促进聚合,但当其过量时会引起电泳时烘胶和谱带变形。且温度高时聚合快,所以应选择合适配方使聚合在40~60 min 中内完成。Acr(单体丙烯酰胺)与交联剂Bis(N,N-甲叉丙烯酰胺)的浓度也是影响凝胶形成的重要因素。此外温度,pH,氧分子都会对凝胶的形成及凝胶的质量造成影响。
本实验在使用最普遍的分子标记辅助选择转Bt 基因水稻同时[18],采用田间除草剂筛选;试纸条检测方法;田间抗虫鉴定等方法相结合筛选出抗虫性良好、农艺性状优良的株系,为后续抗虫育种提供材料。多种方法并用的优势是节省工作量以及增强结果准确性等。通过直接对生长期水稻叶片喷洒Basta 实验,若为阴性植株则单株死亡,可以通过此方法来减少需要进行PCR 阳性检测的植株数量;此外也可以利用此方法来检测通过分子标记辅助选择的植株是否为假阳性,因为假阳性植株在喷洒Basta 液体后一周内会死亡。试纸条检测方法是一种高效、简便、易操作、准确率高的一种方法。将待测水稻叶片加水磨成液体后利用试纸条可在一、两分钟内判断出所测材料为阴性还是阳性。且Cry1Ab/Ac 的为假阳性正是通过试纸条检测方法检测出来的。而田间抗虫鉴定则是检测材料植株是否抗虫及抗性强弱的田间实际标准。
通过转基因育种的手段增强水稻自身的抗虫性,在实践中筛选出抗虫性良好、农艺性状优良的株系,这依然需要更多的时间与实践。
本试验经BC2F3代PCR 检测及试纸条检测的结果显示分别获得cry1C*和cry2A*基因的沪旱1B、秀水123 和湘晴的纯合株系。同时,在田间自然诱发虫害条件下,转基因株系对稻纵卷叶螟虫表现出强抗性。利用分子标记辅助选择(MAS)等技术可以培育出抗虫性、农艺性状优良的转Bt 基因水稻株系或品种。
[1]Phyllis A W M,Russell S T.Worldwide abundance and distribution of Bacillus thuringiensis isolates[J].Appl Environ Microbiol,1989,55(10):2437-2442.
[2]Hannay C L.Crystalline inclusions in aerobic sporeforming bacteria[J].Nature,1953,172(11):1004.
[3]Hannay C L,Fitz J P.The protein crystals of bacillus thuringiensis Berliner[J].Can J Microbiol,1955,1(8):694-710.
[4]Schnepf H E,Whiteley H R.Cloning and expression of the bacillus thuringiensis crystal protein gene in Escherichia coli[J].Proc Natl Acad Sci,1981,78(5):2893-2897.
[5]Zambryskpi P,Joos H,Genetello C,et al.Ti plasmid vector for the introduction of DNA into plant cells without alteration of their normal regeneration capacity[J].EMBO J,1983,2(12):2143-2150.
[6]Adang.In biotechnology in invertebrate pathology and cell culture[M].New York:Academic Press,Inc,1987:85-99.
[7]Barton K A,Whitely H R,Yang N S.Bacillus thuringiensis delta-endotoxin expressed in transgenic Nicotiana tabacum provides resistance to lepidopteran insects[J].Plant Physiol,1987,85:1103-1109.
[8]Vaeck M,Reynaerts A,Hofte H,et al.Transgenic plants protected from insect attack[J].Nature,1987,238:33-37.
[9]Fischhoff D A,Bowdish K S,Perlak F J,et al.Insect tolerant transgenic tomato plants[J].Bio/Technology,1987,5:807-813.
[10]Perlak F J,Deaton R W,Armstrong T A,et al.Insect resistant cotton plants[J].Bio/Technology,1990,8:939-943.
[11]郭三堆,洪朝阳,王京红,等.苏云金芽孢杆菌鲇泽变种7-29 杀虫蛋白质结构基因的改造和表达[J].微生物学报,1992,32(5):167-l75.
[12]Wünn J,Kloti A,Burkhardt P K,et al.Transgenic indica rice breeding line IR58 expressing a synthetic cry1A(b)gene from Bacillus thuringiensis provides effective insect pest control[J].Bio/Technology,1996,14:171-176.
[13]Maqbool S B,Riazuddin S,Loc N T,et al.Expression of multiple insecticidal genes confers broad resistance against a range of different rice pests[J].Mol Breed,2001,7:85-93.
[14]Tu J,Zhang G,Datta K,et al.Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringensis endoprotein[J].Nature BioTechnology,2000,18:1101-1104.
[15]Chen H,Tang W,Xu C G,et al.Transgenic indica rice plants harboring a synthetic cry2A*gene of Bacillus thuringiensis exhibit enhanced resistance against lepidopteran rice pests[J].Theor Appl Genet,2005,111(7):1330-1337.
[16]Tang W,Chen H,Xu C G,et al.Development of insect—resistant transgenie indica rice with a synthetic cry1C gene[J].Mol Breed,2006,18(1):1-10.
[17]Ye R J,Huang H Q,Yang Z,et al.Development of insect-resistant transgenic rice with Cry1C-free endosperm[J].Pest Manag Sci,2009,65:1015-1020.
[18]陈凯,张强,潘晓飚,等.三个中籼稻骨干恢复系对稻瘟病和褐飞虱抗性改良效果的评价[J].核农学报,2013,27(8):1069-1080.
