内蒙古产胎生蜥蜴染色体研究*
2014-12-02赵文阁
刘 恒,赵文阁,刘 鹏
(哈尔滨师范大学)
0 引言
胎生蜥蜴(Lacerta vivipara)是蜥蜴科卵胎生动物的典型代表,在我国分布于黑龙江、内蒙古和新疆等地[1].在国外分布较广泛,从欧洲乌拉山远端的英格兰和爱尔兰,往东通过西伯利亚到远离东海岸的库页岛等广大地区[2].作为一个广泛分布的物种,胎生蜥蜴在染色体组型、性别决定机制上存在一定的地理变异[3].通过对内蒙古地区胎生蜥蜴种群染色体组型的研究,可以与大、小兴安岭地区胎生蜥蜴种群的染色体组型进行比较[1,2],为胎生蜥蜴的起源和物种演化提供参考.
1 材料与方法
2013年5月,在内蒙古的呼伦贝尔地区采集胎生蜥蜴雌雄各5只,采用骨髓细胞蒸汽固定法制备染色体标本[1,2].具体操作方法为:将胎生蜥蜴断指放血,2 h后腹腔处注射0.05%秋水仙素0.12 mL,12 h后处死,取出肱骨和股骨,剔除上面的肌肉,横断两端骨骺.用0.46%KCl溶液将骨髓细胞冲到载玻片上,将载玻片放到玻璃皿中的玻璃棒上低渗处理30 min.吸出低渗液后加入固定液(无水乙醇:冰醋酸:双蒸水 =1∶2∶3)固定2 h,换无水乙醇进行二次固定30 min,再用无水乙醇:冰醋酸=1∶2冲洗.待自然干燥后,用吉姆萨染液进行染色30 min,自来水冲洗干净后,风干装片[4].
使用Leica DC 500显微镜油镜进行观察、统计染色体数目,选取其中20个较好的中期分裂相进行照相.观察雌雄各120个分散良好、边缘清晰且较平整的中期细胞分裂相,确定二倍体染色体数目.用Leica QwinV3软件对染色体进行测量,计算出相对长度和臂比值.
2 结果
如图1、如图2所示,雄性胎生蜥蜴二倍体染色体个数是36,性染色体为ZZ型,2n=34+ZZ.雌性染色体中存在异型的性染色体,二倍体染色体个数为35,性染色体为W型,2n=34+W.除雌性W性染色体为亚端部着丝点(ST)外,全部染色体均为顶端着丝点(T),所以很难根据着丝点位置区分染色体.
根据胎生蜥蜴染色体形态与大小,17对常染色体可分成2组,第1组包括第1~4对染色体(9% >相对长度>7%),其中第1对染色体相对长度明显大于第2对染色体(F=18.80,P<0.05),容易区分,第2~4对染色体相对长度差也较大,大于0.4%,较易于辨别;第2组包括第5~17对染色体(相对长度<7%),第5~9对染色体相对长度在5% ~7%,第10~13对染色体的相对长度在4% ~5%之间,第14~17对染色体的相对长度在3% ~4%之间.Z染色体的相对长度为5.47±0.08%,在第8对与第9对常染色体相对长度之间,W染色体相对长度为5.40±0.26%,臂比值为5.47,为亚端部着丝,在第8对与第9对常染色体相对长度之间(见表1).

图1 雄性胎生蜥蜴染色体中期分裂相及组型
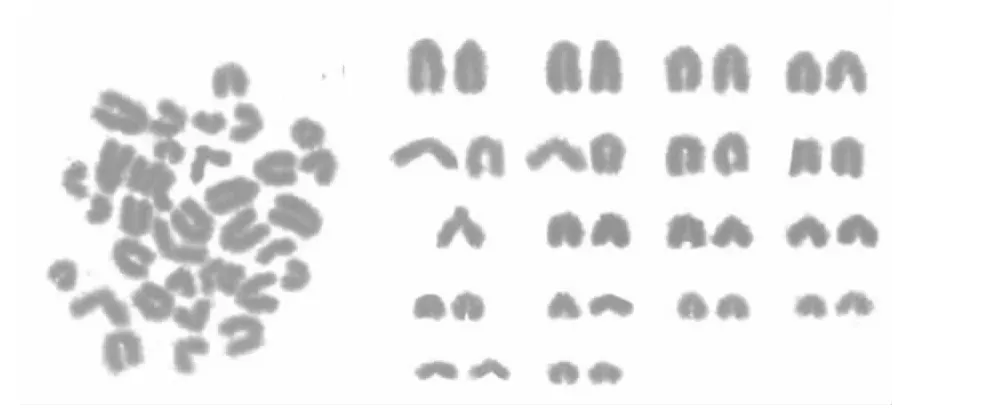
图2 雌性胎生蜥蜴染色体中期分裂相及组型
3 讨论
胎生蜥蜴分布比较广泛,具有卵生与卵胎生两种生殖模式,对其染色体组型的研究比较早[2],且不同地理种群中具有一定的染色体变异[3].内蒙古地区胎生蜥蜴种群染色体数目为雄性2n=36,雌性2n=35,W染色体为亚端着丝染色体,与乌克兰跨喀尔巴阡山脉种群[5]、匈牙利东部种群[6]、俄罗斯部分地区种群、爱沙尼亚种群、芬兰东部种群、瑞典北部种群相似[7],但W染色体类型不同,例如保加利亚种群的W染色体为亚中着丝点[8]、阿尔卑斯山种群W染色体为中部着丝点[5],此外,染色体的数目也存在一定的变化,例如斯洛维尼亚的种群发现染色体个数为雄性2n=36,雌性2n=36,性别决定机制为ZZ/ZW[9],这种同一物种不同种群的染色体数目的地理变异是物种进化的结果,与新物种的形成有关.
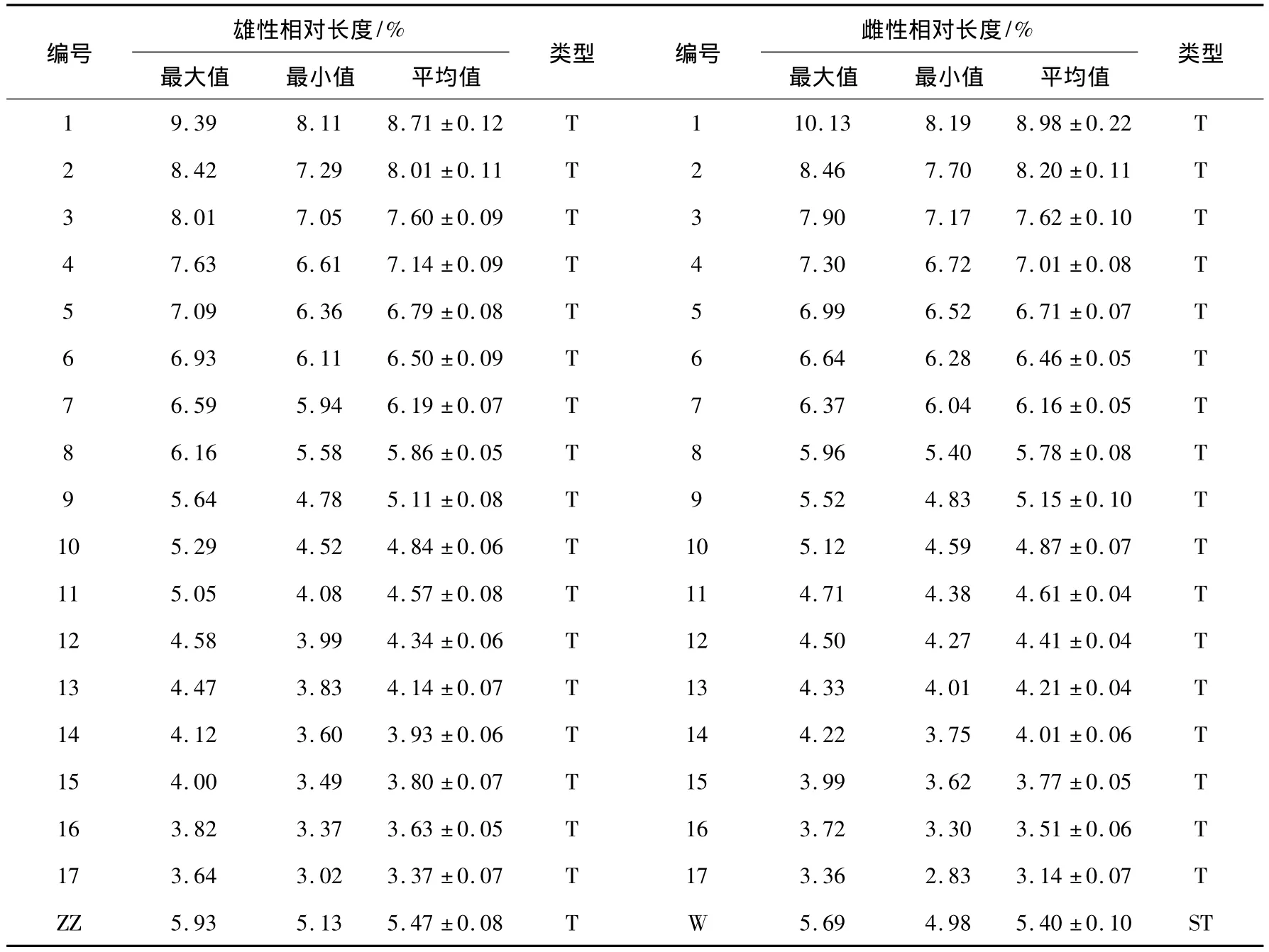
表1 胎生蜥蜴染色体相对长度
研究结果表明,内蒙古地区地处大兴安岭的西坡,胎生蜥蜴种群的W染色体类型为亚端部着丝点[1],不同于大兴安岭东坡的呼玛种群W性染色体的中部着丝[2],而与小兴安岭地区的种群一致,这三个种群W染色体类型的变化对讨论胎生蜥蜴的起源和进化具有重要意义.
[1] 朱丹,赵文阁,刘鹏,等.大兴安岭产胎生蜥蜴的染色体组型研究[J].中国农学通报,2011,27(22):75-77.
[2] 赵文阁,夏玉国,刘琳琳,等.黑龙江省产胎生蜥蜴的染色体组型研究[J].四川动物,2006,25(2):234-237.
[3] ЛА Куп риянова, ЕР Руди. Сравнигельно Карио логическийАна лиз попу ляций жив ордящей ящерицы(L.vivipara)[J].Зоо логиче ский журнал,1990,69(6):93-101.
[4] 吴政安.两栖类骨髓细胞的染色体标本制作法[J].遗传,1982,4(1):38-39.
[5] Odierna G,Aprea G,Capriglione T,et al.Progressive differentiation of the W sex chromosome between oviparous and viviparous populations of Zootoca vivipara[J].Italiana Journal of Zoology,1998,65:295-302.
[6] Puky M,Adam H,Surget-Groba Y,et al.Fajvedelmi programok letjogosultsaga es feladatai Magyarorszagon:az elevenszulo gyik(Zootoca vivipara Mayer& Bischoff,1996)vizsgalatanakeredmenyei es tanulsagai[J].Termeszetvedelmi Kozlemenyek,2004,11:411-418.
[7] Kupriyanova L A,Kuksin A,Odierna G.Karyotype,chromosome structure,reproductive modalities of three Southern Eurasian populations of the common lacertid lizard,Zootoca vivipara[J].Acta Herpetologica,2008,3(2):99-106.
[8] Belcheva R,Biserkov V,Ivlieva H,et al.Karyological studies on Lacerta vivipara collected in Bulgaria[J].Cytologia,1986,3:567-570.
[9] Odierna G,Heulin B,Guillaume C P,et al.Evolutionary and biogeographical implications of the karyological variations in the oviparous and viviparous forms of Lacerta(Zootoca)vivipara[J].Ecography,2001,24:332-340.
