烟草种质材料TSNA含量的关联分析
2014-11-24余义文夏岩石李荣华吕永华郭培国邱妙文赵伟才何其芳
余义文,夏岩石,李荣华,吕永华,郭培国,邱妙文,赵伟才,何其芳
1 广州大学生命科学学院,广州 510006;
2 广东省烟草专卖局(公司),广州 510610;
3 广东省烟草公司南雄科学研究所,南雄 512400
农艺与调制
烟草种质材料TSNA含量的关联分析
余义文1,夏岩石1,李荣华1,吕永华2,郭培国1,邱妙文3,赵伟才3,何其芳1
1 广州大学生命科学学院,广州 510006;
2 广东省烟草专卖局(公司),广州 510610;
3 广东省烟草公司南雄科学研究所,南雄 512400
为寻找与烟叶TSNA 含量显著关联的标记位点,利用分布于烟草24个染色体具多态性的SSR标记和MFLP标记,分析24份烟草种质材料的遗传多样性,在此基础上对烟叶中烟草特有亚硝胺(TSNA)含量的表型变异与标记多态性进行关联分析。结果显示,33对MFLP引物和28对SSR引物在24份烟草材料中共发现188个多态位点;群体结构分析将24份烟草材料分为3个亚群,且亚群划分与烟草类型(烤烟、晾晒烟和白肋烟)基本一致;关联分析发现1个SSR位点和6个MFLP位点至少与1种TSNA含量的相关性在0.01水平上显著,其中标记MFLP26与烤后烟叶中NNN含量的相关性在0.001水平上显著,表型变异解释率最高(R2=0.5831)。这些显著关联的分子标记可为筛选低TSNA含量的烟草材料提供参考。
烟草特有亚硝胺;SSR标记;MFLP标记;关联分析
烟草特有亚硝胺(Tobacco Specific Nitrosamines,TSNA)是烟草及其制品中的特有成分,主要包括N-亚硝基去甲基烟碱(NNN)、4-(N-亚硝基甲基氮)-1-(3-吡啶基)-1-丁酮(NNK)、N-亚硝基假木贼碱(NAB)和N-亚硝基新烟草碱(NAT)四种类型[1-2]。大量研究认为TSNA是一类具有致癌性的化合物[3-4],其中NNK和NNN被指定为一类(Class 1)致癌物,可对小鼠、大鼠及叙利亚金田鼠等动物诱发肺癌;而NAB和NAT由于对实验动物的致癌性的证据较少,相关资料不充分,所以它们的致癌性尚不能确定[5]。
卷烟烟叶原料中的TSNA是烟气中TSNA的主要来源之一[6-7]。不同烟草种质间总TSNA含量不同,且不同形式的TSNA在每种烟草所占比例也不同[8-12]。相同的栽培和调制条件下,不同类型烟草中TSNA的含量存在着种属特异性且差异显著,如烤烟烟叶中无论是叶片还是叶脉,其TSNA水平都要低于白肋烟[10-11],不同烤烟品种及不同白肋烟品种间TSNA含量与基因型显著相关[13-15]。这些研究结果表明,TSNA含量与烟草品种本身的遗传特性明显相关。因此,选育出低TSNA含量的烟草品种,可降低卷烟对人体健康的危害程度。
我国在这方面的研究刚刚起步,具有很大的潜力。烟叶生产中烟碱转化时常存在,需加强对烟碱转化规律的研究,更为重要的是研究这些TSNA及相关前体的遗传规律,采取遗传选择方法对品种进行改良,是降低我国烟叶TSNA含量的一种有效途径[16-18]。如近年来我国采用常规育种方法对白肋烟主栽品种鄂烟1号杂交种的亲本进行遗传改良,并配制出改良杂交种,新品种烟叶的烟碱转化率、降烟碱含量和TSNA含量均大幅度降低,有效地提高了白肋烟的安全性[18]。这些结果更进一步证明烟草中的TSNA含量受主效基因控制,可以在了解其遗传规律的条件下选育出系列符合优质低害要求的烤烟新品种。
到目前为止,尚未见我国开展烤烟TSNA遗传规律方面的研究工作,亦未见有从事适用于TSNA含量的分子标记开发的研究报道。本实验在SSR(Simple Sequence Repeat)标记和 MFLP(Microsatellite-anchored fragment length polymorphism)技术检测不同烟草材料遗传多样性的基础上,对不同烟草材料烟叶中TSNA含量的变异与标记多态性进行关联分析,以期发现与烟叶TSNA 含量显著关联的标记位点,为低TSNA含量的烟草品种的选育提供参考。
1 材料与方法
1.1 植物材料
供试的24个烟草材料由广东省烟草公司南雄科学研究所提供,其中包含了19个烤烟、3个晾晒烟和2个白肋烟,其来源和类型如表1所示。
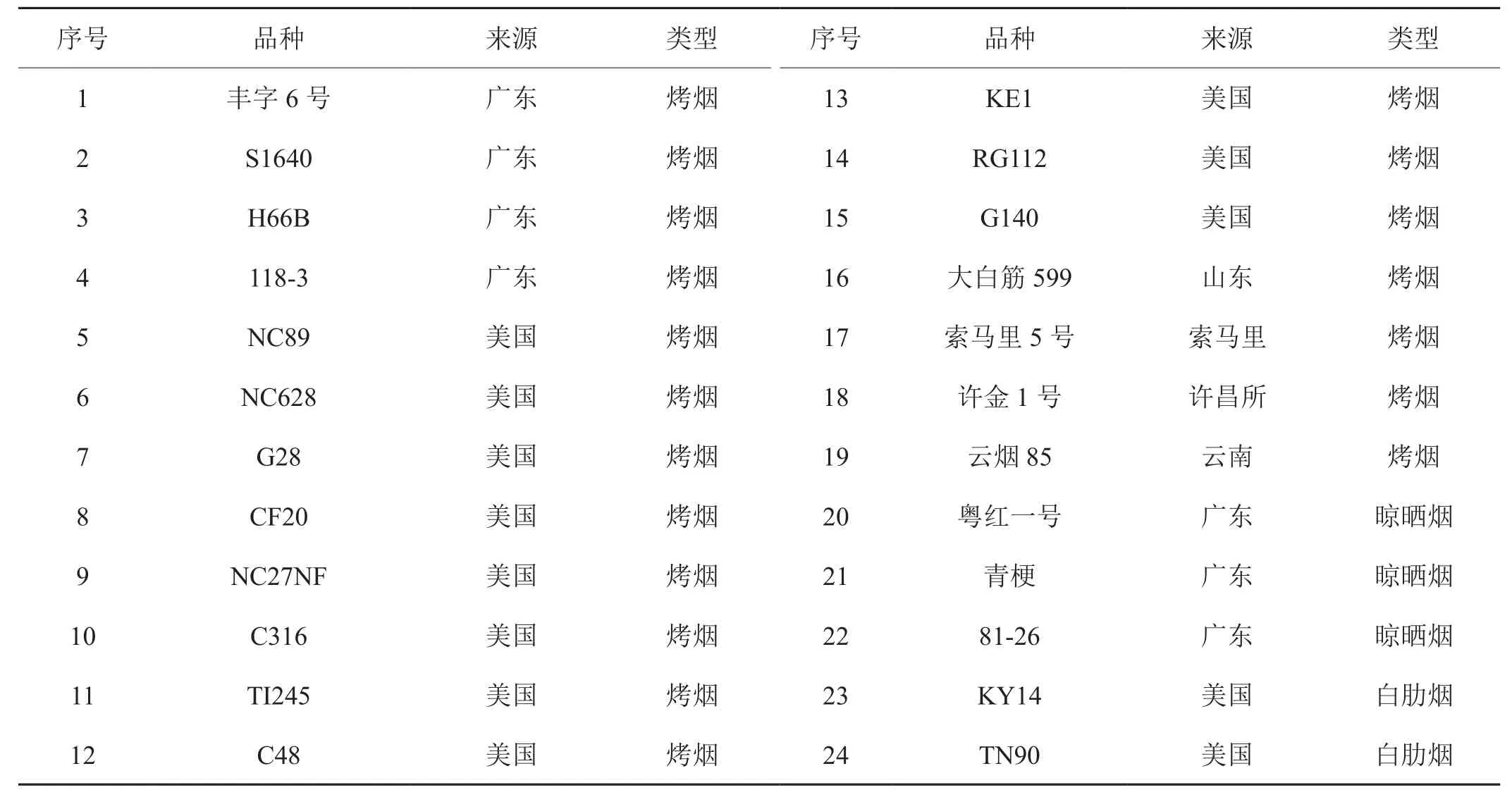
表1 烟草种质材料来源及类型Tab.1 Type and origin of tobacco accessions
1.2 烟叶TSNA含量的测定
烟草材料于2011年种植在广东省烟草公司南雄科学研究所的试验地,并设3个重复,按常规水肥管理。于成熟期采集不同品种相同节位的中部叶片进行杀青(105℃,30 min)和调制处理。参照丁时超等[19]方法测定调制前后烟叶中TSNA的含量。取适量烟叶除去主脉,在研钵中加入液氮迅速磨成粉末状,过40目筛,准确称取1.000 g待测烟粉用0.1%乙酸铵的水溶液(用双蒸水配制)超声萃取30 min,萃取液与等体积的乙腈(溶液中含有100 ng/mL内标)混合,高速离心分离后取上层清液,经0.22 μm水系微孔滤膜过滤,采用LC-MS/MS仪定量分析烟叶中TSNA含量。实验所需内标(NNN-d4和NNK-d4,纯度大于99%)和标样(NNN、NAT、NAB和NNK,纯度大于98%)均购自加拿大的TRC公司。
1.3 烟草材料遗传多样性的分析
采收烟草幼苗,按李荣华等[20]改进的CTAB方法提取烟草样品的DNA。根据Bindler等[21]的研究结果,选用分布于烟草24个连锁群的30个SSR标记(表2)来检测烟草材料的遗传多样性,PCR扩增体系参考何其芳等[22]所描述的方法进行;同时依据Yang等[23]MFLP技术的原理,结合荧光标记技术,检测烟草材料的遗传多样性,试验所使用的MFLP引物信息如表3所示。MFLP操作流程参考何其芳等[24]所描述的方法进行。PCR扩增产物在LICOR 4300 DNA遗传分析仪上进行变性聚丙烯酰胺凝胶电泳检测,使用GelBuddy 软件[25]对荧光扫描图谱中的扩增条带进行判读,根据条带的有无,排除模糊不清的条带和无法准确标识的条带,然后构建0,1二元数据矩阵,根据荧光标记DNA Marker 50-350 分子量标准,确定扩增条带的分子量来计算SSR标记和MFLP标记在烟草群体内的多态性。利用NTSYS-pc 2.11软件(Biostatistics Inc.,USA)计算了各烟草材料之间的遗传相似系数,然后采用非加权算术平均(Unweighted Pair-Group Mean Average,UPGMA)方法构建了烟草材料间的遗传聚类图。试验所需的荧光引物M13-F-IRDye 700(5′-CACGACGTTGTAAAACGAC-3′)购自美国LICOR 公司,其它引物、酶和试剂均购自上海生物工程有限公司。
1.4 关联分析
为了减少基因型与表型关联分析中存在的假阳性, 应用Structure 2.3软件(Pritchard et al.2000),对24个烟草材料进行基于数学模型的类群划分, 并计算材料相应的Q 值(第i材料其基因组变异源于第k 群体的概率)。然后将各个体Q值作为协变量,使用TASSEL软件的GLM(General Linear Model)程序[26],对烟叶的TSNA含量与标记多态性进行线性回归分析,阈值选择以P<0.05为显著性,P<0.01为极显著性。

表2 30个SSR标记的名称,等位位点数及多态信息量(PIC)Tab.2 Name, number of alleles and polymorphism information content (PIC) of 30 SSR markers

表3 MFLP 预扩增和选择性扩增的引物Tab.3 MFLP primers used in pre- and selective-amplification
2 结果与分析
2.1 不同烟草品种调制前后烟叶中TSNA含量的差异
经LC-MS/MS检测分析,24个烟草品种调制前后烟叶中TSNA含量的统计结果如表4所示。由表4可知,调制前烟叶中TSNA含量较低,TSNA总含量的平均值仅为32.85 ng/g,其中NNK在所有材料中都检测不到含量;调制后烟叶中TSNA显著增加,TSNA总含量的平均值增加到144.56 ng/g,其中NNN增加最显著,占TSNA总增加量的70%以上。不同烟草品种间TSNA含量的变异较大,调制前青烟叶中的NNN、NAT和NAB等3种成分的变异系数略低于调制后烟叶,而调制前后烟叶中TSNA总含量的变异系数基本一致,分别为1.47和1.49,这点说明烟叶中TSNA含量受品种基因型影响较大,且TSNA含量在群体中呈现连续分布,表明该性状受多基因控制。烟叶中TSNA各成分的相关性分析显示(表4),除调制前烟叶中NAB成分外,调制前后烟叶中其余各成分之间都存显著性正相关,且调制前烟叶中TSNA总含量与调制后烟叶中TSNA总含量也存显著性正相关,说明控制这4个TSNA含量的多基因中可能存在一些相同的微效基因。

表4 烟叶中TSNA含量及相关性分析Tab.4 TSNA content and its association analysis in tobacco leaves ng/g
2.2 烟草材料的遗传多样性及群体结构
试验所用的30个SSR 标记中,由于2个SSR标记没有扩增出特异性条带,仅用了28个SSR标记对24份烟草材料的遗传多样性进行了分析,共检测到127个等位变异,结果如表3所示;而MFLP多态性分析中, 5个加尾SSR锚定引物分别与15种MseI选择性扩增引物组合后筛选出33对适宜的引物组合,在24个烟草材料中共检测到61个多态位点。基于SSR标记和MFLP标记共同发现的188个多态位点,计算24个烟草材料间的遗传相似系数,再进行UPGMA聚类分析,结果如图1所示。由图1 可知,在相似系数0.65水平上,可将24个烟草品种分为3个分支:C1、C2和C3。第1分支(C1)仅包括两个烤烟品种,T1245 和 S1640; 第2分支(C2)包括2个晾晒烟(81-26和青梗)和1个烤烟品种(索马里5号),第3个分支(C3)包括1个晾晒烟(粤红一号)和16个烤烟品种和2个白肋烟。采用Structure 2.3软件分析参试种质的遗传结构,确定参试种质的亚群数目。结果表明24个烟草种质的等位变异频率特征类型数K=3(即服从Hardy-Weinberger平衡的亚群数目为3)时其模型后验概率最大,因此24个烟草材料分为3个亚群:G1,G2 和G3,分别包含了16、4和4个烟草材料(图2)。由图2可知,除了少数材料,Structure 2.3 软件的亚群分析与UPGMA聚类分析大体一致,其中G1的16个烟草材料都分布在C1中,G2包含了C2的3个烟草材料和C1中的1个晾晒烟,G3 包含了C3的2个烟草材料和C1中的2个白肋烟。进一步分析亚群的生物学意义,发现24个烟草亚群划分与不同烟草类型(烤烟、晾晒烟和白肋烟)基本一致。
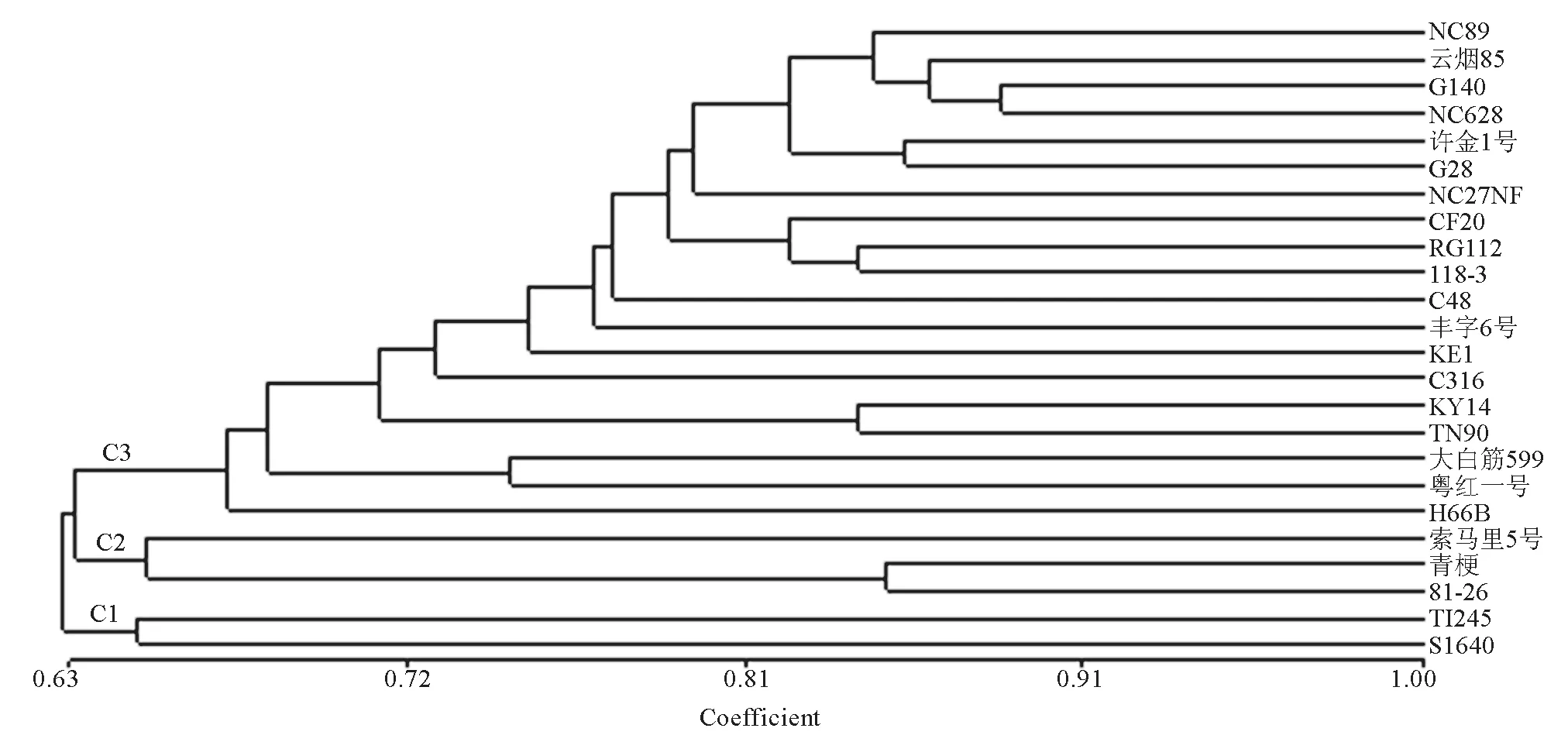
图1 24 个烟草品种基于遗传相似系数的UPGMA聚类分析Fig.1 UPGMA cluster analysis of 24 tobacco accessions based on genetic similarity coefficient
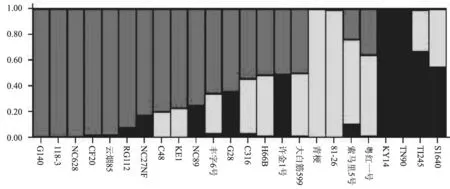
图2 24个烟草品种的群体结构分析Fig.2 Population structure of 24 tobacco accessions
2.3 烟叶TSNA含量与分子标记的关联分析
鉴于烟草材料由3个亚群体组成,将各个体相应的Q值作为协变量,利用TASSEL软件分析28个SSR标记和61个MFLP标记与烟叶中4种TSNA含量之间的关联性。关联分析结果显示,6个MFLP标记和1个SSR标记的变异至少与烟叶中1种TSNA含量在P<0.01水平上显著关联,其中1个MFLP标记(MFLP26)存在极显著相关性(P<0.001);7个关联标记对表型变异的解释率在28.78%~58.31%之间,平均值为36.02% (表5)。
由表5可知,SSR标记(PT30151)与调制后烟叶中NAB、NAT和NNK的含量都显著关联,其表型变异解释率分别为30.93%、40.20%和39.30%;同时,该SSR标记与调制前烟叶中NNN的含量也显著关联,其表型变异解释率41.17%。6个显著关联的MFLP标记中,MFLP55和MFLP5 分别与调制后烟叶中NAB和NNK的含量显著关联,其表型变异的解释率分别为28.78%和31.83%;MFLP26和MFLP1同时与调制后烟叶中NNN的含量显著关联,其表型变异的解释率分别为58.31%和34.12%;MFLP7同时与调制前烟叶中NAT和NNN显著关联,其表型变异的解释率分别为29.27%和31.00%;MFLP31与调制前烟叶中NNN显著关联,其表型变异的解释率为31.28%。

表5 与TSNA含量显著相关的标记位点及对表型变异的解释率Tab.5 Marker loci associated with TSNA content and their explained portion of phenotypic variation
3 讨论
关联分析 (association analysis),又称关联作图(association mapping),是一种以连锁不平衡为基础,以自然群体为研究对象,将目标性状表型的多样性与基因(或标记位点)的多态性结合起来分析,可直接鉴定出与表型变异密切相关且具有特定功能的基因位点或标记位点[27]。与传统的QTL作图相比,关联分析具有3个明显特点:(1)花费时间少,不需要专门构建作图群体,自然群体或种质资源都可作为研究材料;(2)广度大,广泛的遗传材料可同时考察多个性状的关联位点及其等位变异,不受传统QTL作图的“两亲本范围”的限制;(3)精度高,利用自然群体在长期进化中积累的重组信息,具有较高的解析率,可实现数量性状基因座的精细定位[28-29]。目前,关联分析已在拟南芥[30]、玉米[31]、水稻[32]、小麦[33]和高粱[34]等作物中广泛应用。
关联分析中最大的问题是假阳性关联的增大,这是由于材料的群体结构没有被计算而引起的等位变异评估的偏差产生[35]。本研究利用分布烟草24个染色体的28个SSR标记和MFLP标记技术分析烟草材料遗传多样性的基础上,对材料的群体结构进行了分析,24个烟草材料分为3个亚群(G1-G3),与UPGMA聚类分析分布的3个分支(C1-C3)比较,除了少数材料,亚群分布与遗传聚类分支基本对应(G1/C1,G2/C2和G3/C3),且亚群划分与烟草类型(烤烟、晾晒烟和白肋烟)基本一致。早期的一些研究也显示,我国烟草材料不同烟草类型间遗传差异较大,而种内遗传多样性水平较低,且不同地理来源品种间的遗传差异不明显[36-37,22]。因此,群体结构分析比UPGMA聚类分析能更好地从分子水平上揭示烟草种质资源的遗传背景和亲缘关系。
利用了群体结构分析中材料的Q值作为协变量来关联分析多态位点与TSNA含量间的关系,结果显示烟草群体中1个SSR位点和6个MFLP位点至少与1种TSNA的相关性在P<0.01水平上显著,其中SSR位点与4种TSNA显著关联,该位点可能共同作用于这4种TSNA的形成;而4个MFLP位点都与NNN显著关联,则表明NNN可能由多基因控制,其中标记MFLP26可能与其主效基因相关联。刘万峰[13]对不同组合的亲本和F1中的TSNA研究也推断,TSNA含量是一种由寡基因控制的不完全显性遗传或者是一种由微效多基因控制的数量性状。多数研究证实[8,38-41],TSNA在鲜烟叶中积累很少或几乎不产生,其形成与积累主要是在采收后产生的,且大部分是产生于调制期间。在调制过程中,烟草中的硝酸盐被微生物还原为亚硝酸盐以及氮氧化物(NOx),然后与烟草生物碱作用形成TSNA[42],且调制后烟叶中TSNA含量与烟叶中的烟碱及亚硝酸盐的含量呈正相关性[10,41,44]。史宏志等[18]研究显示,烟碱转化在遗传上受显性基因控制,通过遗传改良降低烟碱转化率和降烟碱含量可有效降低烟叶NNN和总TSNA的含量。Julio等[45]利用114个烤烟重组自交系发现了控制降烟碱和假木贼烟碱含量的4个QTL位点,其对变型变异的解释率在9.0%~27.8%,这些QTL位点也间接地影响烟叶TSNA的含量。本次试验结果显示,调制后的烟叶总TSNA的含量比调制前烟叶增加了3.4倍之多,但调制前后烟叶中总TSNA含量间的相关系数仅为0.724,且调制前后显著关联的分子标记也不尽相同,这可能的原因是不同品种间烟叶中烟碱和亚硝酸盐含量存在一定差异,或调制方式及微生物含量的变动,致使调制后品种间烟叶中TSNA含量有较大变化。因此,本实验显著关联的分子标记需要在杂交群体中进一步验证后才能应用于分子辅助育种。
4 结论
利用分布于全基因组的28个SSR标记和MFLP标记技术,分析了24个烟草材料的遗传特性,群体结构分析发现,24个烟草材料分为3个亚群,且亚群划分与不同基因类型(烤烟、晾晒烟和白肋烟)相关。对烟叶中TSNA含量的变异与标记的多态性关联分析显示,6个MFLP标记和1个SSR标记的变异至少与烟叶中1种TSNA含量在P<0.01水平上显著性关联,其中1个MFLP标记(MFLP26)存在极显著性相关联(P<0.001)。这些关联标记可能为进一步的培育出低TSNA的高安全性烟草品种提供参考,也为采用分子生物学手段或常规育种手段来进行遗传改良降低烟草中的TSNA含量提供了可靠的方法与途径。
[1]Hecht S S, Chen C H B, Young R, et al.Mass spectra of tobacco alkaloid-derived nitrosamines, their metabolites,and related compounds[J].Beiträge zur Tabakforschung International, 1981, 11(2): 57-66.
[2]Brunnemann K D, Hoffmann D.Analytical studies on tobaccospecific N-nitrosamines in tobacco and tobacco Smoke[J].Critical Reviews in Toxicology, 1991, 21(4): 235-240.
[3]Hecht S S, Hoffmann D.The relevance of tobacco-specific nitrosamines to human cancer[J].Cancer Surveys, 1989, 8(2):273-294.
[4]Spiegelhalder B, Bartsch H.Tobacco-specific nitrosamines[J].European Journal of Cancer Prevention, 1996, 5 (S1): 33-38.
[5]Hecht S S.Biochemistry, biology, and carcinogenicity of tobacco-specific N-nitrosamines[J].Chemical Research in Toxicology, 1998, 11(6): 559-603.
[6]Wu J C, Joza P, Sharifi M, et al.Quantitative method for the analysis of tobacco-specific nitrosamines in cigarette tobacco and mainstream cigarette smoke by use of isotope dilution liquid chromatography tandem mass spectrometry[J].Analytical Chemistry, 2008, 80(4):1341-1345.
[7]李勇.卷烟主流烟气中几类有害化合物的分析技术的研究与应用[D].湘潭: 湘潭大学, 2007.
[8]Bhide S V, Nair J, Maru G B, et al.Tobacco-specific N-nitrosamines (TSNA) in green mature and processed tobacco leaves from India[J].Beitrage zur Tabakforschung International, 1987, 14(1): 107-112.
[9]史宏志, Bush L P, 黄元炯, 等.我国烟草及其制品中烟草特有亚硝胺含量及与前体物之间的关系[J].中国烟草学报, 2002, 8(1): 14-19.
[10]Chamberlain W J, Chortyk O T.Effects of curing and fertilization on nitrosamine formation in bright and burley tobacco[J].Beiträge zur Tabakforschung international, 1992,2(15): 87-91.
[11]Burton H R, Dye N K, Bush L P.Relationship between tobacco specific N-nitrosamines and nitrite from different air-cured tobacco varieties[J].Journal of agricultural and food chemistry,1994, (42): 2007-2011.
[12]余义文, 夏岩石, 李荣华, 等.不同类型及品种烟草特有亚硝胺含量的分析[J].烟草科技, 2013(4): 46-51.
[13]刘万峰.不同基因型烤烟烟叶中TSNA及其前体物质的研究[D].北京: 中国农业科学学院, 2002.
[14]Lewis R S, Parker R G, David A, et al.Impact of alleles at the yellow burley (Yb) loci and nitrogen fertilization rate on nitrogen utilization efficiency and tobacco-specific nitrosamine(TSNA) formation in air-cured tobacco[J].Journal of agricultural and food chemistry, 2012, 60: 6454-6461.
[15]Guo Peiguo, Xia Yanshi, Li Ronghua, et al.Assessment of tobacco specific nitrosamines in tobacco genotypes[J].Advanced Materials Research, 2013, 641(642):871-874.
[16]张俊杰, 林国平, 王毅, 等.白肋烟低TSNA含量的品种筛选初探[J].中国烟草学报, 2009, 15(3): 54-57.
[17]史宏志, 刘国顺.白肋烟烟碱转化及烟草特有亚硝胺形成[J].中国烟草学报, 2008, 14(增刊): 41-46
[18]史宏志, 李进平, 李宗平, 等.遗传改良降低白肋烟杂交种烟碱转化率研究[J].中国农业科学, 2007, 40(1): 153-160.
[19]丁时超, 杜文, 任建新, 等.LC-MS /MS 定量分析卷烟中的烟草特有亚硝胺(TSNA) [J].中国烟草学报, 2005,11(6):17-21.
[20]李荣华, 夏岩石, 刘顺枝, 等.改进的CTAB提取植物DNA方法[J].实验室研究与探索, 2009, 28(9): 14-16.
[21]Bindler G, Hoeven R, Gunduz I, et al.A microsatellite marker based linkage map of tobacco[J].Theor Appl Genet, 2007,114: 341-349.
[22]何其芳, 李荣华, 郭培国, 等.利用SSR荧光标记技术分析烟草种质的遗传多样性[J].中国农学通报, 2012a,28(10): 95-102.
[23]Yang H, Sweetingham M W, Cowling W A, et al.DNA fingerprinting based on microsatellite-anchored fragment length polymorphisms, and isolation of sequence-specific PCR markers in lupin (Lupinus angustifoliusL.)[J].Molecular Breeding, 2001(7): 203-209.
[24]何其芳, 李荣华, 郭培国, 等.利用荧光MFLP 标记技术分析烟草种质的遗传多样性[J].中国烟草科学, 2012b,33(1): 12-18.
[25]Zerr T, Henikoff S.Automated band mapping in electrophoretic gel image using background information[J].Nucleic Acids Research, 2005, 33: 2806-2812.
[26]Bradbury P J, Zhang Z, Kroon D E, et al.TASSEL: software for association mapping of complex traits in diverse samples[J].Bioinformatics, 2007, 23(19): 2633-2638.
[27]March R E.Gene mapping by linkage and association analysis[J].Mol Biotechnol, 1999, 13: 113-122.
[28]Yu J, Edward S B.Genetic association mapping and genome organization of Maize[J].Current Opinion in Biotechnology,2006, 17: 155-160.
[29]Mackay I, Powell W.Methods for linkage disequilibrium mapping in crops[J].Trends Plant Science, 2007, 12: 57-63.
[30]Ehrenreich I M, Hanzawa Y, Chou L, et al.Candidate gene association mapping of Arabidopsis flowering time[J].Genetics, 2009, 183: 325-335.
[31]Harjes C E, Rocheford T R, Bai L, et al.Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification[J].Science, 2008, 319: 330-333.
[32]Iwata H, Ebana K, Uga Y, et al.Genome-wide association study of grain shape variation amongOryza sativaL.germplasms based on elliptic Fourier analysis[J].Molecular Breeding, 2010, 25: 203-215.
[33]Breseghello F, Sorrells M E.Association mapping of kernel size and milling quality in wheat (Triticum aestivumL.)cultivars[J].Genetics, 2006, 172: 1165-1177.
[34]Casa A M, Pressoira G, Brown P J, et al.Community resources and strategies for association mapping in sorghum[J].Crop Sciences, 2008, 48: 30-40.
[35]Ahmadi N, Negrão S, Katsantonis D, et al.Targeted association analysis identifiedjaponicarice varieties achieving Na+/K+homeostasis without the allelic make-up of the salt tolerantindicavariety Nona Bokra[J].Theoretical and Applied Genetics, 2011, 123: 881-895.
[36]杨友才, 周清明, 尹晗琪.利用RAPD和AFLP标记分析烟草种质资源的遗传多样性[J].农业生物技术学报,2006, 14(4): 585-593.
[37]李凤霞, 王卫锋, 王鲁, 等.烟草属植物遗传多样性和亲缘进化关系的荧光AFLP分析[J].中国农业科学, 2010,43(12): 2418-2427.
[38]杨焕文,李永忠,刘彦中,等.白肋烟晾制中亚硝胺及前体物质的变化[J].西南农业大学学报,1999, 21(5): 444-448.
[39]刘国顺, 赵春华, 崔树毅, 等.烟草特有亚硝胺及其前体物的研究进展[J].中国烟草学报, 2007, 13(1): 44-47.
[40]雷丽萍,Luo L, Shi H, 等.烟叶调制过程中相关物质的测定[J].中国烟草科学,2002, 23(4): 32-34.
[41]Djordjevic M V, Gay S L, Bush L P, et al.Tobacco-specific nitrosamine accumulation and distribution in flue-cured tobacco alkaloid isolines[J].J Agr Food Chem, 1989, 37(3):752-756.
[42]宫长荣,沈剑波,司辉,等.烟草N-TSNAs 及其前体物在烟叶烘烤过程中含量的变化[J].中国农学通报,2007,23(6): 179-182.
[43]Burton H R, Childs G H, Anderson R A, et al.Changes in chemical composition of burley tobacco during senescence and curing.3.tobacco-specific nitrosamines[J].J Agr Food Chem,1989, 37(2): 426-430.
[44]Mackown C T, Eivagi F, Sims L.Tobacco specific N-nitrosamines: effects of burley isolines and nitrogen fertility management[J].J Agr Food Chem, 1984, 32(6): 1269-1272.
[45]Julio E, Denoyes-Rothan B, Verrier J L, et al.Detection of QTLs linked to leaf and smoke properties inNicotiana tabacumbased on a study of 114 recombinant inbred lines [J].Mol Breed, 2006, 18: 69-91.
Association analysis of tobacco specific nitrosamines content in tobacco germplasm
YU Yiwen1, XIA Yanshi1, LI Ronghua1, Lü Yonghua2, GUO Peiguo1, QIU Miaowen3, ZHAO Weicai3, HE Qifang2
1 College of Life Sciences, Guangzhou University, Guangzhou 510006, China;
2 Guangdong Provincial Tobacco Monopoly Administration, Guangzhou 510610, China;
3 Nanxiong Research Institute of Guangdong Tobacco Co.Ltd., Nanxiong, Guangdong 512400, China
Genetic diversity analysis for 24 tobacco accessions was performed with SSR and MFLP markers which distributed in 24 chromosomes.Association analysis was conducted between polymorphic markers and tobacco specific nitrosamine (TSNA) contents through GLM model using software TASSEL.Result showed that 188 allelic variations were detected among 24 tobacco accessions with 33 MFLP and 28 SSR primer combinations.These accessions were clustered into 3 subgroups in genetic analysis of population structure.The subgroup is in coincidence with tobacco types, i.e.flue-cured, sun-cured and burley.Association analysis showed that 6 MFLP markers and 1 SSR marker were associated with at least one of TSNA contents at P<0.01 level.The marker of MFLP26 was significantly associated with NNN content (P<0.001), and explains 58.31% of phenotypic variation for NNN content in cured leaves.These markers could be used in tobacco breeding programmes to assist with selection of tobacco genotypes with low TSNA content.
tobacco specific-nitrosamine; SSR marker; MFLP marker; association analysis
国家烟草专卖局面上项目“烟草特有亚硝胺含量的关联分析及遗传筛选”(2010[99]18);广东省烟草专卖局科技计划项目“烟草特有亚硝胺含量的关联分析及遗传筛选”(200903)
余义文(1987—),硕士研究生,主要从事烟草分子生物学研究,Tel: 020-39366915, Email: 358614803@qq.com
郭培国(1963—),博士,教授,主要从事作物功能基因组和分子遗传育种的教学与研究,Tel: 020-39366915,Email: guopg@yahoo.com
2013-02-12
10.3969/j.issn.1004-5708.2014.03.008
S572.03; Q78 文献标志码:A 文章编号:1004-5708(2014)03-0048-08
