基于静息态功能磁共振低频振幅算法的创造力脑机制研究
2014-12-02梁皑莹梁碧珊张得龙焦冰倩
梁皑莹,梁碧珊,张得龙,焦冰倩,刘 鸣
(华南师范大学心理学院,广东广州510631)
一、引 言
创造力(creativity)通常是指个体能够产生同时具有新颖独创性和实用性的观点或产品的能力(Sternberg&Lubart,1999)。这种能力是人类区别于其他动物的重要特征,其在人类信息加工中发挥着重要作用(Boden,1998)。脑成像技术可以实时地在活体大脑上直接测量其在处理复杂信息时的神经活动,这为创造力神经机制的研究提供了新的途径。从发散性思维(Martindale&Hines,1975;Carlsson et al.,2000;Fink et al.,2009;Chavez-Eakle et al.,2007;Howard-Jones et al.,2005;Reverberi et al.,2005;Shamay-Tsoory et al.,2010)、艺术创造力(Solso,2001;Bhattacharya&Petsche,2005;Kowatari et al.,2009;Limb &Braun,2008)、顿悟(Aziz-Zadeh et al.,2009;Luo et al.,2003;Goel& Vartanian,2005)等不同的角度,研究者对被试在不同创造性任务刺激状态下脑区活动的变化进行分析,探讨了创造力与各个脑区功能之间的关系。由于任务设计、控制变量和基线设定的不同,创造力研究的结果比较分散,甚至出现相互矛盾(Dietrich&Kanso,2010)。时至今日,创造力的神经机制仍然是科学研究面临的重要课题。在心理学研究领域,有关创造力的影响因素、个体差异、心理机制等问题始终是心理学研究的焦点问题。
近年来,静息态功能磁功能成像技术(rs-fMRI)被广泛应用于脑功能研究中。人在静息状态下,仅占人体体重2%的大脑却消耗了全身20%的耗氧量,其中自发神经活动(spontaneous neuronal acivity)消耗了脑内大部分能量(Raichle,2006)。这暗示了大脑在静息态下进行有意义的功能活动。1995年,Biswal等人(1995)首次借助 rs-fMRI发现大脑运动皮层在低频波段存在明显的血氧水平依赖(BOLD)信号低频振荡(low frequency fluctuations,LFFs,0.01-0.08Hz),而且这一波动信号在左右半球的运动皮层保持高度的时间同步性。随后的研究也证实了自发神经活动同样存在于其他系统中,如听觉系统(Cordes et al.,2001)、视觉系统(Cordes et al.,2000)、语言系统(Hampson et al.,2002)等。这一系列发现表明大脑在静息状态下有规律的自发波动与大脑的功能存在直接关联,能够很好地体现大脑的生理—功能状态。2007年,Zang等人(2007)提出低频(0.01-0.08Hz)波动振幅(ALFF)指标,通过计算每个体素的低频振幅值的大小,可以具体地反映局部脑区自发神经活动的强弱。比率低频振幅算法(fALFF)是对ALFF方法的改进,是某个特定频段内的信号振幅强度对可探测到的全频段范围内信号幅值和的相对强度,对探测大脑自发活动具有更好的敏感性和特异性。fALFF从能量的角度反映大脑局部脑区静息态下的激活水平(Zou et al.,2008)。而且,有研究进一步指出大脑的内在活动模式与不同的频带存在关联(Zuo et al.,2010)。大脑神经元所产生的不同振荡频带是由不同的神经机制产生,具有不同的生理意义 (Buzsáki & Draguhn,2004;Engel et al.,2001;Penttonen & Buzsáki,2003)。目前,ALFF/fALFF指标已被广泛应用在临床各类疾病的病理分析中(林海龙等,2013;黄清玲等,2011;孙军等,2011;Zhang et al.,2013;Han et al.,2011)。
在创造力研究领域,Takeuchi等人(2012)应用rs-fMRI探讨创造力与默认网络(DMN)功能连接的关联,发现创造力水平与DMN内部的腹内侧前额叶(mPFC)和后扣带回(PCC)的功能连接强度显著正相关。他们提出高创造力个体静息状态下DMN内部的功能连接越强,越能有效地进行信息的整合和问题的解决。此后,邱江等人在研究中发现创造力的水平与mPFC和颞中回(MTG)的功能连接强度成显著正相关,而且两个脑区之间的功能连接的强度可以通过创造性任务的训练得到提高(Wei et al.,2013)。这些研究表明了大脑DMN自发性神经活动与创造力行为表现之间存在关联,DMN的自发神经活动对创造力具有一定的预测性。静息态功能连接是从功能子系统的层次研究多个静息态功能磁共振时间序列信号之间的相互关系,不能反映每个脑区具体的活动状态,即脑区自发性激活时信号改变的信息。为了探讨创造力与个体静息态下DMN自发神经活动的关系,我们采用fALFF这一指标,比较不同创造力水平的个体在低频波段(0.01-0.08Hz)下的神经振荡振幅的差异,以及在不同的低频频带slow-5(0.01-0.027Hz)和slow-4(0.027-0.073Hz)下神经低频振荡振幅的变化,借以进一步探讨与创造力相关的大脑神经活动机制。
二、实验材料与方法
(一)被试
招募华南师范大学一、二年级在校理科本科生180名(平均年龄18.88 ±1.05,85 男)分别进行创造力和智力测评。根据创造力测评(具体材料说明见下)中创造力的总成绩排序,筛选出最高分的22名被试组成高创造力组(11男),占测评总人数约12%;最低分的22名被试组成低创造力组(11男),占测评总人数的12%。高、低创造力组的被试参与静息态功能磁共振成像扫描。所有被试身心健康,均为右利手,视力或矫正视力正常,并在参与实验前签署了知情同意书和实验完成后获得一定的报酬。
(二)心理测评
实验选用托伦斯创造性思维测验的图形问卷(Torrance Test of Creative Thinking-figural,TTCT)对被试的创造性能力进行调查。TTCT是一套被广泛采用的创造力量表,主要测评发散性思维。发散性思维被认为是创造力的关键组成(Guilford,1967),在创造能力的测量上具有相当的效度(Kim,2008)。TTCT的图形问卷由3部分组成,第1部分是在一个蛋形线条图的基础上设想一幅图画或一个故事;第2部分是在10幅未完成的图画上画出有趣的东西;第3部分是在30对平行直线上添加线条画出不同的物件。3部分都要求被试尽可能发挥想象力,画出与众不同的答案。TTCT最终得出创造力的总分。
实验选用瑞文标准推理测验(Raven’s Standard Progressive Matrices,SPM)作为智力水平的测量材料。瑞文标准推理测量适用于一般智力的测量,具有良好的信效度(Raven,1993)。实验采用由张厚粲教授修订的中国版本,其分半信度达到0.95,间隔15天和30天的再测信度分别为0.82和0.79(张厚粲、王晓平,1989)。
(三)数据记录与处理
1.数据记录
对两组44名被试进行8分钟的静息态扫描,要求被试平卧,保持清醒,闭目平静呼吸,在扫描过程中不进行任何系统性思维运动。使用泡沫头垫固定头部并最大限度减少头部及其他部位的主动和被动运动。被试戴上专用的抗噪音耳塞,以减少扫描仪噪音对实验的影响。
实验采用3.0 T全身磁共振成像仪(Siemens Magnetom Trio Tim)及其配套头部线圈采集静息态功能成像。全脑功能像由T2*加权单次激发梯度回波的EPI序列获得,功能成像相关参数如下:脉冲重复时间(repetition time,TR)为2 000 ms,回波时间(echo time,TE)为30 ms。翻转角(flip angle)为90°,扫描视野(field of view,FOV)为 224×224 ms,平面内矩阵(acquisition matrix)为 64 ×64,层厚为 3.5 mm,体素大小(voxel size)为3.5 ×3.5 ×3.5 mm。全脑在轴位由32层组成,每个被试采集240个时间点图像。
2.数据处理
在Matlab R2011b平台上使用SPM8和DPARSF软件包进行运算。首先进行数据预处理,由于开始磁场信号的不稳定性,前10个时间点的数据不作分析,剩余230个时间点具体处理过程包括:DICOM格式转换,进行时间点对齐和头动校正,功能图像标准化(采用SPM8自带标准模版进行标准化)重采样(分辨率为3×3×3 mm3),对图像进行各向同性高斯平滑(FVHM=4 mm)以及去除线性漂移。
采用软件REST进行fALFF分析,方法概括如下:首先将每个体素的时间序列利用快速傅里叶变换(FFT)转换到频域,得到其功率谱;然后计算功率谱的平方根,将平方根所得值平均在一定的频域范围。功率谱平方根经平均后得到该体素的ALFF值。fALFF值即是某一给定的频段的ALFF与整个可测得频段的 ALFF的比值。除了低频频段(0.01-0.08Hz)外,全频带(0-0.25Hz)可分成五个不同的频带:slow-6(0-0.01 Hz)、slow-5(0.01-0.027 Hz)、slow-4(0.027-0.073 Hz)、slow-3(0.073-0.198 Hz)、slow-2(0.198-0.25 Hz)(Buzsáki& Draguhn,2004),其中 slow-6、slow-3、slow-2三个频带信号主要反应的是低频漂移信号、高频噪声以及白质信号等生理信号噪声(Zuo et al.,2010)。因此本研究只计算全脑每个体素在低频频段、slow-4和slow-5三个频带的fALFF,并将所得值进行正态化转换,得到zfALFF,以降低因个体差异造成的影响。
3.统计分析
量表分析:两组人年龄、智力及创造力得分差异由独立双样本t检验进行评估。若p<0.05,则认为两组之间有显著性差异。
低频振荡振幅组间分析:将高创造力组与低创造力组利用上述方法得到的zfALFF利用RESTV1.8软件系统进行基于体素的独立样本t检验。结果统计图经过AlphaSim校正,阈值水平为p<0.05即通过 p<0.01结合 18个体素(486 m3)达到。
不同低频频带间分析:为了进一步分析自发神经低频振幅对于不同频段的依赖性,将高创造力组的zfALFF利用RESTV1.8软件系统进行基于体素的配对样本t检验。结果统计图经过AlphaSim校正,阈值水平为p<0.05即通过p<0.001结合6个体素(162 m3)达到。
三、实验结果
(一)行为数据
高创造力组与低创造力组在TTCT成绩上具有显著差异,年龄及智力得分均无显著差异(见表1)。
(二)高、低创造力组fALFF差异
独立样本t检验结果如表2所示,高创造力组比低创造力组的低频段fALFF增强的区域有右脑岛、左运动辅助区;高创造力组比低创造力组的低频段fALFF减弱的区域有右楔前叶、左后扣带回(见图1A)。
在 slow-5(0.01-0.027Hz)频段,高创造力组比低创造力组的fALFF增强的区域有双侧颞上回、右脑岛及右额中回;高创造力组比低创造力组的fALFF减弱的区域有双侧颞下回、右枕中回、左额中回、左额上回(见图1B)。
在 slow-4(0.027-0.073 Hz)频段,高创造力组比低创造力组的fALFF增强的区域有双侧的额中回和右额下回;高创造力组比低创造力组的fALFF减弱的区域有右楔前叶、左颞中回、右中央后回(见图1C)。

表1 高创造力组和低创造力组的智力及创造力成绩独立样本t 检验结果

表2 高创造力组和低创造力组的fALFF值独立样本t检验结果
(三)slow-4和slow-5下fALFF差异
配对样本t检验结果如表3所示,高创造力组额中回和颞中回的fALFF在slow-5明显高于slow-4;尾状核的 fALFF在 slow-4明显高于slow-5(见图2)。
四、讨 论
大量研究已经证实,创造力的脑机制涉及广泛分布的脑区,这些脑区在不同的创造力任务中发挥不同的作用(Heilman et al.,2003;Moore et al.,2009;Razumnikova,2000;Razumnikova,2007)。这表明创造力是一个综合的心理活动过程,其对应的脑机制也较为复杂。本研究从静息态角度,对不同创造力个体的自发脑神经活动进行了探讨。据我们所知,这是第一个对创造力与静息态下大脑局部区域低频振幅关联性的研究。我们发现高、低创造力个体的静息态自发神经活动存在明显差异,而且fALFF体现出明显的频带特异性。
首先,在低频波段(0.01-0.08Hz),我们发现高创造力个体右脑岛和左运动辅助区的自发神经激活水平比低创造力个体显著增高,左后扣带回和右楔前叶的自发神经激活水平则显著降低。基于脑岛和运动辅助区属于突显网络(SN)的节点(Dosenbach et al.,2007;Seeley et al.,2007),后扣带回和楔前叶是 DMN的节点(Raichle et al.,2001)。本研究结果表明了在静息状态下,高创造力个体较之低创造力个体的DMN脑区的激活较弱,SN脑区的激活较强。
DMN是最受关注的大尺度脑网络之一,它的活动在执行有外界刺激的认知任务时会被抑制(去激活),而在进行自我映射和自我相关的加工过程中表现出增强(Greicius et al.,2009;Sheline et al.,2009)。Takeuchi等人(2011a)在创造力与DMN的关联性研究中发现,高创造力被试在执行与工作记忆相关的2-back任务时,DMN的重要节点楔前叶出现任务诱发的去激活(task-induced deactivation,TID)减弱,而且减弱程度与创造力水平呈显著正相关。Raichle等人(2001)提出DMN中的后扣带回和相邻的楔前叶组成一个紧张性活跃的区域,不断地采集外部以及内部的信息;在以任务为导向的信息处理过程中,这个区域通常会出现去激活。而且,后扣带回和楔前叶是语义系统的关键组成(Binder et al.,2009),对语义信息的整合处理发挥作用(Buckner et al.,2008;Jung-Beeman,2005)。在静息态下,楔前叶参与和自我相关的心理表征和情景记忆检索等认知活动(Cavanna&Trimble,2006)。据此,Takeuchi等认为创造力与DMN的TID减弱相关可能提示了高创造力个体对于与任务无关信息的抑制控制不足,从静息态下内在的语义处理过程到任务执行下认知过程的注意再分配效率不高(McKiernan et al.,2003)。本研究中高创造力个体静息态下DMN区域振幅的降低,可能表明在静息态下同样存在认知资源分配效率较低的现象(Takeuchi et al.,2011a)。高创造力个体具有低潜在抑制(low latent inhibition)的特质,他们对外面环境的刺激更敏感,他们的大脑无法屏蔽曾经出现过的同样刺激,即使对没有直接作用的事物也会进行深度分解思考(Carson et al.,2003)。

图1
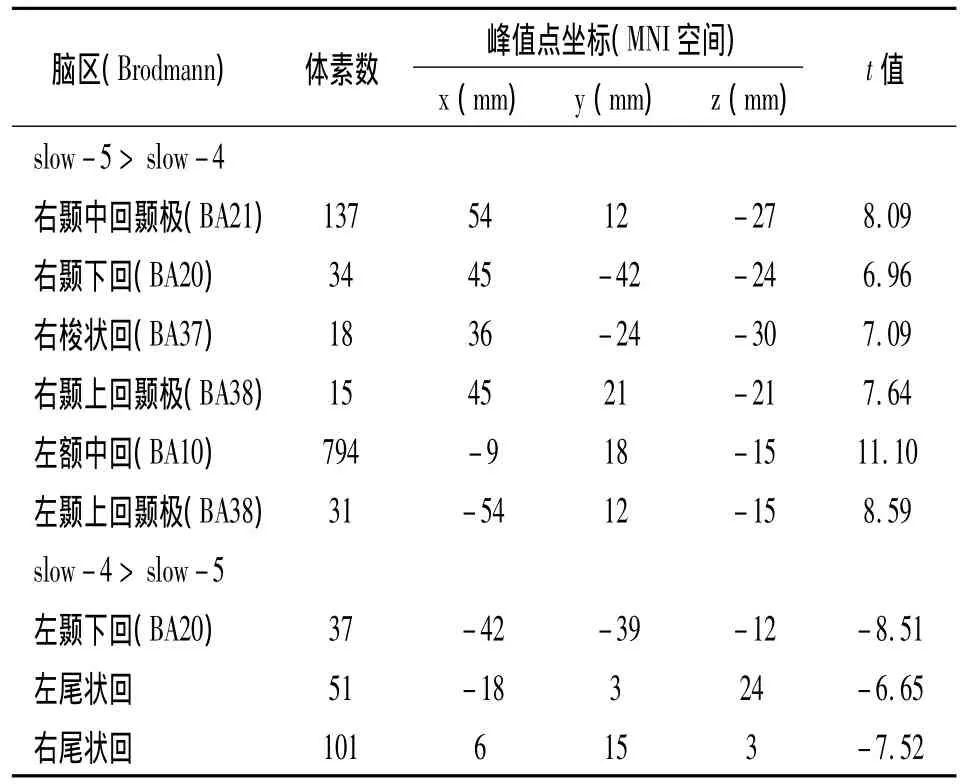
表3 高创造力组fALFF在slow-4和slow-5的配对样本t检验结果

图2
高创造力个体在静息态下DMN的局部区域激活减弱也有可能帮助高创造力个体实现两个不同网络中的概念的结合。关于创造力的概念,已有研究认为创造力是把分离的、几乎没有关联的元素进行整合 (James,1890;Spearman,1931;Guilford,1967)。而且,通过不同大脑网络间的信息联接,高创造力个体善于运用某个范畴的知识去组织形成一个新的范畴(Heilman et al.,2003)。DMN在自我意识有关的内部心理任务中激活,例如情感、情景记忆等(Buckner et al.,2008;Foster,Dastjerdi &Parvizi,2012;Andrews-Hanna et al.,2010)。它与在基于外部注意的认知任务中激活的任务正相关网络(task-positive network,TPN)相互拮抗(anticorrelation),即一个网络激活,另一个网络则去激活(Damoiseaus et al.,2006;Fox et al.,2005)。静息态下,当DMN的活动减弱,TPN的激活相应增强(Fox et al.,2005)。当一个网络对另一个网络抑制减弱,有可能使一些在去激活网络的“无关”信息进入意识,与激活网络的远距离信息结合,形成具有新颖性的想法(Takeuchi et al.,2011a)。
SN对DMN和中央执行网络(central executive network,CEN)实施动态管理(Palaniyappan&Liddle,2012)。它对不同形式的信息输入进行识别与整合,这些输入包括了体外的感知觉信息处理,以及大量的体内感受性信息的自主处理,例如痛苦情绪、饥饿、情感、奖赏等(Seeley et al.,2007)。它的关键节点右脑岛根据任务范式和刺激模式,对DMN和CEN两个网络发出激活或抑制的控制信号,使两个网络之间实现动态切换(dynamic switching)(Sridharan et al.,2008;Menon & Uddin,2010)。高创造力个体SN区域振幅的升高可能提示了高创造力个体可以感知和描绘一些隐藏于一般人的信息,更大量地接收内外部的刺激,一旦显著刺激被探查到,SN的前脑岛就会发出合适的瞬时控制信号给负责注意、工作记忆和高级认知过程的脑区,同时也抑制DMN的活动(Menon&Uddin,2010)。
本研究的另一个发现,进一步揭示了fALFF在反映与创造力有关的大脑自发波动中具有明显的频带特异性。我们发现高创造力个体自发神经活动性在slow-5和slow-4这两个不同的频带下存在显著差异。额中回和颞中回区域的fALFF在slow-5显著高于slow-4。额中回和颞中回属于DMN的节点,区域间存在结构上和功能上的神经连接(Greicius et al.,2003;Raichle et al.,2001)。已有研究指出,不同频段相比之下,较低频频段的神经振荡振幅有利于神经网络的功能整合(Buzsáki&Draguhn,2004)。与此相一致,本研究结果表明了DMN的区域自发神经活动性在较低的频段更强。本研究也发现高创造力个体的基底核(尾状核)在slow-4自发神经激活更强,这个结果与Zuo等人(2010)比较slow-4和slow-5的全脑fALFF的研究结果一致。本研究结果支撑大脑神经元在不同频段的低频振荡与不同的神经机制和生理功能存在联系的说法(Buzsáki& Draguhn,2004;Penttonen&Buzsaki,2003),并提示slow-5频段对于发现与创造力相关的DMN自发性神经低频振幅更敏感。
五、结 论
本研究通过静息态下fALFF比较了不同创造力个体的大脑自发波动特征,我们发现DMN区域神经活动减弱、SN区域神经活动增强是高创造力个体脑神经活动的一个重要特征,这一特征可能与创造力的脑机制具有密切关系。对这一神经活动特征的进一步揭示有助于提升我们对创造力脑机制的认识。同时,本研究也表明fALFF在大脑创造力有关的神经活动特征研究中具有较高的敏感性,可以有效地反应创造力有关的神经活动。
[1]黄清玲,唐勇,王沛弟,等.创伤后应激障碍患者杏仁核为主的边缘系统静息态脑功能磁共振研究.中国神经精神疾病杂志,2011,37(12):705—709.
[2]林海龙,薛蕴菁,康正武,等.静息态fMRI在无痴呆型血管性认知障碍基线脑活动变化中的初步研究.临床放射学杂志,2013,32(8):1070—1074.
[3]刘虎,范国光,徐克,等.低频振幅fMRI评价精神分裂症患者静息状态下脑功能活动.中国医学影像技术,2010(9):1659—1662.
[4]齐印宝,傅先明,王昌新,等.海洛因依赖者低频振幅算法功能性磁共振研究.中华行为医学与脑科学杂志,2011,20(2):119—121.
[5]孙军,刘含秋,孙华平,等.首发抑郁症患者治疗前后的静息态fMRI研究.中国医学计算机成像杂志,2011(3):212—216.
[6]张厚粲,王晓平.瑞文标准推理测验在我国的修订.心理学报,1989(2):113—121.
[7]J.R.Andrews-Hanna.The Brain's Default Network and Its Adaptive Role in Internal Mentation.Neuroscientist,2012,18(3):251—270.
[8]L.Aziz-Zadeh,J.T.Kaplan,M.Lacoboni.Aha!:The Neural Correlates of Verbal Insight Solutions.Human Brain Mapping,2009,30:908—916.
[9]J.R.Binder,R.H.Desai,W.W.Graves,et al.Where Is the Semantic System?A Critical Review and Meta-analysis of 120 Functional Neuroimaging Studies.Cereb.Cortex,2009,19(12):2767—2796.
[10]S.Berthoz,E.Artigues,P.-F.VandeMoortele,et al.Effect of Impaired Recognition and Expression of Emotions on Frontocingulate Cortices:An fMRI Study of Men with Alexithymia.Am J Psychiatry,2002,159(6):961—967.
[11]J.Bhattacharya,H.Petsche.Drawing on Mind’s Canvas:Differences in Cortical Integration Patterns between Artists and Non-artists.Human Brain Mapping,2005,26:1—14.
[12]B.B.Biswal,F.Z.Yetkin,V.M.Haughton,et al.Functional Connectivity in the Motor Cortex of Resting Human Brain Using Echoplanar MRI.Magn Reson Med,1995,34(4):537—541.
[13]M.A.Boden.Creativity and Artificial Intelligence.Artificial Intelligence,1998,103:347—356.
[14]R.L.Buckner,A.Z.Snyder,B.J.Shannon,et al.Molecular,Structural and Functional Characterization of Alzheimer’s Disease:Evidence for a Relationship between Default Activity, Amyloid and Memory. J Neurosci,2005,25(34):7709—7717.
[15]G.Buzsáki,A.Draguhn.Neuronal Oscillations in Cortical Networks.Science,2004,304(5679):1926.
[16]S.J.Broyd,C.Demanuele,S.Debener,et al.Defaultmode Brain Dysfunction in Mental Disorders:A Systematic Review.Neuroscience& Biobehavioral Reviews,2009,33(3):279—296.
[17]I.Carlsson,P.E.Wendt,J.Risberg.On the Neurobiology of Creativity:Differences in Frontal Activity between High and Low CreativeSubjects. Neuropsychologia,2000,38:873—885.
[18]S.H.Carson,J.B.Peterson,D.M.Higgins.Decreased Latent Inhibition Is Associated with Increased Creative A-chievement in High-functioning Individuals.J Pers Soc Psychol,2003,85(3):499—506.
[19]A.E.Cavanna,M.R.Trimble.The Precuneus:A Review of Its Functional Anatomy and Behavioural Correlates.Brain,2006,129:564—583.
[20]R.Chavez-Eakle,A.Graf-Guerrero,J.Garcia-Reyna,et al.Cerebral Blood Flow Associated with Creative Performance:A Comparative Study.NeuroImage,2007,38:519—528.
[21]E.G.Chrysikou,S.L.Thompson-Schill.Dissociable Brain States Linked to Common and Creative Object Use.Human Brain Mapping,2011,33:665—675.
[22]D.Cordes,V.M.Haughton,K.Arfanakis,et al.Mapping Functionally Related Regions of Brain with Functional Connectivity MR Imaging.AJNR Am J Neuroradiol,2000,21(9):1636—1644.
[23]D.Cordes,V.M.Haughton,K.Arfanakis,et al.Frequencies Contributing to Functional Connectivity in the Cerebral Cortex in“Resting-state”Data.AJNR Am J Neuroradiol,2001,22(7):1326—1333.
[24]J.S.Damoiseaux,S.A.Rombouts,F.Barkhof,et al.Consistent Resting-state Networks Across Healthy Subjects.Proc Natl Acad Sci U S A,2006,103(37):13848—13853.
[25]A.Dietrich.The Cognitive Neuroscience of Creativity.Psychonomic Bulletin & Review,2004,11:1011—1026.
[26]A.Dietrich,R.Kanso.A Review of EEG,ERP,and Neuroimaging Studies of Creativity and Insight.Psychological Bulletin,2010,136(5):822—848.
[27]N U.Dosenbach,D A.Fair,F M.Miezin,et al.Distinct Brain Networks for Adaptive and Stable Task Control in Humans.Proc Natl Acad Sci U S A,2007,104(26):11073—11078.
[28]S.Durston,B J.Casey.What Have We Learned about Cognitive Development from Neuroimaging?Neuropsychologia,2006,44:2149—2157.
[29]A.Fink,R.H.Grabner,M.Benedek,et al.The Creative Brain:Investigation of Brain Activity during Creative Problem Solving by means of EEG and fMRI.Human Brain Mapping,2009,30:734—748.
[30]A.Fink,R H.Grabner,D.Gebauer,et al.Enhancing Creativity by Means of Cognitive Stimulation:Evidence from an fMRI Study.Neuroimage,2010,52:1687—1695.
[31]A.Fink,K.Koschutnig,M.Benedek,et al.Stimulating Creativity Via the Exposure to Other People’s Ideas.Human Brain Mapping,2012,33:2603—2610.
[32]E.Fisher,A.Mohanty,J.D.Herrington,et al.Neuropsychological Evidence for Dimension Schizotypy:Implications for Creativity and Psychopathology.J Res Pers,2004,38:24—31.
[33]A.W.Flaherty.Frontotemporal and Dopaminergic Control of Idea Generation and Creative Drive.J Comp Neurol,2005,493(1):147—153.
[34]B.Folley,S.Park.Verbal Creativity and Schizotypal Personality in Relation to Prefrontal Hemispheric Laterality:A Behavioral and Nearinfrared Optical Imaging Study.Schizophrenia Research,2005,80:271—282.
[35]B.L.Foster,M.Dastjerdi,J.Parvizi.Neural Populations in Human Posteromedial Cortex Display Opposing Responses During Memory and Numerical Processing.Proceedings of the National Academy of Sciences of the United States of America,2012,109(38):15514—15519.
[36]M.D.Fox,A.Z.Snyder,J.L.Vincent,et al.The Human Brain Is Intrinsically Organized into Dynamic,Anticorrelated Functional Networks.Proceedings of the National Academy of Sciences of the United States of America,2005,102(27):9673—9678.
[37]M.D.Fox,M.E.Raichle.Spontaneous Fluctuations in Brain Activity Observed with Functional Magnetic Resonance Imaging.Nature Rev Neurosci,2007,8:700—711.
[38]C.Gibson,B.S.Folley,S.Park.Enhanced Divergent Thinking and Creativity in Musicians:A Behavioral and Near-infrared Spectroscopy Study.Brain and Cognition,2009,69:162—169.
[39]V.Goel,O.Vartanian.Dissociating the Roles of Right Ventral Lateral and Dorsal Lateral Prefrontal Cortex in Generation and Maintenance of Hypotheses in Set-shift Problems.Cerebral Cortex,2005,15:1170—1177.
[40]M.D.Greicius,B.Krasnow,A.L.Reiss,et al.Functional Connectivity in the Resting Brain:A Network Analysis of the Default Mode Hyothesis.Proc Natl Acad Sci U S A,2003,100(1):253—258.
[41]M.D.Greicius,K.Supekar,V.Menon,et al.Restingstate Functional Connectivity Reflects Structural Connectivity in the Default Mode Network.Cereb Cortex,2009,19(1):72—78.
[42]J.P.Guilford.The Nature of Human Intelligence.New York,NY:McGraw-Hill,1967.
[43]R.E.Gur,M.E.Calkins,R.C.Gur,et al.The Consortium on the Genetics of Schizophrenia:Neurocognitive Endophenotypes.Schizophrenia Bulletin,2007,33:49—68.
[44]M.Hampson,B.S.Peterson,P.Skudlarski,et al.Detection of Functional Connectivity Using Temporal Correlations in MR Images.Human Brain Mapping,2002,15(4):247—262.
[45]K.M.Heilman,S.E.Nadeau,D.O.Beversdorf.Creative Innovation:PossibleBrain Mechanisms. Neurocase,2003,9:369—379.
[46]I.Falkai,P.Falkai,O.Gruber.A Systematic fMRI Investigation of the Brain Systems Subserving Different Working Memory Compon Ents in Schizophrenia.Eur J Neurosci,2009,30:693—702.
[47]P.A.Howard-Jones,S-J.Blakemore,E.A.Samuel,et al.Semantic Divergence and Creative Story Generation:An fMRI Investigation.Brain Res Cogn Brain Res,2005,25:240—250.
[48]W.James.The Principles of Psychology.Holt,New York,1890.
[49]R.E.Jung,J.M.Segall,H.J.Bockholt,et al.Neuroanatomy of Creativity.Human Brain Mapping,2010,31:398—409.
[50]M.Jung-Beeman.Bilateral Brain Processes for Comprehending Natural Language.Trends in Cognitive Sciences,2005,9(11):512—518.
[51]J.Kasof.Creativity and Breadth of Attention.Creativity Res J,1997,10:303—315.
[52]K.H.Kim.Can We Trust Creativity Tests?A Review of the Torrance Tests of Creative Thinking(TTCT).Creativity Research Journal,2006,18(1):3—14.
[53]V.Kiviniemi,J.Jauhiainen,O.Tervonen,et al.Slow Vasomotor Fluctuation in fMRI of Anesthetized Child Brain.Magn Reson Med,2000,44(3):373—378.
[54]Y.Kowatari,S.H.Lee,H.Yamamura,et al.Neural Networks Involved in Artistic Creativity.Human Brain Mapping,2009,30:1678—1690.
[55]R.Lubow.Construct Validity of the Animal Latent Inhibition Model of Selective Attention Deficits in Schizophrenia.Schizophr Bull,2005,31:139—153.
[56]Q.Luo,C,Perry,D,Peng,et al.The Neural Substrate of Analogical Reasoning:An fMRI Study.Cogn Brain Res,2003,17:527—534.
[57]C.Martindale,D.Hines.Creativity and Cortical Activation during Creative,Intellectual and EEG Feedback Tasks.Biological Psychology,1975,3(2):71—80.
[58]K.A.McKiernan,J.N.Kaufman,J.Kucera-Thompson,et al.A Parametric Manipulation of Factors Affecting Task-induced Deactivation in Functional Neuroimaging.J Cogn Neurosci,2003,15:394—408.
[59]V.Menon,L.Q.Uddin.Saliency,Switching,Attention and Control:A Network Model of Insula Function.Brain Struct Funct,2010,214(5—6):655—667.
[60]B.Miller,K.Boone,J.L.Cummings,et al.Functional Correlates of Musical and Visual Ability in Frontotemporal Dementia.The British Journal of Psychiatry,2000,176(5):458—463.
[61]E.Necka.Creativity and Attention.Pol Psychol Bull,1999,30:85—98.
[62]M.Neuper,C.F.Ebner,A.C.Neubauer.The Creative Brain:Investigation of Brain Activity During Creative Problem Solving by Means of EEG and fMRI.Hum Brain Mapp,2009,30:734—748.
[63]L.Palaniyappan,P.E.Liddle.Does the Salience Network Play a Cardinal Role in Psychosis?An Emerging Hypothesis of Insular Dysfunction.J Psychiatry Neurosci,2012,37(1):17—27.
[64]M.Penttonen,Buzsáki,György.Natural Logarithmic Relationship between Brain Oscillators.Thalamus Relat Syst,2003,2:145—152.
[65]F.E.Polli,J.J.S.Barton,K.N.Thakkar,et al.Reduced Error-related Activation in Two Anterior Cingulate Circuits is Related to Impaired Performance in Schizophrenia.Brain,2008,131:971—986.
[66]M.E.Raichle,A.M.MacLeod,A.Z.Snyder,et al.A Default Mode of Brain Function.Proc Natl Acad Sci U S A,2001,98(2):676—682.
[67]J.Raven.Manual for Raven’s Progressive Matrices and Vocabulary Scales.Oxford:Oxford Psychologists Press,1993.
[68]C.Reverberi,A.Toraldo,S.D’Agostini,et al.Better without(lateral)Frontal Cortex?Insight Problems Solved by Frontal Patients.Brain,2005,128(12):2882—2890.
[69]W.W.Seeley,V.Menon,A.F.Schatzberg,et al.Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control.J Neurosci,2007,27(9):2349—2356
[70]W.W.Seeley,B.R.Matthews,R.K.Crawford,et al.Unravelling Bolero:Progressive Aphasia,Transmodal Creativity and the Right Posterior Neocortex. Brain,2008,131:39—49.
[71]S.G.Shamay-Tsoory,N.Adler,J.Aharon-Peretz,et al.The Origins of Originality:The Neural Bases of Creative Thinking and Originality.Neuropsychologia,2011,49:178—185.
[72]Y.I.Sheline,D.M.Barch,J.L.Price,et al.The Default Mode Network and Self-referential Processes in Depression.Proc Natl Acad Sci U S A,2009,106(6):1942—1947.
[73]R.Solso.Brain Activities in a Skilled Versus a Novice Artist:An fMRI Study.Leonardo,2001,34:31—34.
[74]C.D.Spearman.Creative Mind.Macmillan,London,1931.
[75]D.Sridharan,D.Levitin,V.Menon.A Critical Role for the Right Fronto-insular Cortex in Switching between Central-executive and Default-mode Networks.PNAS,2008,105(34):12569—12574.
[76]R.J.Sternberg,T.I.Lubart.The Concept of Creativity:Prospects and Paradigms∥ R.J.Sternberg(Ed.),Handbook of Creativity.New York:Cambridge University Press,1999:3—15.
[77]H.Takeuchi,et al.The Association between Resting Functional Connectivity and Creativity.Cerebral Cortex,2012,22(12):2921—2929.
[78]H.Takeuchi,Y.Taki,Y.Sassa,et al.Regional Gray Matter Volume of Dopaminergic System Associate with Creativity:Evidence from Voxel-based Morphometry.NeuroImage,2010a,51(2):578—585.
[79]H.Takeuchi,et al.White Matter Structures Associated with Creativity:Evidence from Diffusion Tensor Imaging.NeuroImage,2010,51(1):11—18.
[80]H.Takeuchi,Y.Taki,H.Hashizume,et al.Failing to Deactivate:The Association between Brain Activity during a Working Memory Task and Creativity.NeuroImage,2011a,55(2):681—687.
[81]H.Takeuchi,Y.Taki,Y.Sassa,et al.Working Memory Training Using Mental Calculation Impacts Regional Gray Matter of the Frontal and Parietal Regions.PLoS ONE,2011b,6(8):e23175.
[82]R.Toro,P.T.Fox,T.Paus.Functional Coactivation Map of the Human Brain.Cereb Cortex,2008,18(11):2553—2559.
[83]D.Wei,J.Yang,W.Li,et al.Increased Resting Functional Connectivity of the Medial Frontal Cortex in Creativity by Means of Cognitive Simulation.Cortex,2013,51:92—102.
[84]T.P.White,V.Joseph,E.O’Regan,et al.Alphagamma Interactions Are Disturbed in Schizophrenia:A Fusion of Electroence Phalography and Functional Magnetic Resonance Imaging. Clin Neurophysiol,2010,121:1427—1437.
[85]S.F.Witelson,D.L.Kigar,T.Harvey.The Exceptional Brain of Albert Einstein.The Lancet,1999,353(19):2149—2153.
[86]Y.F.Zang,Y,He,C.Z,Zhu,et al.Altered Baseline Brain Activity in Children with ADHD Revealed by Resting State Functional MRI.Brain Dev,2007(29):83—91.
[87]D.Zhang,B.Liu,J.Chen,et al.Determination of Vascular Dementia Brain in District Frequency Bands with Whole Brain FunctionalConnectivityPatterns.PLos ONE,2013,8(1):e54512.
[88]Q.H.Zou,C.Z.Zhu,Y.Yang,et al.An Improved Approach to Detection of Amplitude of Low-Frequency Fluctuation(ALFF)for Resting-State fMRI:Fractional ALFF.J Neurosci Methods,2008,172(1):137—141.
[89]X.N.Zuo,A.D.Martino,C.Kelly,et al.The Oscillating Brain:Complex and Reliable.NeuroImage,2010,49(2):1432—1445.
