草海流域3种优势树种凋落物叶分解历程中的水文特征
2014-09-21张建利严令斌喻理飞蔡国俊
张建利, 严令斌, 喻理飞, 吴 华,3, 蔡国俊
(1.贵州大学 林学院,贵阳 550025; 2.贵州科学院贵州山地资源研究所, 贵阳 550001; 3.开阳县生态文明建设局, 贵阳 550300)
草海流域3种优势树种凋落物叶分解历程中的水文特征
张建利1,2, 严令斌1, 喻理飞1, 吴 华1,3, 蔡国俊1
(1.贵州大学 林学院,贵阳 550025; 2.贵州科学院贵州山地资源研究所, 贵阳 550001; 3.开阳县生态文明建设局, 贵阳 550300)
采用凋落物分解网袋法对草海湿地流域森林优势植物青冈(Cyclobalanopsisglauca)、桤木(Alnuscremastogyne)和云南松(Pinusyunnanensis)凋落物叶分解特性展开研究,揭示凋落物叶分解过程中自然持水率、最大持水率、最大拦蓄率、有效拦蓄率及凋落物持水率规律等。结果表明:(1) 凋落物叶自然持水率随分解时间的增加呈先增后降趋势。凋落物叶自然持水率在分解历程中存在极显著性差异(青冈:F=213.79,P<0.01;桤木:F=77.53,P<0.01;云南松:F=179.12,P<0.01)。(2) 凋落物叶持水率(Rt)与浸水时间(t)呈显著正相关关系(P<0.05),回归分析得到凋落物叶持水率(Rt)与浸水时间(t)符合对数函数方程Rt=a+blnt;凋落物叶持水速率(v)与浸水时间(t)呈显著负相关关系(P<0.05),持水速率(v)与浸水时间(t)符合幂函数方程v=at-b。(3) 凋落物叶最大拦蓄率、有效拦蓄率随分解时间的推移,呈逐渐增加趋势。
优势树种; 凋落物叶; 水文特征
凋落物作为森林生态系统中的重要组成部分之一,其通过分解作用,将大量有机物质及营养元素归还到环境中,分解过程不但可以改善土壤理化性状,而且还可以提升土壤肥力[1-2]。森林凋落物层结构疏松,吸水能力和透水性强,一方面,减缓了降雨对地表的击溅侵蚀,阻滞和分散降水,减少水土流失,另一方面,凋落物能够延缓地表径流,增加土壤水分下渗,对防止土壤侵蚀和涵养水源具有重要作用[3]。因此,凋落物作为评价森林生态系统物质能量平衡和发挥水源涵养的重要环节,越来越受到林学、生态学、微生物学、土壤学等领域的学者专家所重视[4-8]。目前对于森林凋落物的研究多集中在东北和南方地区[9-12],主要针对凋落物发挥水文服务功能、营养物质释放规律及凋落物分解率与环境之间的关系展开研究[13-15],但对凋落物分解历程中的水文特征研究鲜有报道。本研究选择草海湿地流域3种优势树种凋落物叶为研究对象,针对凋落物分解历程中的自然持水率、最大持水率、最大拦蓄率、有效拦蓄率及凋落物持水率特征等问题展开研究,为草海流域森林植物群落水源涵养功能调控及恢复提供科学依据和理论支持。
1 研究区域概况
研究区域位于贵州省威宁县香炉山镇(26°50′17.1″—26°50′20.2″E,104°23′44.1″—104°23′44.5″N)。该区域属于乌蒙山脉山丛中心,具有云贵高原腹地中少有的天然湿地生态系统特征[16]。局部地形由西、南、东三面高原缓丘合围而成,北面逐渐降低为湖盆泄水方向。该区域属于东南季风与西南季风的过度带,草海湿地属长江水系,是金沙江支流横江上洛泽河的上游湖泊,汇集着周围的雨水及几条发源于泉水的短河,具有冬干夏湿,冬暖夏凉的特点,降水集中夏、秋两季,为湿地补给期,冬季相对干旱,湖水有近4个月的补给,小于蒸发和流失的净失水期[17]。草海湿地年均温10.5℃,年均降水量950.9 mm,无霜期208.6 d,湿地底部海拔2 170 m,1981年恢复水面面积19.8 km2,集水区域面积120 km2。年汇水量约800万~900万m3,目前,平均水深1.35 m,最深2.8 m,湿地特征明显[18-19]。草海湿地流域森林植物主要以松科(Pinaceae)、壳斗科(Fagaceae)、杜鹃花科(Ericaceae)、蔷薇科(Rosaceae)禾本科(Gramineae)、菊科(Asteraceae)为主[20]。
2 研究方法
2.1 植物凋落物收集
于2011年11—12月,分别在草海湿地流域范围内的10个典型森林植物群落样地中(如表1所示)收集新近凋落但尚未分解的青冈(Cyclobalanopsisglauca)、桤木(Alnuscremastogyne)、云南松(Pinusyunnanensis)凋落物叶片作为研究对象。将各个收集点采集到的同物种植物凋落物混合均匀,置于65℃烘箱中烘至恒重。采用分析天平将凋落物分装在20 cm×20 cm(孔径1 mm×1 mm)分解网袋中,每袋分装20 g,每种凋落物分装80袋,共计240袋,小心收放(避免物理破碎),用于凋落物分解观测。
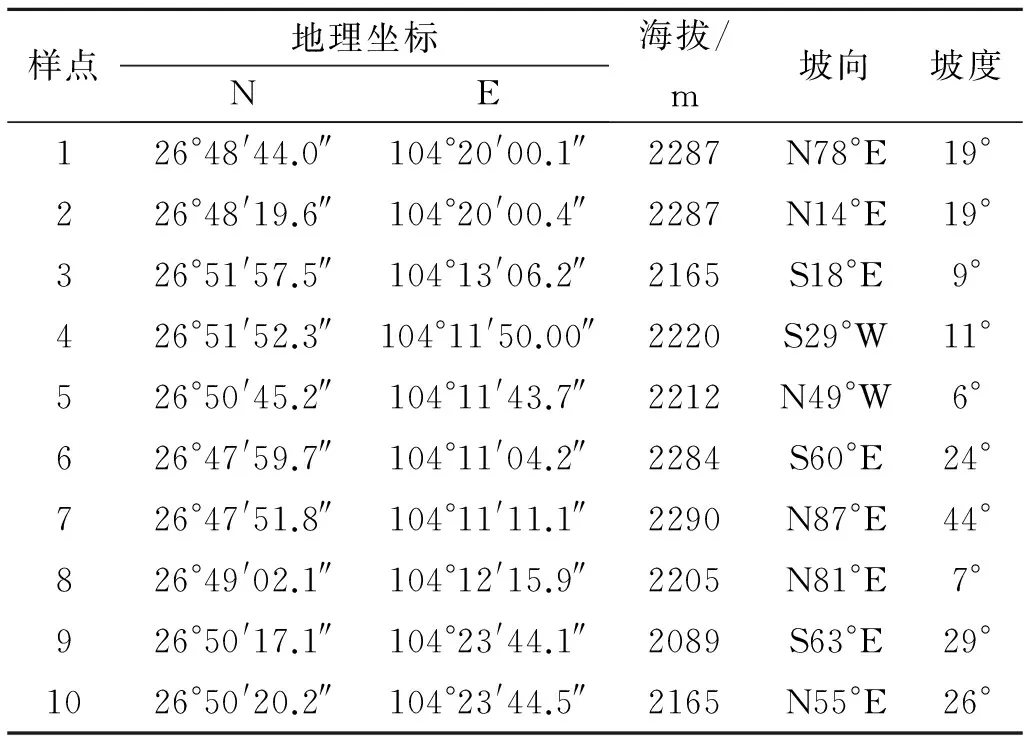
表1 凋落物采集样点描述
注:坡向采用象限角表示;坡度采用坡脚表示。
2.2 投放与回收方法
2012年4月初,在9号凋落物采点(26°50′17.1″E,104°23′44.1″N)设置5个分解场(重复),每个分解场中设置6个分解小区,在分解小区中分别投放每种凋落物16袋,并轻轻掩盖以防止人为破坏[21]。
分别在凋落物投放后的30,60,90,…,360 d后,分别回收每种凋落物5袋,共计15袋,采用自封袋密封保存,带回实验室进行水文特征指标测定。
2.3 统计分析
试验数据采用Microsoft Office Excel 2007软件进行整理,采用SPSS 18.0软件进行方差分析(One-Way ANOVA),采用Sigma plot 12.0软件进行绘图。
2.3.1 凋落物自然持水率 将回收的凋落物网袋带回实验室后,去除泥土和植物根系后,采用分析天平称取其重量,将其置于65℃烘箱内,烘至恒重,称其重量。
凋落物叶自然持水率(%),计算公式为:
式中:Rn——凋落物叶自然持水率(%);Mi0——第i次凋落物叶取样鲜重(g);Mi——第i次凋落物叶取样干重(g)。
2.3.2 凋落物叶持水率、最大持水率、最大拦蓄率、有效拦蓄率 将回收的凋落物叶置于80℃烘箱,烘干至恒重后装入分解网袋中(孔径1mm×1mm),扎好袋口称重,然后将其分别浸入清水中1/12,1/4,1/2,1,2,4,8,12,24,48h后捞起,静置待凋落物不再滴水及时称重(湿重)。
凋落物叶水文特征计算公式[22-23]:
Rlmax=Rmax-Rn
Rsv=0.85Rmax-Rn
式中:Rt——浸水t时间段凋落物叶持水率(%);Mtw——凋落物叶浸水t时间后湿重(g);Md——凋落物叶干重(g);v——凋落物叶持水速率(g/g·h);t——凋落物浸水时间(h);Rmax——凋落物最大持水率(%);M48w——凋落物叶浸水48 h后湿重(g);Rlmax——凋落物最大拦蓄率(%);Rsv——凋落物有效拦蓄率(%);0.85——有效拦蓄系数。
3 结果与分析
3.1 凋落物叶自然持水率特征
对3种不同凋落物叶分解历程中的自然持水率变化规律分析如图1所示。凋落物叶自然持水率随分解时间的增加呈先增加后降低的趋势。
同种凋落物叶分解历程中自然持水率变化如表2所示。青冈凋落物叶分解历程中,不同分解时期的凋落物叶自然持水率间存在极显著性差异(F=213.79;P<0.01)。分解180 d的凋落物叶自然持水率最高,为170.49%±5.23%,比分解90 d的凋落物叶自然持水率增加了53.73%,分解90 d的凋落物叶自然持水率为110.90%±4.94%。分解270 d和360 d的凋落物叶自然持水率分别为63.00%±4.62%,23.80%±1.34%,分别比分解180 d的凋落物叶自然持水率降低了63.04%和86.04%。分解360 d的凋落物叶自然持水率比分解270 d降低了62.22%。
桤木凋落物叶分解历程中,不同分解时期的凋落物叶自然持水率间存在极显著性差异(F=77.53;P<0.01)。分解180 d的凋落物叶自然持水率最高,为141.29%±4.47%,比分解90 d的凋落物叶自然持水率增加了18.58%,分解90 d的凋落物叶自然持水率为119.15%±4.43%。分解270 d和360 d的凋落物叶自然持水率分别为64.95%±8.77%,37.26%±1.40%,分别比分解180 d的凋落物叶自然持水率降低了54.03%和73.63%。分解360 d的凋落物叶自然持水率比分解270 d降低了42.63%。
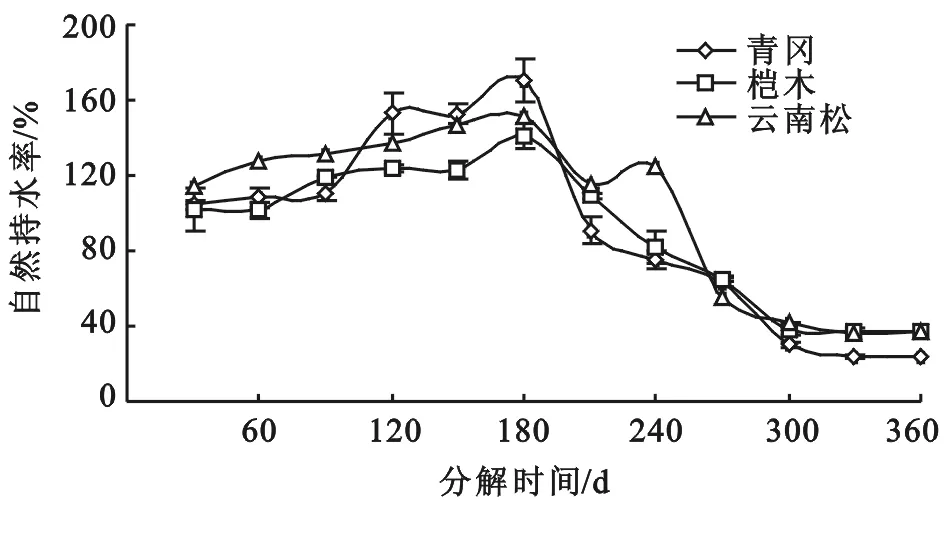
图1 凋落物自然持水率特征
云南松凋落物叶分解历程中,不同分解时期的凋落物叶自然持水率间存在极显著性差异(F=179.12;P<0.01)。分解180 d的凋落物叶自然持水率最高,为151.81%±5.46%,比分解90 d的凋落物叶自然持水率增加了15.14%,分解90 d的凋落物叶自然持水率为131.85%±5.51%。分解270 d和360 d的凋落物叶自然持水率分别为54.96%±2.48%,36.92%±2.21%,分别比分解180 d的凋落物叶自然持水率降低了63.80%和75.68%。分解360 d的凋落物叶自然持水率比分解270 d降低了32.82%。
这一结果表明,不同凋落物叶在分解过程中,其自然持水率变化规律一致,自然持水率表现出趋同现象。
相同分解时期的不同凋落物叶自然持水率分析如表2所示。分解90 d的凋落物叶自然持水率大小关系为云南松>桤木>青冈,不同凋落物叶自然持水率间差异不显著(F=4.49;P=0.06);分解180 d的凋落物叶自然持水率大小关系为青冈>云南松>桤木,不同凋落物叶自然持水率间差异显著(F=8.51;P<0.05);分解270 d的凋落物叶自然持水率大小关系为桤木>青冈>云南松,不同凋落物叶自然持水率间差异不显著(F=0.81;P=0.49);分解360 d的凋落物叶自然持水率大小关系云南松>桤木>青冈,不同凋落物叶自然持水率间差异极显著(F=20.18;P<0.01)。
3.2 凋落物叶持水特征与最大持水率
随着凋落物的分解,凋落物性状发生改变,其水文特征表现各异,凋落物持水率是评价凋落物水文特征的重要指标之一。对3种植物凋落物叶分解历程中的持水率(Rt)与浸水时间(t)进行相关性分析发现,凋落物叶持水率(Rt)与浸水时间(t)呈显著正相关关系(P<0.05),通过回归分析得到凋落物叶持水率(Rt)与浸水时间(t)符合对数函数关系,拟合方程为Rt=a+blnt,不同凋落物叶分解历程中的拟合方程如表3所示。对3种不同植物凋落物叶分解历程中的持水速率(v)与浸水时间(t)进行相关性分析发现,凋落物叶持水速率(v)与浸水时间(t)呈现显著负相关关系(P<0.05),通过回归分析得到凋落物叶持水速率(v)与浸水时间(t)符合幂函数关系,拟合方程为v=at-b,不同植物凋落物叶分解历程中的拟合方程如表4所示。
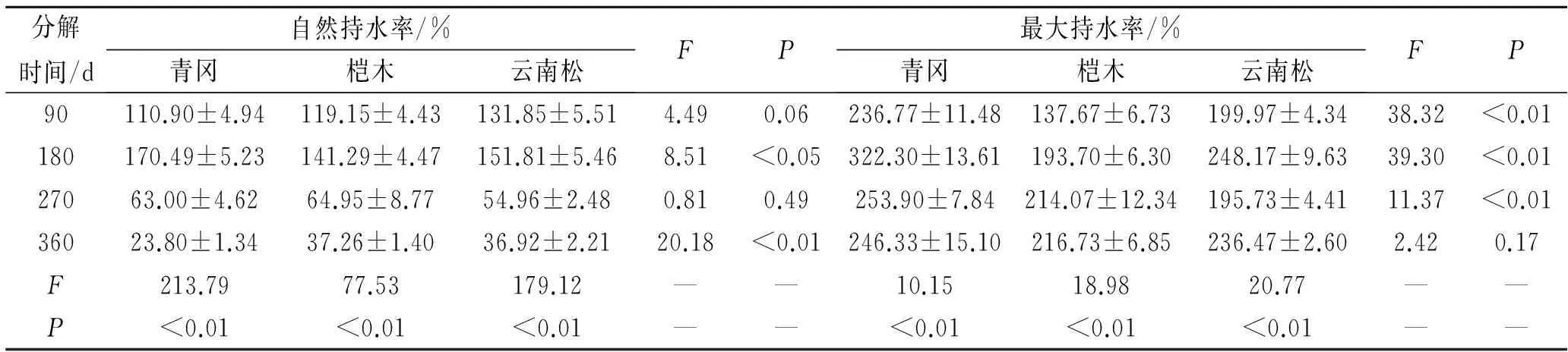
表2 凋落物叶自然持水率和最大持水率特征
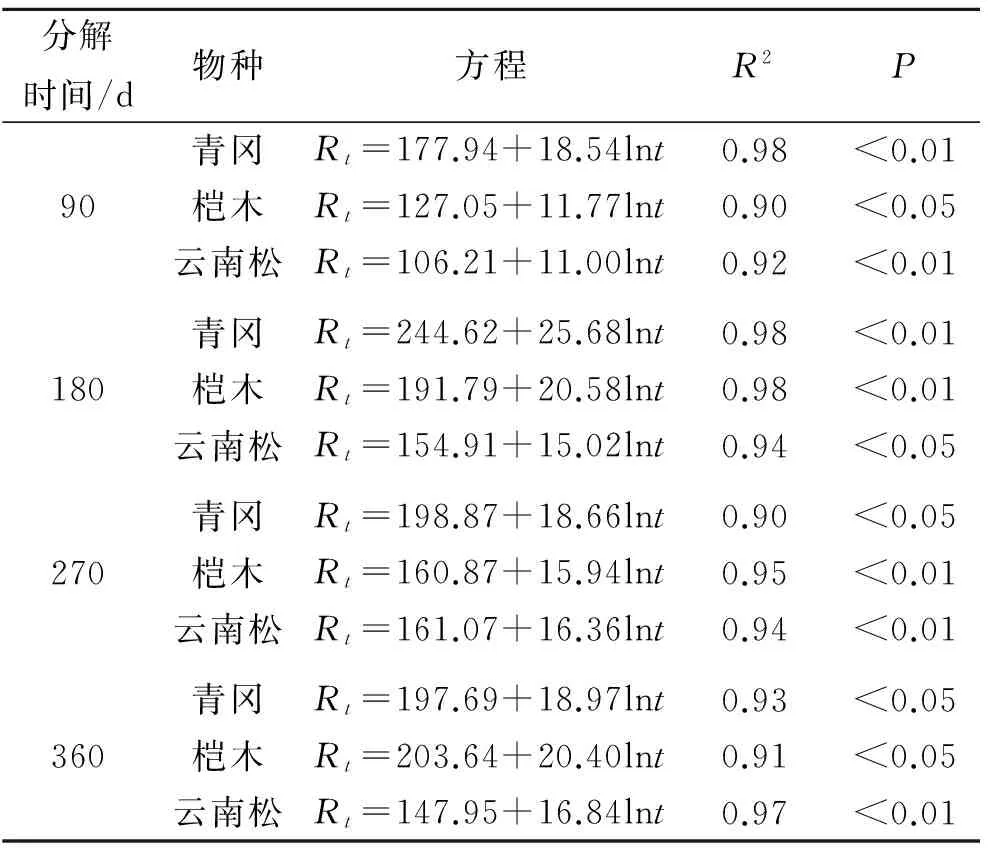
表3 凋落物持水率与浸水时间拟合方程
对3种植物凋落物叶最大持水率分析如表2所示。凋落物叶最大持水率变化规律不明显。对相同植物凋落物叶不同分解历程中的最大持水率分析发现。青冈凋落物叶分解历程中最大持水率呈先增加后降低的趋势,不同分解时期的凋落物叶最大持水率间存在极显著性差异(F=10.15;P<0.01)。分解180 d的凋落物叶最大持水率最高,为322.30%±13.61%,比分解90 d的凋落物叶最大持水率增加了36.12%,分解90 d的凋落物叶最大持水率为236.77%±11.48%。分解270 d和360 d的凋落物叶最大持水率分别为253.90%±7.84%,246.33%±15.10%,分别比分解180 d的凋落物叶最大持水率降低了21.22%和23.57%。分解360 d的凋落物叶最大持水率比分解270 d的凋落物叶最大持水率降低了2.98%。桤木凋落物叶分解历程中,最大持水率呈增加趋势,不同分解时期的最大持水率间存在极显著性差异(F=18.98;P<0.01)。凋落物叶分解360 d的最大持水率最高,为216.73%±13.61%,比分解90 d,180 d,270 d的凋落物叶最大持水率分别增加了57.43%,11.52%,1.24%,分解90 d,180 d,270 d的凋落物叶最大持水率分别为137.67%±6.73%,193.70%±6.30%,214.07%±12.34%。云南松凋落物叶分解历程中,凋落物叶最大持水率变化趋势不明显,不同时期的凋落物叶最大持水率间存在极显著性差异(F=18.98;P<0.01)。凋落物叶分解90 d,180 d,270 d和360 d的最大持水率分别为199.97%±4.34%,248.17%±9.63%,195.73%±4.41%和236.47%±2.60%。
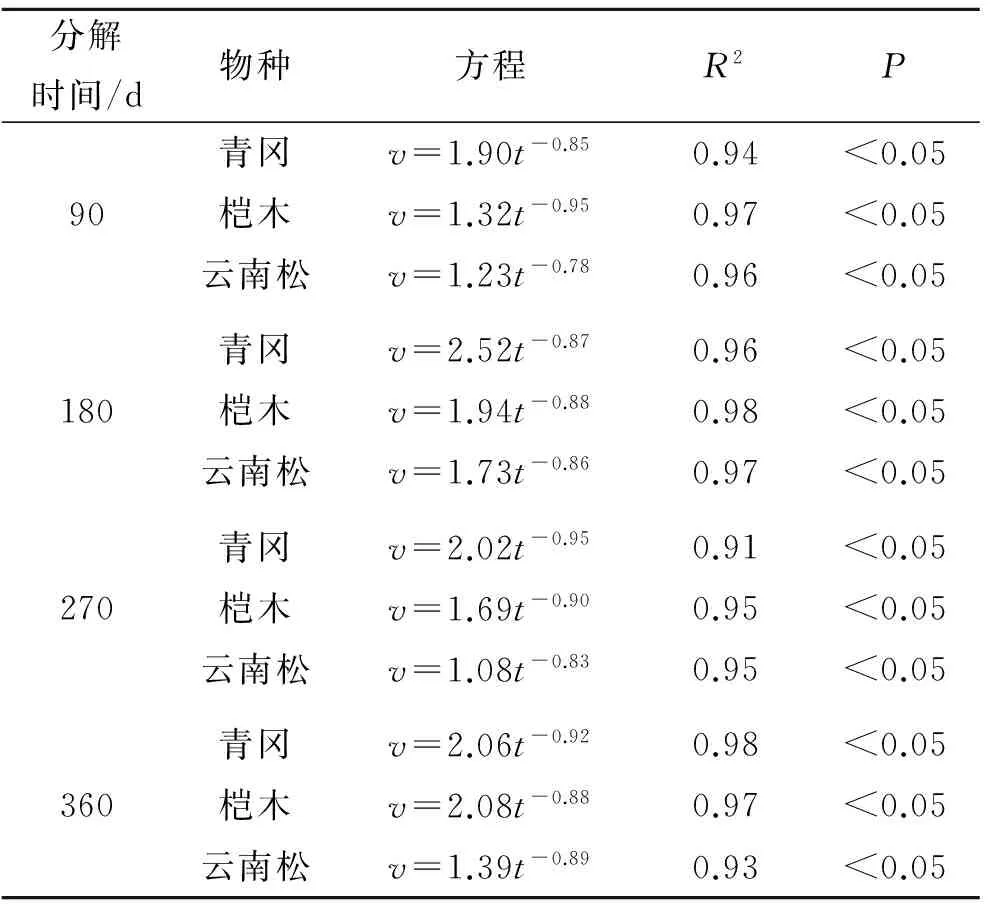
表4 凋落物持水速率与浸水时间拟合方程
相同分解时期的不同植物凋落物叶最大持水率分析如表2所示。分解90 d的凋落物叶最大持水率大小关系为青冈>云南松>桤木,不同植物凋落物叶最大持水率间差异极显著(F=38.32;P<0.01);分解180 d的凋落物叶最大持水率大小关系为青冈>云南松>桤木,不同植物凋落物叶最大持水率间差异极显著(F=39.30;P<0.01);分解270 d的凋落物叶最大持水率大小关系为青冈>桤木>云南松,不同植物凋落物叶最大持水率间差异极显著(F=11.37;P<0.01);分解360 d的凋落物叶最大持水率大小关系为青冈>云南松>桤木,不同植物凋落物叶最大持水率间差异不显著(F=2.42;P=0.17)。
3.3 凋落物叶最大拦蓄率
对3种凋落物叶不同分解历程中的最大拦蓄率分析如表5所示。凋落物叶在分解历程中的最大拦蓄率均呈现增加趋势。
对同种凋落物叶在不同分解期内的最大拦蓄率分析发现(表5)。青冈凋落物叶的最大拦蓄率差异极显著(F=17.66,P<0.01)。分解360 d的凋落物叶最大拦蓄率最高,为222.53%±15.78%,分别比分解90 d,180 d,270 d的凋落物叶最大拦蓄率增加了76.79%,46.59%,16.59%。分解90 d,180 d,270 d的凋落物叶最大拦蓄率分别为125.87%±6.66%,151.80%±14.71%,190.87±8.01%。桤木凋落物叶的最大拦蓄率差异极显著(F=40.40,P<0.01)。分解360 d的凋落物叶最大拦蓄率最高,为179.47%±6.72%,分别比分解90 d,180 d,270 d的最大拦蓄率增加了868.49%,242.50%,20.37%。分解90 d,180 d,270 d的凋落物叶最大拦蓄率分别为18.50%±9.45%,52.40%±9.54%,149.10%±21.10%;云南松凋落物叶的最大拦蓄率差异极显著(F=62.88,P<0.01)。分解360 d的凋落物叶最大拦蓄率最高,为199.60±3.90%,分别比分解90 d,180 d,270 d的凋落物叶最大拦蓄率增加了193.10%,107.12%,41.79%。分解90 d,180 d,270 d凋落物叶最大拦蓄率分别为68.10%±3.35%,96.37%±15.03%,179.47%±6.72%。
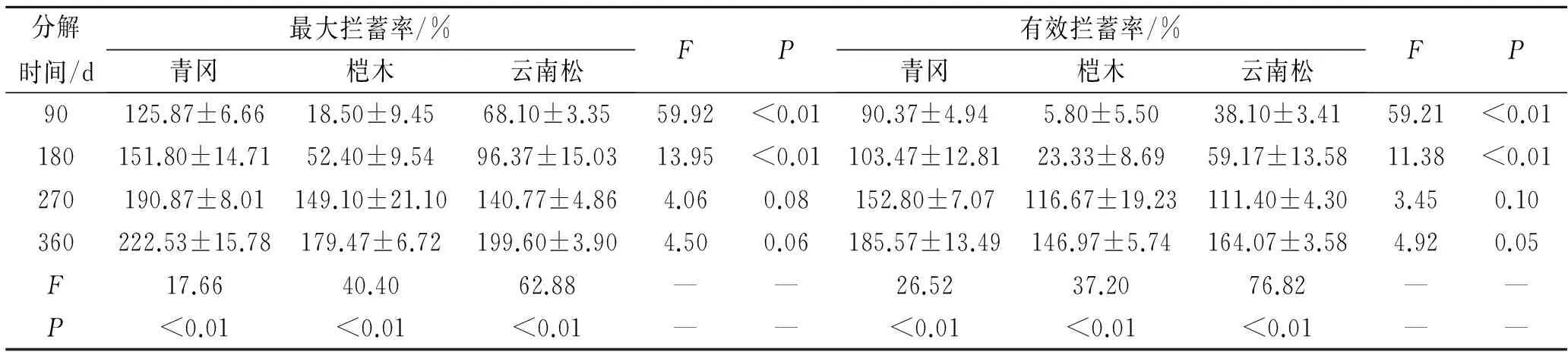
表5 3种凋落物叶最大拦蓄率与有效拦蓄率特征
相同分解时期的不同植物凋落物叶最大拦蓄率分析如表5所示。分解90 d的凋落物叶最大拦蓄率大小关系为青冈>云南>松桤,不同植物凋落物叶最大拦蓄率间差异极显著(F=59.92,P<0.01)。分解180 d的凋落物叶最大拦蓄率大小关系为青冈>云南松>桤木,不同植物凋落物叶最大拦蓄率间差异极显著(F=13.95;P<0.01);分解270 d的凋落物叶最大拦蓄率大小关系为青冈>桤木>云南松,不同植物凋落物叶最大拦蓄率差异不显著(F=4.06;P=0.08);分解360 d的凋落物叶最大拦蓄率大小关系为青冈>云南松>桤木,不同植物凋落物叶最大拦蓄率差异不显著(F=4.50;P=0.06)。
3.4 凋落物叶有效拦蓄率
对3种凋落物叶不同分解历程中的有效拦蓄率分析如表5所示。凋落物叶在分解历程中的有效拦蓄率变化趋势与最大拦蓄率一致,均呈现增加趋势。青冈凋落物叶在不同分解期的有效拦蓄率差异极显著(F=26.52,P<0.01),分解360 d的凋落物叶有效拦蓄率最高,为185.57%±13.49%。桤木凋落物叶在不同分解期的有效拦蓄率差异极显著(F=37.20,P<0.01),分解360 d的凋落物叶有效拦蓄率最高,为146.97%±5.74%。云南松凋落物叶在不同分解期的有效拦蓄率差异极显著(F=76.82,P<0.01),分解360 d的凋落物叶有效拦蓄率最高,为164.07%±3.58%。
凋落物叶解90 d的有效拦蓄率大小关系为青冈>云南松>桤木,有效拦蓄率间差异极显著(F=59.21,P<0.01);分解180 d的凋落物叶有效拦蓄率大小关系为青冈>云南松>桤木,有效拦蓄率间差异极显著(F=11.38,P<0.01);分解270 d的凋落物叶有效拦蓄率大小关系为青冈>桤木>云南松,有效拦蓄率间差异不显著(F=3.45,P=0.10);分解360 d的凋落物叶有效拦蓄率大小关系为青冈>云南松>桤木,有效拦蓄率差间异不显著(F=4.90,P=0.05)。
4 结 论
通过对草海流域3种优势凋落物叶分解历程中的自然持水率、最大持水率、最大拦蓄率、有效拦蓄率以及凋落物持水规律等分析得出:
(1) 凋落物叶自然持水率随分解时间的增加呈现先增加后降低的趋势。凋落物叶自然持水率在分解历程中存在极显著性差异(青冈:F=213.79,P<0.01;桤木:F=77.53,P<0.01;云南松:F=179.12;P<0.01)。不同凋落物叶在分解过程中,自然持水率变化规律一致。
(2) 凋落物叶持水率(Rt)与浸水时间(t)呈显著正相关关系(P<0.05),持水率(Rt)与浸水时间(t)符合对数函数关系Rt=a+blnt;凋落物叶持水速率(v)与浸水时间(t)呈显著负相关关系(P<0.05),持水速率(v)与浸水时间(t)符合幂函数关系v=at-b;凋落物叶最大持水率变化规律不明显。青冈凋落物叶分解历程中最大持水率呈先增加后降低的趋势,分解180 d最大持水率达到峰值(322.30%±13.61%),之后缓慢降低,不同分解时期的凋落物叶最大持水率存在极显著性差异(F=10.15;P<0.01)。桤木凋落物叶分解历程中,最大持水率呈增加趋势,分解360 d后的最大持水率为216.73%±6.85%,不同分解时期的最大持水率存在极显著性差异(F=18.98;P<0.01)。云南松凋落物叶分解历程中,凋落物最大持水率变化趋势不明显,不同时期的凋落物叶最大持水率间存在极显著性差异(F=18.98;P<0.01)。
(2) 凋落物叶在分解历程中的最大拦蓄率、有效拦蓄率均呈现增加趋势。3种凋落物叶分解历程中最大拦蓄率、有效拦蓄率差异均达到极显著水平(P<0.01);分解90 d,180 d的凋落物叶最大拦蓄率、有效拦蓄率差异均达到极显著水平(P<0.01),而分解270 d,360 d的凋落物叶最大拦蓄率、有效拦蓄率差异不著(P≥0.05);分解90 d,180 d,360 d的凋落物最大拦蓄率、有效拦蓄率均呈青冈>云南松>桤木的趋势,而分解270 d的凋落物最大拦蓄率、有效拦蓄率均呈青冈>桤木>云南松的趋势。
[1] Yu S Y, Jian F G, Guang S C et al. Litterfall, nutrient return, and leaf-litter decomposition in four plantations compared with a natural forest in subtropical China[J]. Annals of Forest Science,2004,61(5):465-476.
[2] Garcia-Pausas J, Casals P, Romanyà J. Litter decomposition and faunal activity in Mediterranean forest soils:efects of N content and the mass layer[J]. Soil Biology and Biochemistry,2004,36(6):989-997.
[3] 刘效东,乔玉娜,周国逸等.鼎湖山3种不同演替阶段森林凋落物的持水特性[J].林业科学,2013,49(09):8-15.
[4] 彭少麟,刘强.森林凋落物动态及其对全球变暖的响应[J].生态学报,2002,22(9):1534-1544.
[5] 王清奎,汪思龙,于小军,等.杉木与阔叶树叶凋落物混合分解对土壤活性有机质的影响[J].应用生态学报,2007,18(6):1203-1207.
[6] 刘强,彭少麟.植物凋落物生态学[M].北京:科学出版社,2010.
[7] 汪思龙,陈楚莹.森林残落物生态学[M].北京:科学出版社,2010.
[8] 曾锋,邱治军,许秀玉.森林凋落物分解研究进展[J].生态环境学报,2010,19(1):239-243.
[9] 莫江明,薛璟花,方运霆.鼎湖山主要森林植物凋落物分解及其对N沉降的响应[J].生态学报,2004,24(7):1413-1420.
[10] 魏晶,吴钢,邓红兵.长白山高山冻原生态系统凋落物养分归还功能[J].生态学报,2004,24(10):2211-2216.
[11] 王洪岩,王文杰,邱岭,等.兴安落叶松林生物量、地表枯落物量及土壤有机碳储量随林分生长的变化差异[J].生态学报,2012,32(3):833-843.
[12] 曾从盛,张林海,王天鹅,等.闽江河口湿地植物枯落物立枯和倒伏分解主要元素动态[J].生态学报,2012,32(20):6289-6299.
[13] 李海涛,于贵瑞,李家永,等.井冈山森林凋落物分解动态及磷、钾释放速率[J].应用生态学报,2007,18(2):233-240.
[14] 葛晓改,曾立雄,黄志霖,等.土壤温度和水分含量对三峡库区马尾松林凋落物叶分解的影响[J].林业科学,2013,49(9):153-157.
[15] 刘瑞鹏,毛子军,李兴欢,等.模拟增温和不同凋落物基质质量对凋落物分解速率的影响[J].生态学报,2013,33(18):5661-5667.
[16] 孔凡翠,杨瑞东,林树基.从威宁草海的演化分析0.73Ma来贵州威宁地区喀斯特环境的演变[J].地球与环境,2010,38(2):138-145.
[17] 郭媛,林昌虎,何腾兵,等.草海自然保护区生态环境问题及保护对策[J].贵州科学,2011,29(6):26-30.
[18] 齐建文,李矿明,黎育成,等.贵州草海湿地现状与生态恢复对策[J].中南林业调查规划,2012,31(2):39-40.
[19] 李一兵,彭熙,黄仁海,等.草海湿地保护区土壤重金属及其污染评价[J].贵州科学,2012,30(3):57-62.
[20] 张建利,吴华,喻理飞,等.基于群落数量特征的喀斯特湿地森林群落优势种分析[J].生态环境学报,2013,22(1):58-65.
[21] 张建利,张文,高玲苹,等.云南马龙县山地封育草地凋落物分解与氮释放的研究[J].草业科学,2008,25(7):77-82.
[22] 许文盛,王一峰,童晓霞,等.紫鹊界梯田区植被凋落物水资源调配特性[J].水土保持学报,2013,27(4):130-135.
[23] 常雅军,陈琦,曹靖,等.甘肃小陇山不同针叶林凋落物量、养分储量及持水特性[J].生态学报,2011,31(9):2392-2400.
ResearchforLeaf-LitterHydrographicEffectintheDecompositionProcessofThreeDominantTreeSpeciesintheCaohaiWetlandWatershed
ZHANG Jian-li1,2, YAN Ling-bin1, YU Li-fei1, WU Hua1,3, CAI Guo-jun1
(1.CollegeofForestry,GuizhouUniversity,Guiyang550025,China; 2.InstituteofMountainResourcesofGuizhouProvince,Guiyang550001,China; 3.ConstructionofEcologicalCivilizationBureauofKaiyang,Guiyang550300,China)
The method of leaf litter decomposition bag was used to study natural water-holding rate, maximum water holding rate, maximum interception rate,modified interception rate & holding water trait etc of leaf litter in decomposition process, dominant plant speciesCyclobalanopsisglauca,Alnuscremastogyne&Pinusyunnanensiin Caohai wetland watershed forest were selected as the research examples. The result showed that: (1) the leaf litter natural water-holding rate first increased and then decreased with the decomposition time extension. The leaf litter lnatural water-holding rate was highly significantly different in the decomposition process; (2) the leaf litter water-holding rate (Rt) & immersion time (t) showed a significant positive correlation, the regression analysis for leaf litter water-holding rate (Rt) & immersion time (t) followed the function equationRt=a+blnt. The leaf litter water absorption rate (v) & immersion time (t) showed a significantly negative correlation, the regression analysis for the leaf litter's water absorption rate (v) vs immersion time (t) can be described by the equation,v=at-b; (3) the leaf litter maximum interception rate,modified interception rate gradually increased with the decomposition time duration.
dominant tree species; leaf litter decomposition; hydrographic trait
2014-03-01
:2014-04-25
国家科技支撑计划项目(2011BAC02B02);贵州省科学技术资助项目(黔科合J字[2012]2213号);贵州科学院青年基金重点项目(黔科院J合字[2013]03号);贵州省国际科技合作计划项目(黔科合外G字[2012]7032)。
张建利(1981—),男,甘肃省庆阳市人,博士,助理研究员,主要从事植物生态和喀斯特退化森林生态系统恢复方向研究。E-mail:zhangjl-z@163.com
喻理飞(1963—),男,贵州省贵阳市人,博士,教授,博士生导师,主要从事喀斯特退化森林生态系统恢复方向研究。E-mail:lfyu@gzu.edu.cn
S718.5;Q948
:A
:1005-3409(2014)06-0061-06
