纤毛虫伪红色双轴虫营养细胞和休眠包囊的超微结构观察
2014-09-22运迷霞顾福康范鑫鹏
孙 磊,运迷霞,顾福康,范鑫鹏
(华东师范大学生命科学学院,上海200062)
原生动物纤毛虫在不利生长条件下经常会形成包囊,而一旦环境条件变为有利时其形成包囊的细胞又会脱囊而出,进行正常的生命活动.纤毛虫形成包囊和脱包囊期间,细胞结构也相伴发生去分化和再分化的过程,这可能是纤毛虫细胞在特殊生理条件下进行结构重组的一种生存策略[1].研究纤毛虫的包囊现象,对于揭示细胞结构的形成和调控具有重要学术意义[2-3].目前,对纤毛虫形成包囊和脱包囊过程中纤毛结构的分化已有较为深入的了解,根据纤毛虫形成包囊后纤毛结构的分化特征,提出了纤毛虫形成毛基体吸收型包囊、毛基体非吸收型包囊和尾柱虫类包囊三种类型包囊[4-5],但对不同类群的纤毛虫形成包囊后细胞内细胞器的结构分化及其生命活动特征尚知之不多.作者通过对腹毛类纤毛虫伪红色双轴虫(Diaxonella pseudorubra)营养期细胞和包囊细胞的超微结构观察,探索了不同生理条件下细胞核器、线粒体、黏液泡等结构的分化,获得了若干新的结果,报道如下.
1 材料与方法
1.1 材料
伪红色双轴虫采自山东省济南市趵突泉.在室温下,应用池塘过滤水加麦粒制备细菌发酵培养液,接种纤毛虫进行纯系培养.将纤毛虫培养至较高密度后,不更换培养液,致使培养液“老化”,诱导纤毛虫形成包囊.以非分裂状态的营养期纤毛虫和包囊时期的纤毛虫为材料制备扫描电镜(Scanning Electron Microscopy,SEM)和透射电镜(Transmission Electron Microscopy,TEM)样品.
1.2 扫描电镜样品制备和观察
扫描电镜制样方法参考顾福康[6]等的方法作少许调整,收集生长状态较好的营养期细胞和不同时期的包囊细胞,用池塘过滤水清洗3~4次后固定,固定液由1%的锇酸和饱和升汞溶液按1∶6(体积比)混合而成.固定5 min后,用0.1 mol/L PBS清洗3次,5 min/次.此后以梯度乙醇脱水,转入醋酸异戊酯,经临界点干燥,装台喷金制得样品,在JSM-5610扫描电子显微镜下观察、照相.
1.3 透射电镜样品制备和观察
透射电镜制样方法参考顾福康[7]的方法作少许调整,收集生长状态良好的纤毛虫营养期细胞和形成包囊的细胞,用池塘过滤水清洗3~4次后固定.其中前固定液由5%的戊二醛和2%的锇酸等体积混合而成,于4℃下固定10 min,0.1 mol/L PBS清洗3次,5 min/次,然后用1%锇酸后固定1 h,梯度丙酮脱水,Epon812树脂包埋,LKB超薄切片机切片,醋酸双氧铀和柠檬酸铅染色,JEM2100透射电镜观察和照相.
2 结果
2.1 营养期纤毛虫的超微结构
2.1.1 扫描电镜术显示的表面结构
伪红色双轴虫细胞长约145μm、宽约35μm,胞体呈长棒状,前部钝圆,尾部略尖.皮层腹面纤毛器中,口围带约由34片小膜组成,波动膜2片;额区棘毛包括额棘毛和口棘毛两部分,其中额棘毛3根,拟口棘毛约4根,口棘毛5根;中腹棘毛列约由15对棘毛组成,延伸至虫体3/5处;横棘毛约8根;左缘棘毛2或3列,自右至左每列的棘毛数依次减少,右缘棘毛一列,纵贯细胞全长(图1:1).背皮层纤毛器中,背触毛3或4列(图1:2).
2.1.2 透射电镜观察
伪红色双轴虫细胞外围是由质膜和表膜泡组成的表膜结构,表膜下微管按规则排列形成由单层微管加固的表质层(图2:1).在经过口围带的切片中,各片小膜按一定间隔排列,各含3列纤毛或纤毛基体(图2:2).
大核由两层核膜包围,在核膜位置偶见核孔,此外也有不少膜样结构或膜泡与核膜外膜相联系;大核染色质聚集成大小不一的团块状,非均一地分布在核内,也有些染色质附着在核膜内膜;核仁较小,着色较深,分布在核内一侧(图2:3).小核由两层核膜包围,核内染色质成致密均一分布,染色质颗粒大小相似,但很少附着于核膜上(图2:4).
在表膜下细胞质至细胞深部含有线粒体、食物泡、黏液泡和色素颗粒等细胞器.其中,线粒体由双层膜包围,内膜折叠形成管状嵴(图2:5),在皮层纤毛器、细胞核及食物泡附近大量分布(图2:2,3,6,7);食物泡呈不规则的球形,其外围单层膜,泡内充满处于消化中的膜泡样和颗粒状的物质(图2:8);黏液泡在营养期细胞中大量存在(图2:9),其由单层膜包围,内含颗粒状晶体(图2:10);色素颗粒主要包括直径约0.2μm和直径约0.5μm的两种不同大小,也偶见直径大于0.5μm的颗粒(图2:12).此外,在细胞质内见到少数电子密度较高的直径和长度约为0.25μm和0.5μm的梭形体,其被包围在粘滞状的细胞质内,可能是一类共生性的细菌(图2:11).
2.2 包囊期纤毛虫的超微结构
2.2.1 纤毛虫形成包囊的形态学
伪红色双轴虫形成包囊时,细胞逐渐收缩,胞体经历了从长棒形至椭圆形、直至球形的变化,腹面纤毛器中口围带和波动膜、额腹横腹棘毛和左右缘棘毛汇聚在一起,并伴随着退化、瓦解和消失的过程,期间背面背触毛则在无明显移位的情况下发生退化和瓦解,最终细胞被其分泌形成的包囊壁包围起来,成为圆球形的休眠包囊(图3).
2.2.2 休眠包囊的超微结构
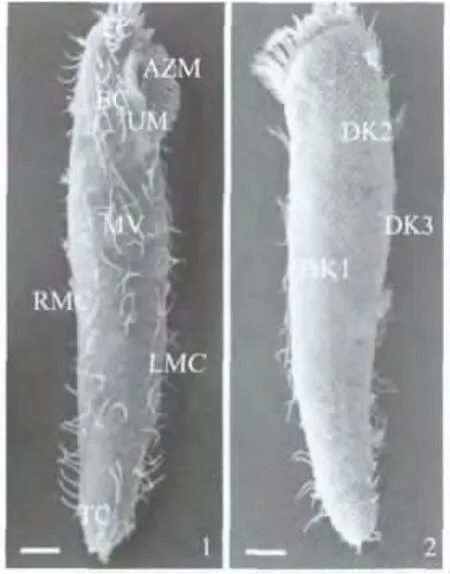
图1 伪红色双轴虫细胞的扫描电镜观察(标尺:10μm)Fig.1 SEM image of vegetative cells of D.pseudorubra(Scale bars:10μm)
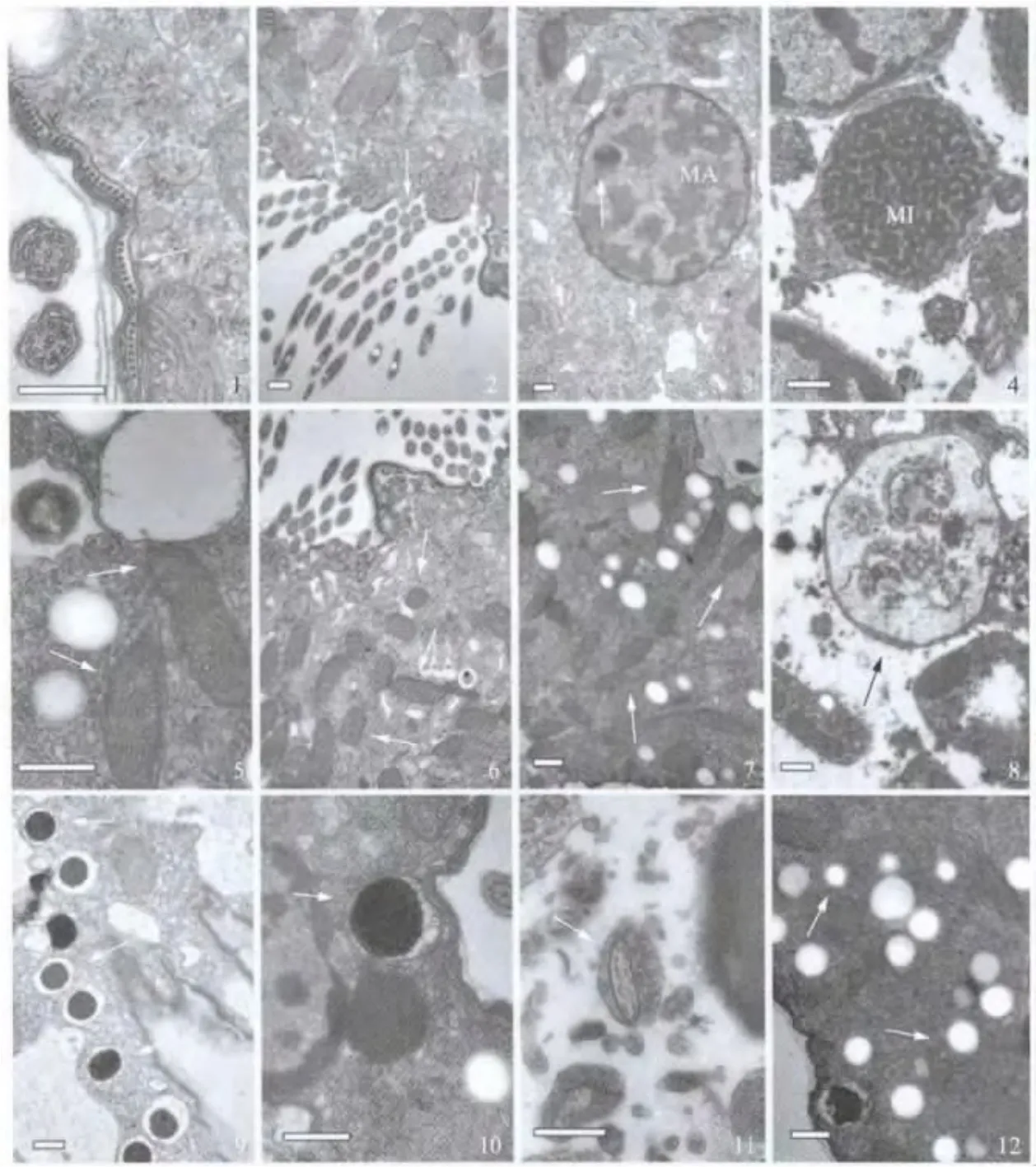
图2 伪红色双轴虫细胞的透射电镜观察(标尺:0.5μm)Fig.2 TEM image of vegetative cells of Diaxonella pseudorubra(Scale bars:0.5 μm)
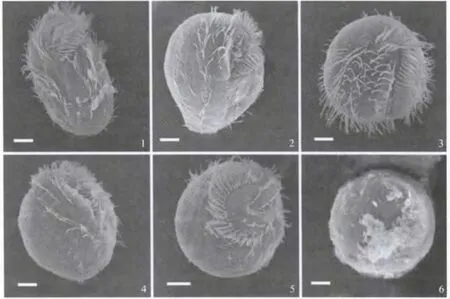
图3 伪红色双轴虫包囊形成时期的扫描电镜观察(标尺:10μm)Fig.3 SEM image of Diaxonella pseudorubra in encystation period(Scale bars:10 μm)
伪红色双轴虫休眠包囊细胞外围的包囊壁包括外壁层,中壁层和颗粒层三层壁结构.其中外层壁厚,壁内含物电子密度低;中壁层电子密度高,壁较薄;内层壁薄,电子密度介于前两者之间(图4:1).休眠细胞的表膜紧贴包囊壁内壁,表膜下部分区段尚存在微管层,但也见部分微管层微管结构已显模糊,或处于退化状态;皮层纤毛器中大部分结构被吸收,但也有少数纤毛结构其纤毛杆被吸收而基体则残留在表膜下细胞质中(图4:2).休眠细胞的大核进一步凝缩,核外除偶见泡状物附着在核膜上外,少见与之相联系的细胞质膜性结构;核内除核仁外,大部分染色质附着在核内膜,仅少部分处于核质中(图4:3);小核核膜清晰可见,核内染色质成均一分布,染色质大小相近(图4:4).
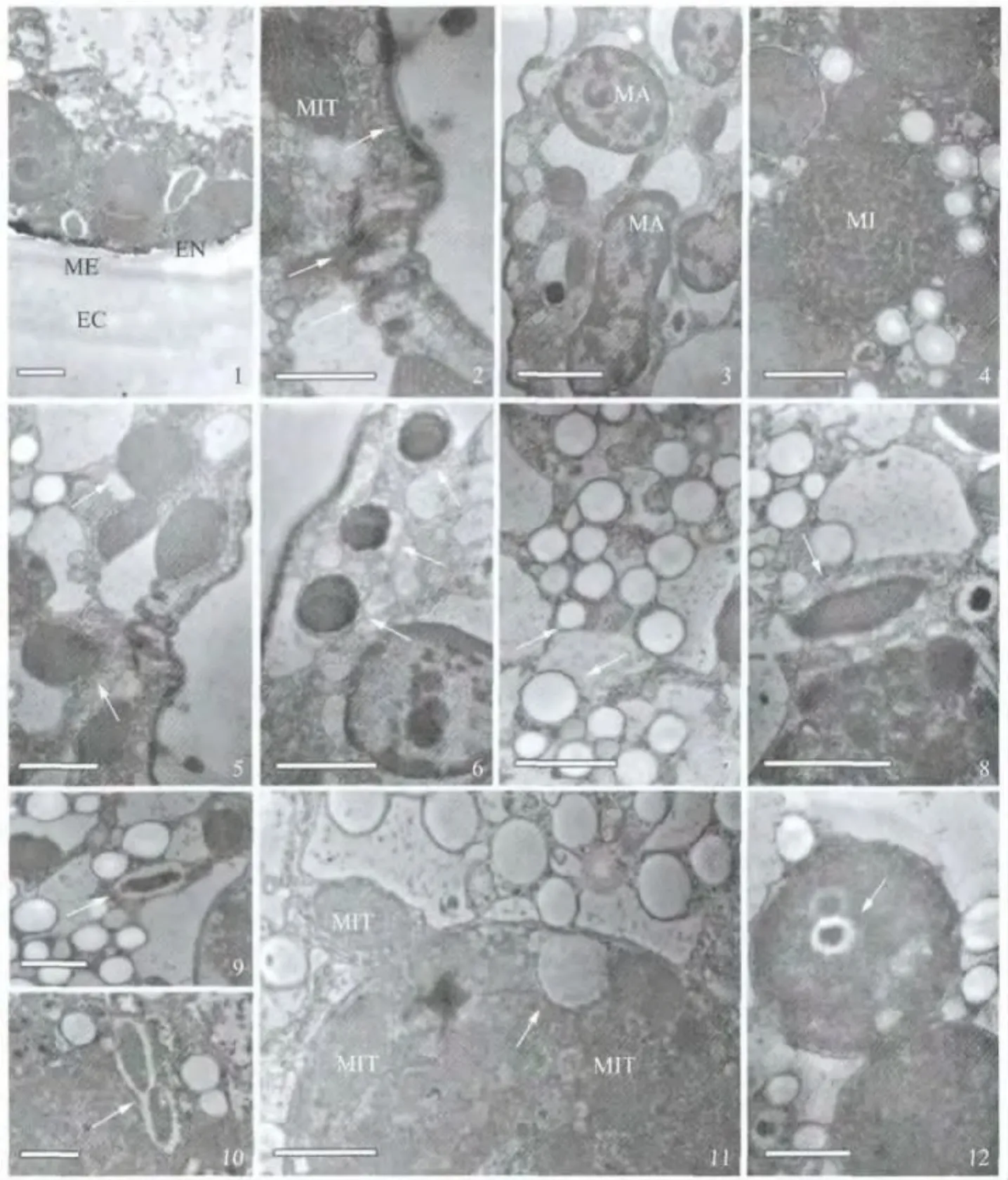
图4 伪红色双轴虫休眠包囊的透射电镜观察(标尺:0.5μm)Fig.4 TEM image of Diaxonella pseudorubra in cysts period(Scale bars:0.5 μm)
其中,线粒体大多分布于细胞表膜下,呈椭圆形和梭形,其内外两层膜及其嵴结构清晰可见(图4:5);黏液泡数量仍较多,形态结构与营养时期非常相似(图4:6);色素颗粒主要以两种形态存在,一种颗粒直径约为0.2μm,另一种颗粒直径约为0.5μm,直径大于0.5μm的颗粒很少见(图4:7);共生细菌外围一层低电子密度的保护层,其菌体呈较高电子密度(图4:8-9),也偶见刚完成分裂并已经分离的前后两个共生菌(图4:10);自噬泡近似圆形,但大小不一(图4:11-12),泡中含有正在被消化的线粒体、色素颗粒(图4:11)和共生菌(图4:12),也见部分泡外的线粒体膜与自噬泡膜发生联系,然后整个线粒体被包裹在自噬泡内,其膜和嵴都逐渐消融,经历被自噬泡消化的过程(图4:11).
3 讨论
3.1 伪红色双轴虫形成包囊的类型及结构特点
前人在对腹毛类纤毛虫形成包囊的研究中观察到,尖毛虫类纤毛虫形成包囊后,其细胞的纤毛结构和表膜下微管全部吸收,形成“毛基体吸收型”包囊[4];游仆虫类纤毛虫形成包囊后,其细胞的大部分纤毛和微管结构被吸收,但部分纤毛基体则被保留下来,形成“毛基体非吸收型包囊”;尾柱虫类纤毛虫形成包囊后,其细胞的大部分纤毛结构被吸收,但保留了少部分的毛基体和微管,形成又一类特殊的“尾柱虫类包囊”[4-5].
本研究工作发现,伪红色双轴虫形成包囊的过程中,细胞内部发生明显的自噬作用,细胞分泌物质形成由内壁层、中壁层和外壁层包裹的三层壁结构,细胞皮层纤毛器中纤毛杆全部被吸收,但一部分基体和表膜下的微管保留下来.根据纤毛虫休眠包囊中皮层纤毛器和微管结构的分化特征,该纤毛虫形成的包囊为“尾柱虫类包囊”.
3.2 伪红色双轴虫自噬泡消化现象
休眠细胞中自噬泡的最重要的作用就是将细胞失去功能和结构的细胞器和细胞颗粒进行消化,为细胞的休眠生理活动提供物质和能量[7].以往的研究中曾报道过在包囊游仆虫(Euplotes encysticus)、魏氏拟尾柱虫(Paraurostyla weissei)和冠突伪尾柱虫(Pseudourostyla cristata)的休眠包囊中观察到细胞内自噬泡消化现象,但尚未观察到自噬泡消化细胞内胞器的具体过程[8-11].本文进一步观察到,伪红色双轴虫休眠细胞中也存在自噬泡消化现象,其中,自噬泡不仅将失去功能的线粒体等膜性细胞器进行消化,也将细胞内色素颗粒、黏液泡及共生菌等包裹在内进行消化,其中值得注意的是,部分失去功能的线粒体在退化过程中能与附近的大自噬泡融合在一起,经历消化过程,这种现象未见报道.所得结果表明,纤毛虫休眠细胞中,自噬泡消化是细胞生命活动中渡过不良环境条件的基本过程,自噬泡消化的对象不仅涉及细胞内胞器,还可能涉及细胞内共生体.
3.3 伪红色双轴虫休眠细胞中其他胞质细胞器和细胞质颗粒的可能作用
与营养期细胞比较,如同其他纤毛虫例如包囊游仆虫那样[12],伪红色双轴虫休眠细胞中小核没有变化,但大核则发生了明显的变化,其两者不同的特征与各自的功能有关,也表明大核在细胞不同生理状态下的功能活动与结构分化的特征;在营养期纤毛虫中线粒体的数量非常多,尤其在纤毛器和其他各种细胞器的功能活动位置线粒体进一步聚集,表明线粒体在细胞生命活动及其能量利用中的作用,但在纤毛虫休眠细胞中线粒体数量减少,这与细胞在休眠生命活动中的代谢及能量需求降低是相一致的,也与细胞调控有关;休眠细胞中的黏液泡、色素颗粒及共生菌呈现与营养细胞中的同种结构相似的形态,尽管三者在纤毛虫营养细胞中行使不同的功能并具有各自的传递机制[13-16],但在形成包囊过程中均没有发生明显的变化,可能是由营养细胞分化为包囊的过程中未发生变化而保留下来的,这也说明所述细胞器或结构在细胞不同生理条件下相对稳定存在的可能原因.但值得注意的是,除细胞核外,在纤毛虫休眠细胞中均观察到线粒体、黏液泡、色素颗粒及共生菌被自噬泡消化的现象,表明这些细胞器或结构在细胞休眠生命活动中可能是细胞内物质利用和能量的主要来源.
[1]顾福康,张作人.纤毛虫形成包囊和脱包囊的研究及其意义[J].动物学杂志,1992,27(5):48-53.
[2]吴月华,季玲妹,顾福康.原生动物细胞休眠现象的研究进展[J].动物学杂志,2004,39(5):91-95.
[3]Verni F,Rosati G.Resting cysts:A survival strategy in Protozoa Ciliophora[J].Italian Journal of Zoology,2011,78(2):134-135.
[4]Walker G K,Mangel T K.Encystment and excystment in hypotrich ciliate.Ⅱ.Diophrys scutum remarks on comparative features[J].Protistologica,1980,16:525-531.
[5]Rios,R M,Torres A.,Calvo P,et al.The cyst of Urostyla grandis(Hypotrichida:Urostylidae):ultrastructure and evolutionary implications[J].Protistologica,1985,21:481-485.
[6]顾福康,倪 兵.原生动物扫描电镜样品制备方法的探讨[J].电子显微学报,1993,12(6):525-529.
[7]Gutiérrez JC,Callejas S,Borniquel S,et al.Ciliate cryptobiosis:a microbial strategy against environmental starvation[J].International Microbiology,2001,4(3):151-157.
[8]顾福康,倪 兵.包囊游仆虫休眠包囊的超微结构研究[J].实验生物学报,1995,28(2):163-166.
[9]顾福康,倪 兵,季玲妹,等.魏氏拟尾柱虫休眠包囊及其细胞器超微结构的观察[J].动物学研究,1999,20(6):206-210.
[10]陈 灵,倪 兵,顾福康.魏氏拟尾柱虫休眠包囊细胞器的电镜酶细胞化学研究[J].动物学研究,2000,21(3):199-203.
[11]顾福康,倪 兵,杨振云,等.冠突伪尾柱虫营养期和形成包囊期间细胞的超微结构[J].动物学报,2002,48(2):251-257.
[12]顾福康,张作人.包囊游仆虫包囊形成和解脱过程中大、小核的研究[J].动物学报,1992,38(2):208-213.
[13]柳伟君,栾菊敏,俞丽丽,等.伪红色双轴虫皮层色素颗粒和黏液泡的显微、亚显微结构观察[J].复旦学报:自然科学版,2009,48(3):381-385.
[14]章 骏,倪 冰,盛 春,等.大尾柱虫粘液泡的超微结构观察[J].复旦学报:自然科学版,2007,46(6):972-975.
[15]Kovács P,Müller WE,Csaba G.A lectin-like molecule is discharged from mucocysts of Tetrahymena pyriformis in the presence of insulin[J].Journal of Eukaryotic Microbiology,1997,44(5):487-491.
[16]周海英,王正君,生 欣,等.Trichototaxis songi细胞皮层色素颗粒的显微和亚显微观察[J].华东师范大学学报,2008,2008(4):66-70.
