弓形虫SAG2和GRA2基因疫苗的免疫保护研究*
2014-11-10全娟花喻才元楚佳奇
全娟花 李 鹏 喻才元 楚佳奇
(1.广东医学院附属医院干细胞研发与细胞治疗中心,广东湛江 524001;2.广东医学院附属医院消化内科,广东湛江 524001)
刚地弓形虫Toxoplasmagondii是一种专性细胞内寄生性原虫,可引起人兽共患的弓形虫病,尤其对孕妇、免疫抑制和免疫缺陷患者的影响更为严重(Pappasetal. , 2009;Sakikawaetal. , 2012)。近年来,弓形虫疫苗经历了全虫疫苗、虫体特异组分疫苗和核酸疫苗等研究阶段。结果发现,全虫疫苗和虫体特异组分疫苗能够预防弓形虫感染或降低感染程度,但前者易引起再感染,后者的免疫保护作用较差(Angusetal., 2000;Innesetal., 2006)。DNA疫苗是将编码抗原的基因插入载体质粒中构成重组体,直接接种机体,在接种部位摄取表达并达到抗感染目的的一种疫苗。它的优点是内源性表达的抗原以天然构型出现,诱导全身性免疫应答。DNA疫苗相较于传统的灭活及减毒疫苗更加高效并克服了回复突变的潜在危险,相较于基因工程多肽疫苗降低了成本和简化了制备过程(Ismaeletal., 2003;Fachadoetal., 2003)。
近年研究表明,含有人巨细胞病毒(Human cytomegalovirus,CMV)高效启动子/增强子序列的真核表达载体在哺乳动物细胞内高效表达成为DNA疫苗载体。当前,有不少学者对弓形虫表面膜抗原(Surface antigen,SAG)基因、棒状体蛋白(Rhoptry protein,ROP)基因、致密颗粒(Dense granule protein,GRA)基因、微线体蛋白(Microneme protein,MIC)等进行核酸疫苗研究(李运娜等, 2011)。SAG2是由P22基因编码的天然蛋白质,只在弓形虫的侵入期速殖子表达。抗SAG2抗体能阻止虫体在宿主细胞膜的固定附着,从而阻止其入侵宿主细胞,并促使包囊的形成增加,SAG2在弓形虫感染的免疫中发挥一定的作用(Rachineletal., 2004)。高世同等(2004)针对弓形虫SAG2编码基因构建了真核表达质粒pVAX1-SAG2,并接种小鼠发现,免疫组小鼠血清中出现抗弓形虫特异性IgG抗体并且能够诱导小鼠产生较强的体液免疫和细胞免疫应答。致密颗粒蛋白(GRA)是弓形虫致密颗粒细胞器分泌的一类具有免疫活性的蛋白质。它们在修饰调理纳虫泡中起重要作用,并关联于泡内网络结构,参与虫体在细胞内的存活和复制(Desolmeetal. , 2000)。有研究报道,用pVAC-GRA4真核表达重组质粒免疫小鼠,观察诱导的免疫应答及其对弓形虫感染的保护作用,结果显示pVAC-GRA4免疫组小鼠存活率显著高于对照组,诱导产生以细胞免疫为主的免疫应答(林绮萍等, 2005)。
DNA疫苗在抗弓形虫病方面具有良好的开发应用前景,国内外以开展较多,但有关弓形虫pSAG2-GRA2双基因疫苗的系统研究尚未见报道。本研究选用SAG2和GRA2基因作为重组子,以pCMV-Taq2B为载体,利用基因重组技术,成功制备了pSAG2、pGRA2及pSAG2-GRA2。DNA疫苗直接肌注免疫小鼠,观察其在宿主细胞内不断表达及在体内诱导的免疫应答反应,并通过动物攻击实验,评价其免疫保护效果,为构建安全有效的弓形虫DNA疫苗的研制奠定基础。
1 材料与方法
1.1 材料
1.1.1 试剂:质粒提取试剂盒和胶回收试剂盒为Qiagen公司产品;核酸内切酶BamHI、PstI、EcoRI、EcoRV、T4连接酶均为Roche公司产品;胎牛血清、RPMI1640、表皮生长因子(EGF)、Opti-MEM和Lipofectamine LTX&PLUS均为Gibco公司产品;GelRed是Biotium公司产品;羊抗小鼠IgG购自美国Sigma公司。
1.1.2 质粒pCMV-Taq2B、FHs 74 Int细胞株、大肠杆菌DH5α为本室保存。
1.1.3 弓形虫RH株由韩国忠南大学医学院感染生物学教研室教授Young-Ha Lee馈赠。BALB/c及ICR小鼠,6周龄左右,购自广东省医学实验动物中心。
1.2 方法
1.2.1 真核表达载体的构建:按照所选取的目的片段由武汉博洪生物科技有限公司设计以下两对引物:SAG2,C G C G G A T C C T C A A A G A C C A C G A G C C T A GC和A A T C T G C A G C T T G C C C G T G A G A G A C A C AG,下划线部分分别为BamHI、PstI酶切位点;GRA2,C C G G A A T T C A T G T T C G C C G T A A A A C A T TG和A A T G A T A T C T T G G G G C T C C A C T G G A A C CT,下划线部分分别为EcoRI、EcoRV酶切位点。以弓形虫RH株速殖子cDNA为模板扩增SAG2和GRA2目的片段。按常规方法将该目的基因分别插入pCMV-Taq2B真核表达质粒BamHI、PstI和EcoRI、EcoRV酶切位点之间,转化大肠杆菌DH5α,重组质粒经DNA测序鉴定正确后命名为pSAG2和pGRA2。SAG2目的基因插入pGRA2质粒BamHI、PstI酶切位点之间,该重组质粒命名为pSAG2-GRA2。
1.2.2 质粒DNA的提取及纯化:碱裂解法分别提取重组质粒pSAG2、pGRA2、pSAG2-GRA2和空质粒pCMV-Taq2B,溶于灭菌PBS。定量使得浓度均为1 mg/mL。
1.2.3 细胞转染:将FHs 74 Int人小肠上皮细胞以1×105每孔接种于6孔板中,加入2 mL培养基(含有10% FBS和20 ng/mL EGF的RPMI1640),置于37 ℃,5% CO2培养箱中培养,细胞密度达80%时参照Lipofectamine LTX&PLUS转染试剂说明进行细胞转染。
1.2.4 逆转录聚合酶链反应(RT-PCR)检测SAG2和GRA2 mRNA表达:按Trizol试剂说明提取转染FHs 74 Int细胞总RNA,按M-MLV cDNA合成试剂盒说明合成cDNA第一链并用上述的SAG2和GRA2引物按照说明进行PCR反应。以GAPDH作为内参照。
1.2.5 DNA疫苗的免疫接种:取SPF级雌性8周龄BALB/c小鼠115只,随机分为5组,即注射PBS、pCMV-Taq2B空质粒对照组和注射重组质粒pSAG2、pGRA2、pSAG2-GRA2实验组,每组23只。小鼠双侧胫前肌相同部位分别注射50 μL(50 μg)质粒,每只小鼠每次免疫的质粒总量为100 μg。于初次免疫后第14、28 d分别加强免疫。
1.2.6 弓形虫速殖子培养裂解物(TLA)的制备:将复苏后的弓形虫RH强毒株腹腔注射ICR小鼠,待小鼠出现症状(如竖毛、不食、肛门拖粪、呼吸困难等)后,用生理盐水冲洗腹腔,抽取腹腔液再腹腔注射给其他ICR小鼠传代,3 d传代1次。收集腹腔中的速殖子,洗涤后悬浮于PBS,反复冻融10次。离心后取上清经0.22 μm无菌滤器过滤后,即为 TLA。
1.2.7 ELISA 法测定小鼠血清IgG:分别于免疫前及末次免疫后第14、28 d,小鼠断尾法取血,分离收集小鼠血清。将每次所取血清样品用间接ELISA法检测免疫小鼠IgG抗体,具体方法参见文献(Quanetal. , 2012)。
1.2.8 免疫小鼠抗弓形虫攻击感染实验:每组小鼠(n=20)均注射经ICR小鼠腹腔传代的弓形虫RH株速殖子,注射剂量1 000个/鼠,监测小鼠存活时间。
1.2.9 统计方法 实验数据分析采用单因素方差分析检验(One-way ANOVA),P< 0.05为有统计学意义。
2 结果
2.1 重组质粒在小肠上皮细胞中表达
以弓形虫RH株cDNA为模板,扩增出SAG2和GRA2片段,PCR产物大小与预期值相符(图1)。对构建的3种重组质粒进行DNA测序,结果与GenBank收录的SAG2和GRA2序列完全一致,基因读码框架正确。为了鉴定pSAG2、pGRA2及pSAG2-GRA2 DNA疫苗能否成功表达具有免疫原性的SAG2和GRA2蛋白,本实验以pCMV-Taq2B空载体为对照转染FHs 74 Int细胞,用RT-PCR法在转录水平鉴定SAG2和GRA2蛋白表达。结果显示,在经pSAG2、pGRA2或pSAG2-GRA2转染的小肠上皮细胞总RNA中扩增出一条DNA片段,该片段大小与SAG2、GRA2的PCR扩增产物一致(图2)。而在未转染的阴性对照小肠上皮细胞和空载体转染细胞中虽能扩增出GAPDH mRNA,但未检测到SAG2或GRA2 mRNA,从而证实SAG2和GRA2在转染细胞中有效表达。

图1 弓形虫表面膜抗原2(SAG2)和致密颗粒蛋白2(GRA2)PCR产物Fig.1 PCR products of Toxoplasma gondii Surface antigen 2(SAG2)and Dense granule protein 2(GRA2)
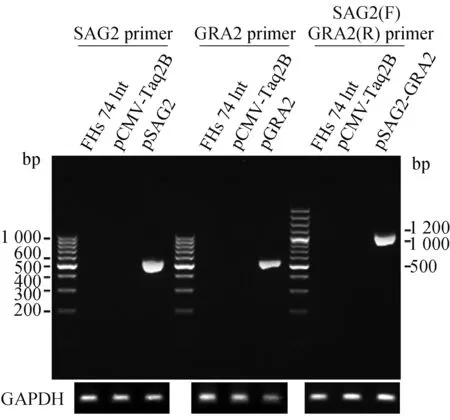
图2 RT-PCR 检测FHs 74 Int转染细胞中弓形虫表面膜抗原2(SAG2)和致密颗粒蛋白2(GRA2)基因mRNA的表达结果Fig.2 Expression of Toxoplasma gondii Surface antigen 2(SAG2)and dense granule protein 2(GRA2)mRNA in FHs 74 Int transfected cells by RT-PCR
2.2 免疫小鼠弓形虫特异性抗体应答
ELISA结果(图3)表明,pSAG2、pGRA2和pSAG2-GRA2基因疫苗免疫组小鼠均能诱导产生特异性IgG抗体,均于末次免疫后第28 d达到最高值,此时pSAG2(0.265±0.087)、pGRA2(0.382±0.66)和pSAG2-GRA2(0.548±0.137)免疫组吸光度值(OD490 nm)显著高于免疫前吸光度值(0.139±0.017、0.136±0.007、0.134±0.009)(P<0.05),表明基因疫苗免疫小鼠产生的IgG抗体为阳性。基因疫苗组血清抗体的吸光度值均高于空载体和PBS阴性对照组,尤其是pGRA2(0.382±0.66)和pSAG2-GRA2(0.548±0.137)组吸光度值显著高于PBS(0.137±0.016)或pCMV-Taq2B(0.135±0.010)对照组吸光度值(P<0.05)。

图3 ELISA法检测免疫小鼠血清中IgG抗体Fig.3 Detection of IgG in serum of immunized mice by ELISA
2.3 抗攻击保护作用
为了评价DNA疫苗对急性弓形虫感染的保护作用,末次免疫后第28 d每只小鼠经腹腔注射1 000个速殖子后每天观察生存时间(d)直到所有攻击感染的小鼠死亡。 攻击小鼠2~3 d后,免疫基因疫苗pSAG2、pGRA2及pSAG2-GRA2的实验组小鼠均出现竖毛、倦怠、活动及饮食减少等表现。实验小鼠均未能耐受攻击弓形虫RH株速殖子攻击感染。小鼠攻虫感染后的存活率曲线见图4,免疫单基因疫苗pSAG2(14.3±1.8)、pGRA2组(13.5±2.1)小鼠存活时间明显长于PBS(4.3±1.6)及pCMV-Taq2B对照组(4.1±1.3)(P<0.05),且双基因疫苗pSAG2-GRA2组(19.6±2.4)小鼠存活时间明显长于单基因疫苗pSAG2组及pGRA2组(P<0.05),证明真核表达质粒pSAG2、pGRA2、pSAG2-GRA2作为DNA疫苗进行免疫能诱导机体产生一定的针对弓形虫感染的保护性免疫应答,使实验小鼠的存活时间明显延长,且双基因疫苗优于单基因疫苗。

图4 腹腔注射弓形虫速殖子后免疫小鼠生存率Fig.4 Survival rate of mice in different groups after oral challenged with T. gondii tachyzoite
3 讨论
DNA疫苗可通过多种途径进行,溶于PBS后,可直接肌肉、皮内、皮下或静脉注射,也可将其沉淀到金颗粒上,通过基因枪导入细胞内。其中直接肌注法是目前比较认可的有效途径之一。本研究构建了真核表达重组质粒pSAG2、pGRA2和pSAG2-GRA2并应用胫前肌直接肌注法疫苗免疫小鼠,引发了免疫保护反应。
pSAG2、pGRA2和pSAG2-GR2基因疫苗实验组血清IgG抗体浓度均随免疫接种次数的增多而显著增高,尤其在末次免疫接种28 d后血清抗体浓度达到最高水平,两对照组血清中抗体浓度无变化。实验组血清抗体浓度均高于空载体和PBS阴性对照组,尤其pSAG2-GRA2组显著高于对照组(P<0.05)。攻击感染实验结果显示基因疫苗实验组感染小鼠的存活时间明显延长,但最终均未能存活,其主要原因可能是本实验中弓形虫速殖子毒力完好且攻击量(1 000/鼠)较高,疫苗诱导的机体免疫应答不能完全清除感染的速殖子。弓形虫自然感染中间宿主的主要途径是卵囊经口感染,若攻击感染改用经口途径模拟自然感染,或用弱毒株进行攻击感染,则更能较客观地评价其免疫保护效果。Wu等(2012)研究显示,用真核表达载体pVAX1-GRA1和pVAX1-SAG1对实验小鼠作直接的肌肉注射,可诱发机体产生抗攻击保护,与本实验结果基本吻合。
本研究通过对免疫小鼠特异性抗体测定和抗弓形虫攻击感染实验,证明抗弓形虫单基因疫苗pSAG2、pGRA2和双基因疫苗pSAG2-GRA2能诱导机体产生部分抗弓形虫保护性免疫应答。尤其pSAG2-GRA2双基因疫苗免疫小鼠所诱导的免疫保护作用,可显著降低弓形虫感染所致的死亡率。本研究为构建安全有效的弓形虫DNA疫苗提供理论和实践基础。
