基于格兰杰因果分析的针刺不同效应阶段中枢神经传导通路研究
2014-12-13孙传铸白丽君牛璇陈红艳陈鹏张明劳力行
孙传铸,白丽君*,牛璇,陈红艳,陈鹏,张明,劳力行
1.西安交通大学生命科学与技术学院,生物医学信息工程教育部重点实验室,西安 710049
2.西安交通大学第一附属医院影像科,西安 710061
3.首都医科大学附属北京天坛医院放射科,北京 100050
4.首都医科大学附属北京中医医院针灸中心,北京 100010
5.香港大学中医药学院,香港
6.美国马里兰大学医学院结合医学中心,巴尔的摩 21201
针刺作为一种补充与替代医疗方式,近年来在国际范围掀起了应用热潮。虽然针刺已获得了广泛的社会认可,但针刺潜在的生理生物学机制仍然是一个悬而未决的科学问题,仍需进行更为深入的研究。在众多的问题中,针刺效应所涉及的中枢神经传导通路的相关证据还很缺乏。
至今,绝大多数神经影像研究着重探讨针刺即时效应中枢神经响应的空间分布。事实上,针刺存在两个特征性的作用阶段:(1)针刺捻针操作阶段,这一过程主要涉及对深层组织的刺激(表皮刺戳)以及机体对组织伤害的生化反应;(2)针刺刺激结束后的持续性效应作用阶段。人类行为和动物研究得到的实验证据也表明:针刺镇痛存在一个显著特征,即针刺的持续性效应,即针刺作用需要经历一段时间才可到达峰值,然后逐渐回归到基线水平[1-3]。研究者还证实针刺单纯的物理刺激和针刺的持续性效应,可以激活大脑许多重叠脑区[4-7]。因此,考量针刺引发大脑的动态响应变化的过程模式,对于理解针刺的中枢神经作用机制就显得至关重要。
针刺类似于一项复杂的体感刺激过程,需要大脑特定脑区信息的流动和整合,因此脑网络连接度分析特别适宜于此项问题的解决。采用独立成分分析的研究报道表明,真针刺相较于对照组,可以引起默认脑网络和感觉运动脑网络内部核团间连接度的增加,并与副交感神经的输出信号显著相关[8]。我们最近的研究也提出针刺可以进一步增强反相关脑网络的二分性,并且主要对凸显网络产生特异性调节,特别是旁边缘系统的脑区和脑干[6]。虽然目前研究已得到了一些有价值的研究证据,但仍有一些重要问题亟待解决:(1)这些被针刺调节的广泛脑网络区域间是否存在因果作用关系,是否可以为针刺功能特异性相关的信息流动作出解释;(2)刺激后的静息状态的因果相互作用拓扑结构是否与先前的刺激状态有关。
为了回答这些问题,我们首先对比针刺足三里穴和旁开非经非穴即时效应所引发的大脑激活响应,接着应用多元格兰杰因果有效连接分析对脑区间的信息流动以及方向性进行判定,并由此推测针刺在刺激操作阶段及持续性效应阶段是否会引发其功能特异性的调节机制[9]。近些年,格兰杰因果分析在脑认知研究方面得到了广泛应用,已成为一种适用于神经生理学数据的频率和时间域指标来评价神经元之间相互作用方向性的有效研究途径[10-12]。该方法认为如果第二个时间序列与第一个时间序列之间存在因果关系的相互影响,则可以通过包含第二个时间序列的过去信息来提高对第一个时间序列的自回归预测。在这个研究中,因果作用关系的研究通过多元格兰杰因果分析的直接传递函数(direct transfer function)来实现。
1 材料与方法
1.1 研究对象
为了降低被试者间差异性的影响,16名被试者均为在校大学生,男女各8名;平均年龄(22.5±1.8)岁,身体状况良好,无针刺过敏反应,经Edinburgh利手问卷调查均为右利手;被试者及其一级亲属无精神病史,头部无损伤,近1个月内无药物服用经历;无任何fMRI扫描禁忌。在向被试者详细说明实验具体实施步骤后,被试者个人自愿签订了由西部中国医院委员会的人类计划制定的同意协议书。本实验同时也符合赫尔辛基宣言的内容。
目前针刺的fMRI研究大都采用多组块实验设计模式,fMRI技术的实验研究基于如下假设:大脑对于刺激的反应是线性时不变的,只存在相应刺激阶段和基线状态。由于针刺行为是缓慢发展并发挥作用的,针刺的血液动力学响应与多组块实验设计的基本假设是不相符的[7]。除此以外,在短时间内使用多个刺激模块,研究人员可能无法区分当前的大脑活动是由针刺引起的还是受其延迟效应的影响。在本实验中,采用了一个全新的实验模式:非重复事件相关的功能MRI实验设计来研究针刺的持续性效应。
本实验包括三个步骤。静息状态持续12.5 min。针刺真穴和针刺假穴均采用非重复事件相关的实验设计模式,持续1.5 min的针刺,两次刺激之间有1分钟的静息扫描,针刺结束后仍有12.5 min的静息扫描。实验设计对被试者盲试,要求被试者保持放松并且不要有太多的思绪。在实验进行过程中,要求被试者闭眼休息,以避免被试者观察到正在实施的刺激类型。根据实验过后参与者的报告,他们申明在整个实验过程中保持着清醒状态。针刺足三里与针刺假穴的实验顺序完全伪随机,对于所有被试者实施顺序完全平衡。为了避免针刺后效应的影响,2次针刺实验之间至少相隔72 h[7]。
针刺真穴组选取右腿足三里穴。该穴被广泛用于临床针刺镇痛以及内脏调节,是针刺机理研究中普遍采用的穴位。针刺操作过程选用无菌、一次性、不锈钢针(直径为0.2 mm,长为40 mm);入针深度为2~3 cm,针刺刺激采用平补平泻提拉捻转的手法,旋转频率为1 Hz;所有被试者的针刺过程均由1名具有专业资格的中医针灸师来完成。针刺假穴是由1名经验丰富的针灸师设计的,刺激点为非经非穴(足三里旁开2~3 cm处);并且与针刺真穴具有相同的刺激深度,刺激强度和操作方法。
在针刺实验结束后,要求每名被试者对实验过程中体验的得气感按以下的几个维度进行打分(10分VAS量表),酸、麻、胀、冷、热、尖锐痛、钝痛、重、麻刺感、痒、持续疼、挤压感。打分量表的评定标准为:1~3 轻度,4~6 轻度不适,7~8 不适,9~10 极度不适,10以上为无法忍受[13]。
1.2 数据采集和分析
实验采用GE公司的3.0 T 的MR扫描仪。一个头部固定器用来防止头动影响。32个轴位片(参数:FOV 240 mm×240 mm,matrix 64×64,thickness 5 mm),平行于AC-PC平面和覆盖使用T2加权单次获得全脑,功能图像采用BOLD对比梯度回旋脉冲序列(参数为:TR 1500 ms,TE 30 ms,flip angle 90°)。在进行功能扫描之前,结构扫描以1 mm×1 mm×1 mm的体素进行结构定位,并且使用3D MRI序列进行扫描(参数:TR 2700 ms,TE 3.39 ms,matrix 256×256,FOV 256 mm×256 mm,fl ip angle 7°,slice thickness 1 mm)。
1.3 数据预处理
采用统计参数图分析软件(SPM5,http://www.fil.ion.ucl.ac.uk/spm/)。首先,采用最小二乘法对功能数据进行头动校正(所有被试者的头动距离平均都未超过1 mm,旋转角度未超过一度);其次,对校正后的数据利用MNI标准模板进行空间标准化,并将体素冲切到2 mm×2 mm×2 mm。最后,采用6 mm FWHM的高斯核函数对数据进行平滑,以达到降低噪声,提高信噪比的目的。数据都被颜色标记并且映射到Talairach空间[14]。
1.4 从格兰杰因果分析中得到有效连接网络
为了探索在不同针刺刺激下脑网络的不同交互作用关系,将真针与假针在即时效应阶段的差异性脑区作为感兴趣区域,并进行后续的格兰杰因果分析。在广泛的边缘系统和皮层下区域,针刺真穴组和针刺假穴组相比,大脑表现了增强的激活响应(P<0.005,FDR 校正8个连续体素 的空间范围)。降低的激活响应区域并未达到统计显著值。对于双侧激活区域,只选择T值较大的侧别作为代表。最终,我们挑选了15个感兴趣区域点应用于随后的格兰杰因果分析方法中。以每个区域的峰值坐标点为中心做6 mm半径的球型区域提取时间序列,并将时间序列进行平均。格兰杰因果分析方法使用了直接传递函数(DTF)[9],在选定的种子点中计算时间序列的多元自回归模型。在这项研究中,我们还采用了加权直接传递函数用来加强直接连接并抑制间接连接的影响[8,10,15]。为了实现连接强度的显著性检验,采用替代性数据产生2500次随机采样的数据分布,同时利用这一数据分布计算DTF值[9,15-16],最后将DTF值和空的分布通过P值为0.01的单边检测进行比对(多重比较的FDR校正)。
2 结果
2.1 行为数据结果
对被试者报告的得气感12种类型的体验频率进行了统计。其中,相较于针刺假穴,麻(真针:43.4%;假针:22.1%,P<0.01),胀(真针:58.9%,假针:21.7%,P<0.005),和酸(真针:68.3%,假针:23.8%,P<0.0005)这三种感觉在针刺足三里后出现频率较高(Fish检验)。体验强度也进行了统计。针刺足三里以及针刺假穴所引发得气感的平均强度基本相等(足三里2.4±1.7;假穴2.2±1.9),强度值都比较低(轻度到中度)。
2.2 即时刺激效应下的有效连通网络
因果连接图借助箭头的粗细表示因果影响的强度(图1)。 显著的路径权重在图1中呈现,同一方向和相反方向的变化表征bold信号在2个感兴趣区域间的正相关性和反相关性。从连接图中可以看出,对于真针,背侧丘脑中部是脑网络的中枢,与背外侧前额叶以及壳核有双向连接,同时投射信号到前脑岛。背外侧前额叶和壳核存在双向作用,也同时投射到前脑岛。与丘脑内侧上行的神经通路相比,丘脑外侧的连接主要位于躯体感觉皮层。从腹后侧丘脑产生的路径通常都直接指向初级体感区和次级体感区。初级体感区和次级体感区以及腹 后侧丘脑都有双向作用,同时传到前额下部脑区。而针刺假穴,网络构成节点和针刺真穴一样。然而,传出的中心主要位于腹后侧丘脑,指向初级体感区和次级体感区。受到前脑岛和下前额脑区影响的背外侧前额叶,也将控制信号传向初级体感区。

图1 多元格兰杰因果连接度图[真针(上面),假针(下面)]。路径权值(任意单位)的相对强度分别由箭头的宽度表示。实际路径的权重分别列于表1,2。SI:初级体感区;SII:刺激体感区;AI:前脑岛;OFC:前额眶部;DLPFC:背外侧前额叶皮质;Amy:杏仁核;PAG:中脑导水管周围灰质;SMA:辅助运动区;Tha MDvc:中央背侧丘脑;Tha VPL:腹后侧丘脑;PCC:后扣带回;pC:楔前叶;Hypo:下丘脑;IPL:顶下小叶;L:左;R:右 图2 在不同的静息条件下的多重格兰杰因果连接度:针刺真穴前(顶部), 针刺假穴后(中间)和静息状态(底部)。路径权值(任意单位)的相对强度分别由箭头的宽度表示。AI:前脑岛;DLPFC:背外侧前额叶;SI:初级体感区;SII:次级体感区;OFC:前额眶部;Amy:杏仁核;PAG:中脑导水管周围灰质;SMA:辅助运动区;Tha MDvc:中央背侧丘脑;Tha VPL:腹后侧丘脑;PCC:后扣带回;pC:楔前叶;Hypo:下丘脑;IPL:顶下小叶;L:左;R:右Fig.1 Multivariate Granger causality connectivities among selected ROIs(ACUP>SHAM,P<0.005 uncorrected)in ACUP(top)and SHAM(bottom).Relative strengths of path weights(in arbitrary units)were indicated by the width of arrows.The actual path weights were tabulated in Table I and II.Abbreviations:SI:primary somatosensory cortex.SII:secondary somatosensory cortex.AI:anterior insula.OFC:orbital frontal cortex.DLPFC:dorsolateral prefrontal cortex.Amy:amygdale.PAG:periaqueductal gray.SMA:supplementary motor area.Tha MDvc:mediodorsal nucleus of thalamus.Tha VPL:ventroposterior lateral of thalamus.PCC:posterior cingulate cortex.pC:precuneus.Hypo:hypothalamus.IPL:inferior parietal lobule.L-left:R-right.Fig.2 Multivariate Granger causality connectivity under different resting conditions:PARS(top),PSRS(middle)and Rest(bottom).Relative strengths of path weights(in arbitrary units)were indicated by the width of arrows.The actual path weights were tabulated in Table III,IV and V.Abbreviations:SI:primary somatosensory cortex.SII:secondary somatosensory cortex.AI:anterior insula.OFC:orbital frontal cortex.DLPFC:dorsolateral prefrontal cortex.Amy:amygdale.PAG:periaqueductal gray.SMA:supplementary motor area.Tha MDvc:mediodorsal nucleus of thalamus.Tha VPL:ventroposterior lateral of thalamus.PCC:posterior cingulate cortex.pC:precuneus.Hypo:hypothalamus.IPL:inferior parietal lobule.L:left.R:right.
2.3 持续针刺效应下的有效连接网络
格兰杰因果分析表明:在静息状态时,凸显网络的前脑岛和其他网络节点有最多数量的因果联系(hypothalamus,PAG,MDvc and putamen);凸显网络的传出主要指向缺省模式网络里的后扣带回/楔前叶和顶下小叶,以及中央执行脑区里的背外侧前额叶,下前额脑区和辅助运动区(图2)。由凸显网络里的节点指向前脑岛的路径都呈现明显的共变关系,而由前脑岛指向中央执行脑区和缺省模式网络节点大都呈现反共变的关系。前脑岛的输入和输出信号相对平衡,可能是惟一的产生中央执行脑区和缺省模式网络交叉点的控制信号的位置。后扣带回、楔前叶作为缺省模式网络中因果导入中心,后扣带回、楔前叶被其他网络中的节点驱动,包括顶下小叶和杏仁核的内侧颞叶。值得注意的是,传入后扣带回、楔前叶最强的信号来源于前脑岛。
相较于静息状态,真针前的每个节点的因果关系的强度和指向均呈现不同的模式(图2):和6个节点均存在显著的指向关系,6个节点有显著的被指向关系。前脑岛仍然是中央执行脑区和缺省模式网络节点中心。前脑岛与其他节点的相互作用形成一些共变和异变的路径:由前脑岛传出指向中央执行脑区中的背外侧前额叶和下前额脑区,从缺省模式网络脑网络的后扣带回和顶下小叶传入指向前脑岛。除此以外,观察到前脑岛和凸显网络中的其他节点有相对增加的因果关系:下丘脑和中央背侧丘脑。在每个节点的因果影响中,由下丘脑指向中脑导水管周围灰质最强。另一方面,在假针的情况,格兰杰因果分析连接图则表现的更为稀疏。与静息情况下相比,从背外侧前额叶传到初级体感区的因果关系相对增强。在这个环路中,初级体感区有一个相对更高的传入情况,主要从后扣带回/楔前叶、背外侧前额叶和次级体感区传入。此外,和针刺真穴相比,针刺假穴在凸显网络脑网络中的前脑岛节点与其他节点之间的因果关系表现不显著。
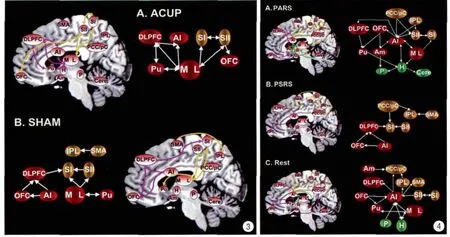
图3 针刺真穴和针刺假穴的中枢神经调节通路。因果相互作用主要包括由丘脑发出上行传导路径,以及来自额叶皮质到体感区的自上而下的控制信号。AI:前脑岛;DLPFC:背外侧前额叶;SI:初级体感区;SII:次级体感区;OFC:前额眶部;M:中央背侧丘脑;L:腹后侧丘脑;Pu:壳核;IPL:顶下小叶;SMA:辅助运动区 图4 不同静息状态下的中枢神经通路(Rest:纯静息态;PARS:针刺真穴的后静息;PSRS:针刺假穴的后静息)。PCC/pC:后扣带回/楔前叶;IPL:顶下小叶;DLPFC:背外侧前额叶;OFC:前额眶部;SII:次级体感区;AI:前脑岛;Pu:壳核;Am:杏仁核;M:中央背侧丘脑;L:腹后侧丘脑;H:下丘脑;P:中脑导水管周围灰质;Cere:小脑Fig.3 The schematic illustration refers to neural circuits involved in ACUP and SHAM.The neural substrates involving the casual interaction networks were generally the same under both conditions.Their causal interactions mainly included the ascending path from the thalamus as well as the top-down control signal from the frontal cortices to the somatosensory areas.Abbreviations:AI:anterior insula.DLPFC:dorsolateral prefrontal cortex.SI:primary somatosensory cortex.SII:secondary somatosensory cortex.OFC:orbital frontal cortex.M:mediodorsal nucleus of thalamus.L:ventroposterior lateral of thalamus.Pu:putamen.IPL:inferior parietal lobule.SMA:supplementary motor area.Fig.4 The schematic illustration refers to heterogeneous neural pathways under different resting conditions(PARS,PSRS and Rest).At least three neural circuits anchored by the AI were involved in the PARS.Neural pathway central to processing the emotion-motivation aspects(AIOFC-Am-P)(A).Antinociceptive action of the inhibitory system(AI-H-P)(B).mediating infl uence of sensory processes(AI-SII-SI)(C).By contrast,the primary pathway during the PSRS was from frontal cortices to somatosensory areas.During the Rest,the AI had the highest number of causal infl ow connections from other nodes within the SN(H,P,M and Pu),its outfl ows mainly directed to the PCC/pC and IPL within the DMN,as well as the DLPFC,OFC and SMA within the CEN.Abbreviations:PCC/pC:posterior cingulate cortex/precuneus.IPL:inferior parietal lobule.DLPFC:dorsolateral prefrontal cortex.OFC:orbital frontal cortex.SII:secondary somatosensory cortex.SI:primary somatosensory cortex.AI:anterior insula.Pu:putamen.Am:amygdale.M:mediodorsal nucleus of thalamus.L:ventroposterior lateral of thalamus.H:hypothalamus.P:periaqueductal gray.Cere:cerebellum.
3 讨论
目前的研究考察了在针刺刺激的即时效应阶段和持续性效应阶段(刺激后的动态观测用来判断因果脑网络作用关系的发展演变),在针刺的不同阶段存在不同脑区之间交互作用关系的动态重组。值得注意的是,不同脑区间的因果作用关系在针刺刺激结束后并没有减弱。相反的是,脑区间的因果作用将进一步加强并且为其在针刺不同效应阶段扮演的角色作出重要解释。针刺足三里穴包含多级神经回路,与之相关的大脑网络则更加广泛、更具时变效应。这些回路涉及编码痛情绪相关脑区和抑制伤害性信息的脑区。不同干预措施的针刺方法(真穴与旁开假穴)存在潜在的异质脑网络,并且具备随时间变化的动态重组特点。
以往的神经影像学研究着重考察针刺中枢神经响应的空间分布。较为一致的证据表明,广泛的边缘系统相关区域以及疼痛下行抑制通路成为构成针刺功能特异性的基石[5-7,13]。然而,传统方法本身的局限已无法满足针刺所涉及脑网络内部核团信息流动以及相应的动态信息重组而发挥作用的研究需求。随着fMRI在探索脑区间系统时间响应关系方面敏感性的提高,可靠的证据支持时间响应关系可以通过刻画fMRI的血液动力学方法追踪神经信号的改变来证实。通过应用fMRI来评价脑区间的协同加工机制以及利用时间测量方法来追踪这些脑区之间信号流动的方向性,格兰杰因果脑网络可作为一种有效的研究方法之一[9,11]。多元格兰杰因果分析的结果支持,针刺足三里穴与针刺假穴存在潜在不同的中枢神经作用机制,信号调制会沿着不同的中枢神经通路到达高级皮层(图3)。外周刺激输入的一个主要上升传导脊髓通路是脊髓丘脑侧束(STT)[17]。它主要投射在丘脑的躯体感觉区(VPL),从这里出发,再投射至相应的大脑皮层区域(初级体感区、次级体感区),被命名为脊髓丘脑侧束-丘脑-第一体感区、第二体感区神经传导通路。另一条主要的上行通路是沿着脊髓路径至中间丘脑核,再投射至前脑岛,被命名为脊髓-平行侧枝-前脑岛神经传导通路。脊髓丘脑侧束-丘脑-初级体感区/次级体感区对于编码识别感觉的强度和疼痛知觉的定性特征能力十分重要,推测直接通过脊髓的信息传入至边缘结构可能极大的参与了信息的自动化处理和与身体觉醒和情绪相关的行为[18]。通过探测在针刺状态下脑区的因果作用关系,预测了针刺足三里穴可能会引起从多条脊髓上升通路传导的与疼痛有关的信息(脊髓丘脑侧束-丘脑-初级体感区、次级体感区和脊髓-平行侧枝-前脑岛),而针刺假穴仅存在脊髓丘脑侧束-丘脑-初级体感区、次级体感区这条神经通路的传导。综合以上结论表明:不同条件下可能引发不同的中枢神经上行传导通路,而中枢神经加工回路所提供的自上而下的调制通路几乎是一样的。背外侧前额叶皮质对疼痛下行抑制区域与疼痛信号加工脑区的至上而下的调节通路(初级体感区和丘脑)。这些神经回路的建立出现在即时针刺阶段,更多脑区之间功能特异性的交互作用则表现在针刺持续性效应阶段。
在纯静息状态时,后扣带回成为一个网络中枢,直接影响着默认网络中的其他节点(图4)。这个结果与以往的神经影像学的发现相一致,即后扣带回可能在信息加工和默认网络中的脑区间交互作用方面占据重要地位[17]。一系列的论文研究采用无任务的、固有连接性的分析,已经表明大脑组成广泛分布的反相关功能脑网络;“缺省模式”下大脑活动的潜在脑网络,主要在静息状态时出现并且在各种有目的的任务状态时会大幅度衰减[17]。在这两种刺激干预措施实施后,后扣带回由静息状态的信息流的目的地,转变为主要向中央执行网络投射的信息源。关于这个现象的功能意义占主导地位的假设表明:在一个新的特定任务执行时,信息的处理从通常主要参与默认网络的区域转移至与执行当前任务相关的脑区。这些证据佐证一个观点:固有脑网络间的因果信息流动,在针刺干预后,向着与针刺刺激功能特异性相关的脑区发生了转移与重新分配。
因果关系图表明在针刺的持续性效应中前脑岛是信息流入与流出的交互中心节点。我们发现,杏仁核接收前额脑区的信息流入,而信息流出却投射到中央导水管周围灰质。前额下部脑区与杏仁核之间的信息流动主要完成对疼痛这种伴有显著情绪刺激的识别,进而产生对响应刺激的应对情感状态,影响疼痛中负性情绪的表达[19-20]。前额下部脑区-杏仁核通路的具体功能尚不明确,但可能反映了一种自上而下的参与针刺行为情绪调节相关的控制信号。从杏仁核到中脑导水管周围灰质的路径则表现为一个自上而下的控制信号传递。
这种显著的相互作用部分是由于杏仁核参与了伤害性信息的调制过程,可能介导传递至中脑导水管周围灰质[14]。此外,本研究结果还表明可能通过这一路线的感觉信号是由前脑岛发出而传到下丘脑,最后传至中脑导水管周围灰质。下丘脑具有极为丰富的内啡肽神经元分布并且会直接对下行抑制通路上的重要核团中脑导水管周围灰质产生调节作用,以往研究也表明这样的调节方式在针刺镇痛作用的发挥中至关重要[7,12,21-23]。中脑导水管周围灰质传递到脊髓背角,构成了下行镇痛系统输出通道[24]。 这些结构之间的因果关系表明针刺中枢神经传导通路包括了下行抑制通路系统[25-27]。
本研究借助非重复事件相关的设计模式和格兰杰因果有效连接度分析方法,考察了针刺刺激所发挥复杂中枢神经调控机制的实验现象,以及所涉及的可能中枢神经传导通路。结果表明:大脑网络应对不同的干预措施(真穴位与假穴针刺)将引发异质性脑区间的传导信息重组,特别是表现在针刺持续性效应阶段。据推测,这种重组存在随时间变化的动态过程,其中一些相关(或反相关)区域可能只在针刺的“延迟效应”阶段呈现。针刺不同作用阶段涉及不同的因果脑网络的重组对理解针刺涉及发挥复杂多系统效应提供了实验证据支持,但该初步结论还需进一步证实。
[References]
[1]Mayer DJ,Price DD,Rafi i A.Antagonism of acupuncture analgesia in man by the narcotic antagonist naloxone.Brain research,1977,121(2):368.
[2]Pomeranz B,Chiu D.Naloxone blockade of acupuncture analgesia:endorphin implicated.Life Scien,1977,19(11):1757.
[3]Price DD,Rafii A,Watkins LR,et al.A psychophysical analysis of acupuncture analgesia.Pain,1984,19(1):27-42.
[4]Bai L,Qin W,Liang J,et al.Spatiotemporal modulation of central neural pathway underlying acupuncture action:a systematic review.Current Med Imaging Review,2009,5(3):167-173.
[5]Bai L,Qin W,Tian J,et al.Detection of dynamic brain networks modulated by acupuncture using a graph theory model.Prog Nat Sci,2009,19(7):827-835.
[6]Bai L,Qin W,Tian J,et al.Acupuncture modulates spontaneous activities in the anticorrelated resting brain networks.Brain Res,2009,1279:37-49.
[7]Bai L,Qin W,Tian J,et al.Time-varied characteristics of acupuncture effects in fMRI studies.Hum Brain Mapp,2009,30(11):3445-3460.
[8]Dhond RP,Yeh C,Park K,et al.Acupuncture modulates resting state connectivity in default and sensorimotor brain networks.Pain,2008,136(3):407-418.
[9]Stilla R,Deshpande G,LaConte S,et al.Posteromedial parietal cortical activity and inputs predict tactile spatial acuity.J Neurosci,2007,27(41):11091-11102.
[10]Dhamala M,Rangarajan G,Ding M.Analyzing information fl ow in brain networks with nonparametric Granger causality.Neuroimage,2008,41(2):354-362.
[11]Wang X,Chen Y,Ding M.Estimating Granger causality after stimulus onset:a cautionary note.Neuroimage,2008,41(3):767-776.
[12]Zhou Z,Chen Y,Ding M,et al.Analyzing brain networks with PCA and conditional Granger causality.Hum Brain Mapp,2009,30(7):2197-2206.
[13]Hui KK,Liu J,Marina O,et al.The integrated response of the human cerebro-cerebellar and limbic systems to acupuncture stimulation at ST 36 as evidenced by fMRI.Neuroimage,2005,27(3):479-496.
[14]Talaraich J,Tournoux P.Co-planar stereotactic atlas of the human brain.Thieme Medical Publishers,Weinheim:Wiley-VCH,1988:437-460.
[15]Deshpande G,LaConte S,James GA,et al.Multivariate Granger causality analysis of fMRI data.Hum Brain Mapp,2009,30(4):1361-1373.
[16]Kus R,Kaminski M,Blinowska KJ.Determination of EEG activity propagation:pair-wise versus multichannel estimate.IEEE Trans Biomed Eng.,2004,51(9):1501-1510.
[17]Fox MD,Snyder AZ,Vincent JL,et al.The human brain is intrinsically organized into dynamic,anticorrelated functional networks.Proc Natl Acad Sci U S A,2005,102(27):9673-9678.
[18]Treede RD,Kenshalo DR,Gracely RH,et al.The cortical representation of pain.Pain,1999,79(2):105-111.
[19]Schoenbaum G,Saddoris MP,Stalnaker TA.Reconciling the roles of orbitofrontal cortex in reversal learning and the encoding of outcome expectancies.Ann N Y Acad Sci,2007,1121:320-335.
[20]Tolle TR,Kaufmann T,Siessmeier T,et al.Region-specifi c encoding of sensory and affective components of pain in the human brain:a positron emission tomography correlation analysis.Ann Neurol,1999,45(1):40-47.
[21]Wu MT,Hsieh JC,Xiong J,et al.Central nervous pathway for acupuncture stimulation:localization of processing with functional MR Imaging of the brain-preliminary experience.Radiology,1999,212(1):133-141.
[22]Yu LC,Han JS.Involvement of arcuate nucleus of hypothalamus in the descending pathway from nucleus accumbens to periaqueductal grey subserving an antinociceptive effect.Internat J Neuroscien,1989,48(1-2):71-78.
[23]Chen FY,Shen ZW,Guan JT,et al.Observation of the relation between brain activity and de qi sensation with manual acupuncture at LI4(Hegu).Chin J Magn Reson Imaging,2011,2(2):112-117陈凤英,沈智威,关计添,等.手法针刺合谷穴得气与脑功能激活关系的探讨.磁共振成像,2011,2(2):112-117.
[24]Vanegas H,Schaible HG.Descending control of persistent pain:inhibitory or facilitatory? Brain Res Brain Res Rev,2004,46(3):295-309.
[25]Peets JM,Pomeranz B.CXBK mice defi cient in opiate receptors show poor electroacupuncture analgesia.Nature,1978,273(5664):675-676.
[26]Ren K,Dubner R.Descending modulation in persistent pain:an update.Pain,2002,100(1-2):1-6.
[27]Takeshige C,Oka K,Mizuno T,et al.The acupuncture point and its connecting central pathway for producing acupuncture analgesia.Brain Res Bull,1993,30(1-2):53-67.
