山东省孤独症4号染色体基因扫描研究*
2014-12-06杨树林刘金同
杨树林 杨 帆 翟 静 刘金同 张 燕 陈 刚
孤独症(autism)是儿童广泛性发育障碍(pervasive developmental disorder,PDD)的一种类型,以三主征为主要表现,即言语发育障碍、人际交往障碍、兴趣狭窄和行为方式刻板。发病后患者的社会能力将受到严重影响,有较高的致残率,给社会和患者家庭带来沉重负担。近年的流行病学研究显示,孤独症的患病率比以往有明显增加,美国为4/10 000~10/10 000、欧洲为10/10 000、中国2.8/10 000 ~11/10 000,男女比例为(4 ~10)∶1[1,2]。本研究以山东省 38 个孤独症核心家系及正常对照者为研究对象,使用覆盖4号染色体的22个微卫星遗传位点进行了基因组扫描,并通过关联分析寻找孤独症的相关性位点。
1 对象和方法
1.1 对象 以国际疾病分类10版(ICD-10)为诊断标准,选择就诊于山东省精神卫生中心的孤独症核心家系,包括:患者38例,其中女性患者2例,男性患者36例,年龄为2~10岁,平均年龄(5.10±1.61)岁;患者父母75名,其中女性38名,男性37名;1 000名正常对照选自山东省血液中心的健康献血者,女性570名,男性430名,年龄为 18~53岁,平均年龄(22.2±4.45)岁。本研究得到了山东省医学科学院基础医学研究所伦理委员会的批准,对受试者均进行了知情告知。
1.2 方法
1.2.1 基因组DNA制备 采用酚-氯仿法提取外周血样本中的基因组DNA。采用nanodrop 1000微量光度计测量DNA浓度,每个样本测量两次,取平均值。用无菌去离子水将样品稀释至20 ng/μl。
1.2.2 DNA混合池(DNA pooling)的构建 每组每个样品取10 μl DNA溶液混合,分别构建患者组、对照组和患者父母组的DNA混合池。混合池DNA终浓度为20 ng/μl。
1.2.3 遗传位点的选择 选择针对4号染色体的Linkage Mapping Set2.5 套装试剂盒(Applied Biosystem,美国),共22个微卫星位点。引物的标记荧光分别为绿色、蓝色和黄色,分子量标记荧光为橙色。
1.2.4 PCR反应 使用Gene Amp 9700 DNA扩增仪(Applied Biosystem,美国)进行PCR反应。PCR反应体系总体积为 15 μl,包括:DNA 模板 1.5 μl(20 ng/μl),PrimeSTAR HS Premix(Takara)7.5 μl,等量正反向引物混合液 1 μl(5 μM),无菌去离子水 5 μl。反应程序为降落式PCR(Touch Down PCR):94℃变性90 s,63℃退火90 s(每循环一次,退火温度降低0.5℃),72 ℃延伸90 s,14个循环;94 ℃变性30 s,56℃退火90 s,72℃ 延伸90 s,30个循环;72℃延伸7 min;4℃保存。
1.2.5 毛细管凝胶电泳及基因分型 按照PCR产物荧光颜色和片段大小的不同,将PCR产物搭配组合进行电泳观察。患者组、对照组和患者父母组分别上样,取混合后的 PCR 产物 4 μl与 9 μl上样液(0.15 μl Liz 500,9 μl去离子甲酰胺)混合 ,加入96孔板中。95℃变性5 min,并迅速冰浴冷却。使用ABI-3130(Applied Biosystem,美国)毛细管电泳仪进行电泳观察。试验数据由GeneMapper4.0软件(Applied Biosystem,美国)进行分析。
1.3 统计学处理 样品经过PCR扩增和基因分型后,即可得到等位基因图谱(allele image pattern,AIP)。图谱中峰的数量为遗传位点上的等位基因数量,峰高则代表对应等位基因的染色体数量。每个混合池中染色体的数量并不相等,相同位点两组间的信号强度不可直接比较。所以,首先算出各等位基因占所有等位基因的比率,再乘以总染色体数,即可得到该等位基因在相应实验组中的染色体数量。以对照组为例,人类为二倍体核型,1 000例对照组样本在常染色体上的遗传位点由2 000个等位基因组成,将某个等位基因的比率乘以2 000即可得到相应等位基因在混合池中的染色体数量。患者组与患者父母组使用相同方法计算。采用CLUMP软件对患者组和对照组、患者组和患者父母组每个位点上的等位基因频率进行比较。两组间基因频率差异以P<0.01作为有显著性的标准。
2 结果
最终得到3个混合池样本4号染色体上22个微卫星位点的等位基因图谱。根据图谱计算各组位点中等位基因的染色体数量,采用CLUMP软件统计比较各组间等位基因频率的差异(见表1)。发现患者组与患者父母组的等位基因频率在D4S392(4q13.3)位点有显著性差异(P<0.01);患者组与对照组的等位基因频率在D4S1535(4q35.1)位点有显著性差异(P<0.01)。进行3次重复实验,结果得到一致性验证,确认此位点与孤独症呈现关联。与孤独症相关联的D4S392(4q13.3)位点和 D4S1535(4q35.1)位点的等位基因图谱见图1和图2。
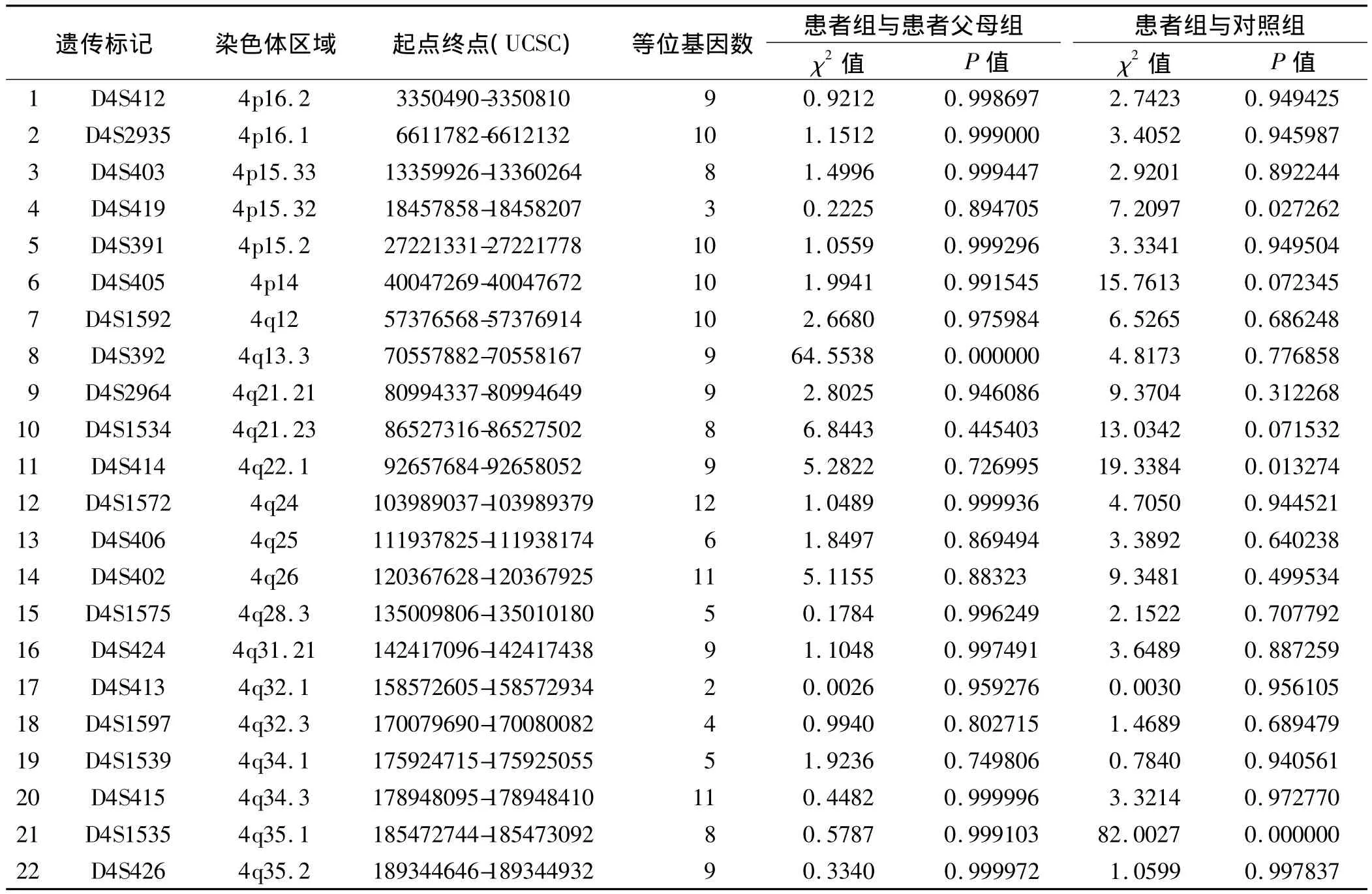
表1 孤独症患者组、患者父母组与对照组的4号染色体微卫星位点扫描关联分析结果
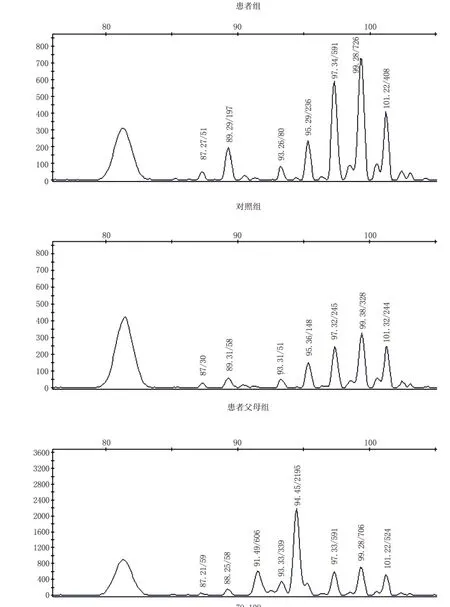
图1 孤独症患者组、患者父母组与对照组的D4S392位点基因扫描图
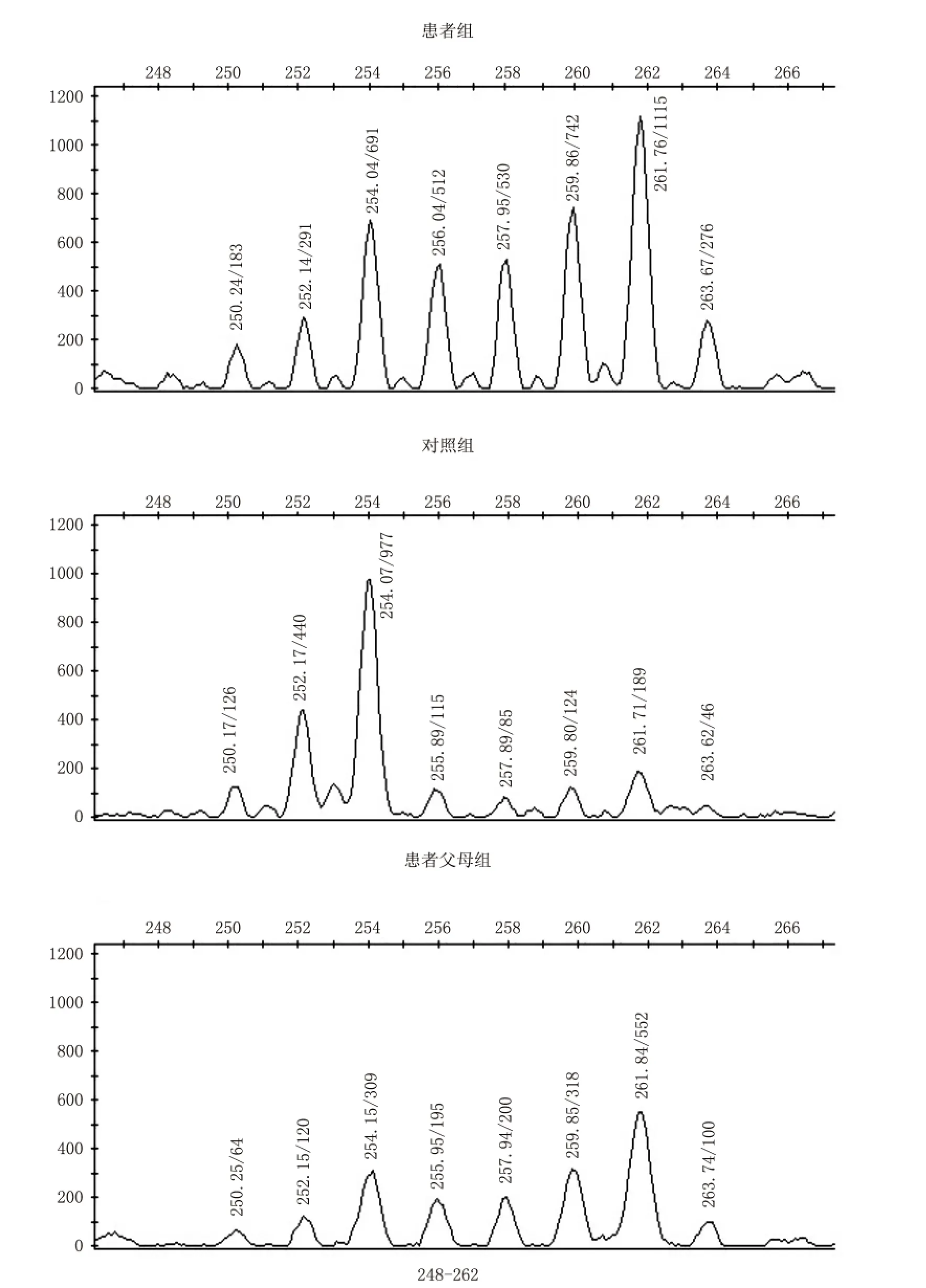
图2 孤独症患者组、患者父母组与对照组的D4S1535位点基因扫描图
3 讨论
孤独症属于多基因遗传病,有较高的遗传异质性,遗传因素在孤独症的发病过程中发挥了重要作用,其遗传度约为91%[2,3]。关联与连锁分析被较多地用于孤独症易感基因的研究。孤独症的分子遗传学研究中也常以孤独症谱系疾病(autism spectrum disorder,ASD)和广义孤独症(broad autism phenotype)作为研究对象。近20年来,国际上陆续发现了一些孤独症遗传位点。其中已确定与孤独症连锁的遗传位点和基因有24个,分别是 7q22(AUTS1)、13q14(AUTS3)、15q11(AUTS4)、2q(AUTS5)、17q11(AUTS6)、17q21(AUTS7)、3q25-q27(AUTS8)、7q31(AUTS9)、7q36(AUTS10)、1q41(AUTS11)、21p13-q11(AUTS12)、12q14(AUTS13)、16p11.2(AUTS14A 、AUTS14B)、CNTNAP2基因(AUTS15)、SLC9A9基因(AUTS16)、SHANK2基因 (AUTS17)、CHD8 基因 (AUTS18)、NLGN3基因(AUTSX1)、NLGN4 基因(AUTSX2)、MECP2 基因(AUTSX3)、Xp22.11(AUTSX4)、RPL10基因(AUTSX5)和TMLHE基因(AUTSX6)。
迄今,对4号染色体与孤独症之间的相关性也进行了一系列的关联与连锁分析。Schellenberg等[4]以微卫星为遗传学标记对孤独症患者进行了全基因组扫描研究,通过连锁不平衡分析,在有女性患者的家系中发现4号染色体上存在易感位点。Field等[5]在对有阅读障碍患者家系的关联分析中发现4q13.1、7q36.1-q36.2、7q36.3、16p12.1 和17q22 五个位点的 lod 值大于2.3,与疾病存在关联。其中4q13.1位点与儿童多动症存在关联,7q36.1-q36.2、7q36.3、16p12.1 和17q22位点与孤独症存在关联。Zuo等[6]发现4号染色体上的ADH基因是非洲裔美国人精神分裂症和欧洲裔美国人孤独症的危险因素。但是,Auranen等[7]对芬兰隔离人群中的孤独症复杂家系进行了配对连锁分析和同胞对分析中并未发现4号染色体上存在孤独症的易感性位点。
核型分析是遗传性疾病病因研究的重要方法。人们发现染色体的异位、倒位、缺失和重迭在孤独症的发生过程中发挥了重要作用,同时也是寻找孤独症易感基因的重要途径。Sabaratnam M等[8]报道了一例有dup(4)p12-p13染色体畸变的18岁女性孤独症患者。Ramanathan S等[9]报道了一例有 46,XY del 4(q31.3-q33)染色体畸变的孤独症患者,他们又通过分子遗传学的研究方法发现该患者存在AMPA2基因,GLRA3基因,GLRB基因,NPY1R基因和NPY5R基因的半合子状态,这些基因在学习、记忆能力和某些神经递质的表达中起到重要作用。Vincent JB等[10]报道了两个有46,XY,inv(4)(p12-p15.3)染色体畸变的孤独症同胞,这个畸变可能影响到了γ-氨基丁酸受体基因家族,并由此引发孤独症。为了验证γ-氨基丁酸系统对孤独症的影响,Collins AL等[11]以高加索人种和非洲裔美国人种的孤独症复杂家系进行了研究,通过对35个SNP位点的筛查,他们发现这两个人种在GABRB1基因和GABRA4基因中均存在阳性位点。Chien WH 等[12]报道了两个在 4q35.1-35.2 区域和8p23.2-pter分别存在 6.8 Mb 和 2.4 Mb 缺失的男性孤独症患者,而且这种染色体畸变在其他患者和正常对照人群中均未被发现。Shimada S等[13]报道了一例有智力障碍和孤独症症状的塞-科二氏综合征的患者,通过染色体表型分析,发现该患者在4q13.2和7p21.1区域同时存在缺失。与同样存在7p21.1区域缺失的此类患者相比,该患者的精神症状较重,所以4q13.2区域的缺失可能是导致这一现象的原因。
此外,其他研究方法也表明4号染色体上存在孤独症的易感位点。三碱基重复序列在人类DNA中广泛存在,如CCG等,研究表明这些重复序列的变化可以影响某些疾病的遗传易感性,如双向情感障碍、精神分裂症、孤独症和焦虑症。Kleiderlein JJ等[14]通过cDNA探针发现4q35区域就存在这种重复序列。在DNA拷贝数变异(copy number variants,CNVs)的研究中,Gau SS等[15]报道了一例孤独症患者在 4q12-13.1和5q32区域存在分别为4.5 Mb和1.8 Mb的CNVs,这两处变异分别来自无任何孤独症症状的患者母亲和父亲,所以可能是这两处变异的综合效应导致了孤独症的发生。
在之前的研究中,我们发现患者组与对照组的等位基因频率在D7S513(7p21.3)位点有显著性差异(P<0.01)[16]。此次对4 号染色体的研究中,我们发现患者组与患者父母组的等位基因频率在D4S392位点(4q13.3)有显著性差异(P <0.01);患者组与对照组的等位基因频率在D4S1535位点(4q35.1)有显著性差异(P<0.01)。这两个位点与孤独症的相关性在我国也是首次报道。我们计划在 D4S392和D4S1535附近挑选基因,选择SNP等高密度遗传标记,通过关联分析、构建单倍体型等方法,进一步筛选孤独症的易感基因。
[1]Folstein SE,Rosen-Sheidley B.Genetics of autism:complex aetiology for a heterogeneous disorder[J].Nat Rev Genet,2001,2(12):943-955
[2]杨树林,陈刚.孤独症遗传学研究进展[J].精神医学杂志,2009,22(1):60-65
[3]Bailey A,Le Couteur A,Gottesman I,et al.Autism as a strongly genetic disorder:evidence from a British twin study[J].Psychol Med,1995,25(1):63-77
[4]Schellenberg GD,Dawson G,Sung YJ,et al.Evidence for multiple loci from a genome scan of autism kindred[J].Mol Psychiatry,2006,11(11):979,1049-1060
[5]Field LL,Shumansky K,Ryan J,et al.Dense-map genome scan for dyslexia supports loci at 4q13,16p12,17q22;suggests novel locus at 7q36[J].Genes Brain Behav,2013,12(1):56-69
[6]Zuo L,Wang K,Zhang XY,et al.Association between common alcohol dehydrogenase gene(ADH)variants and schizophrenia and autism[J].Hum Genet,2013,132(7):735-743
[7]Auranen M,Nieminen T,Majuri S,et al.Analysis of autism susceptibility gene loci on chromosomes 1p,4p,6q,7q,13q,15q,16p,17q,19q and 22q in Finnish multiplex families[J].Mol Psychiatry,2000,5(3):320-322
[8]Sabaratnam M,Turk J,Vroegop P.Case report:autistic disorder and chromosomal abnormality 46,XX duplication(4)p12-p13[J].Eur Child Adolesc Psychiatry,2000,9(4):307-311
[9]Ramanathan S,Woodroffe A,Flodman PL,et al.A case of autism with an interstitialdeletion on 4q leadingto hemizygosity for genes encoding for glutamine and glycine neurotransmitter receptor sub-units(AMPA 2,GLRA3,GLRB)and neuropeptide receptors NPY1R,NPY5R[J].BMC Med Genet,2004,5:10
[10]Vincent JB,Horike SI,Choufani S,et al.An inversion inv(4)(p12-p15.3)in autistic siblings implicates the 4p GABA receptor gene cluster[J].J Med Genet,2006,43(5):429-434
[11]Collins AL,Ma D,Whitehead PL,et al.Investigation of autism and GABA receptor subunit genes in multiple ethnic groups[J].Neurogenetics,2006,7(3):167-174
[12]Chien WH,Gau SS,Wu YY,et al.Identification and molecularcharacterization oftwo novelchromosomal deletions associated with autism[J].Clin Genet,2010,78(5):449-456
[13]Shimada S,Okamoto N,Nomura S,et al.Microdeletions of 5.5 Mb(4q13.2-q13.3)and 4.1 Mb(7p15.3-p21.1)associated with a saethre-chotzen-like phenotype,severe intellectual disability,and autism[J].Am J Med Genet A,2013,161(8):2078-2083
[14]Kleiderlein JJ,Nisson PE,Jessee J,et al.CCG repeats in cDNAs from human brain[J].Hum Genet,1998,103(6):666-673
[15]Gau SS,Liao HM,Hong CC,et al.Identification of two inherited copy number variants in a male with autism supports two-hit and compound heterozygosity models of autism[J].Am J Med Genet B Neuropsychiatr Genet,2012,159B(6):710-717
[16]杨树林,陈刚,翟静.山东省孤独症7号染色体基因扫描研究[J].精神医学杂志,2009,22(1):1-4
