陕北沙地小叶杨“小老树”的水力适应性
2014-09-19陈佳村李秧秧左力翔
陈佳村, 李秧秧, 左力翔
(1. 中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100;2. 中国科学院大学,北京 100049;3. 西北农林科技大学水土保持研究所, 杨凌 712100;4. 西北农林科技大学资源环境学院,杨凌 712100)
土壤水分不足是导致黄土高原“小老树”形成的主要因素[1-2],这些 “小老树”突出表现为:植株生长矮小、顶端枯稍严重、病虫害普遍、生产力低下。探索这些“小老树”的形成机制对于其预防及改造有重要的理论和实践指导价值。以往这方面的研究主要集中在土壤水分的影响和调控上[3],而从植物水分运动特性方面探讨其形成机制极少。
大量研究表明:植物木质部水分传输能力与光合作用之间呈显著正相关[4-5]。“小老树”叶净光合速率和气孔导度显著低于正常水分下生长的树木[6-7],间接表明其水分传输能力受到限制。土壤干旱时植物木质部导管会发生空穴化,从而导致部分输水通道栓塞而失去运输水分的功能,进而影响叶片气孔导度,降低植物的生长速率,严重情况下甚至会造成死亡。杨树是一类需水较多且对干旱敏感的树种,其对空穴化亦很敏感[8-9]。空穴化是导致水滨边生长的杨树树冠出现大量枯枝的重要原因[10],但不同杨树树种抵抗空穴化形成的能力并不相同。迄今为止,关于黄土高原“小老树”分布面积最大的速生树种小叶杨对栓塞的脆弱性尚无报道。植物除通过木质部投资的增加来增强其抵抗空穴化形成的能力以外,还常常通过气孔关闭来使叶水势维持在木质部发生大量栓塞的临界水势之上,从而保持一定的水分传输安全距离(safety margin)[11-12]。因而植物抵抗空穴化能力和气孔调节能力是植物重要的水分关系参数。故本文重点研究了不同水分生境下小叶杨抵抗空穴化形成的能力和气孔调节等水力特性,试图揭示小叶杨对干旱生境的适应机制。
1 材料与方法
1.1 试验区概况
试验在位于陕西省榆林市神木县的六道沟流域(38°46′—38°51′N,110°21′—110°23′E,海拔1094—1274 m)进行。该流域地处黄土高原向毛乌素沙漠的过度地带,属典型的水蚀风蚀交错带。研究区属中温带半干旱气候,冬春季干旱少雨、多风沙,夏秋多雨,多年平均降水量为437.4 mm,70%以上的降水集中在7—9月,年均气温8.4 ℃,≥10 ℃活动积温为3248 ℃,全年无霜期为153 d,年日照时数为2836 h,年总太阳辐射为5922 MJ/m2。 主要土壤类型有绵沙土、新黄土、红土及在沙地发育起来的风沙土和沙地淤土。植被为沙生植物和黄土区植物交错出现,属森林草原带向干草原过度地带[13]。流域内除残存的少量天然灌木如山榆(Ulmusmacrocarpa)、文冠果(Xanthocerassorbifolia)、黄刺玫(Rosexanthina)和绣线菊(Spiraeasalicifolia)等以外,主要以人工林为主,包括柠条(Caraganakorshinskii)、沙柳(Salixpsammophila)、紫穗槐(Amorphafruticosa)等人工灌木林,及小叶杨(Populussimonii)和旱柳(Salixmatsudana)为主的人工乔木林。
1.2 试验设计
试验在2012年8月进行,试验的小叶杨树龄约为38年左右。在水分相对好的沟道(生境A)和干旱的梁坡片沙地(生境B)上布设样地,其中生境A布设1块样地(CK),生境B布设2块样地(S1和S2),样地基本情况见表1。试验开始时每个样地打5钻,用烘干法测定土壤含水量(图1),生境A的CK样地1.6 m以上大部分土层的水分明显好于生境B的两块样地。各样地的土壤表层养分含量无显著差异,0—20 cm有机质含量为1.65—1.82 g/kg,全N为0.16—0.31 g/kg,矿质氮12.4—18.3 mg/kg,Olsen-P为0.53—0.64 mg/kg。整个试验持续近1月,期间有18.7 mm(8月14日)和16 mm(8月17日)的降雨各1次。除脆弱性曲线和气孔导度对叶水势的反应曲线外,其它测定项目均在雨前完成。
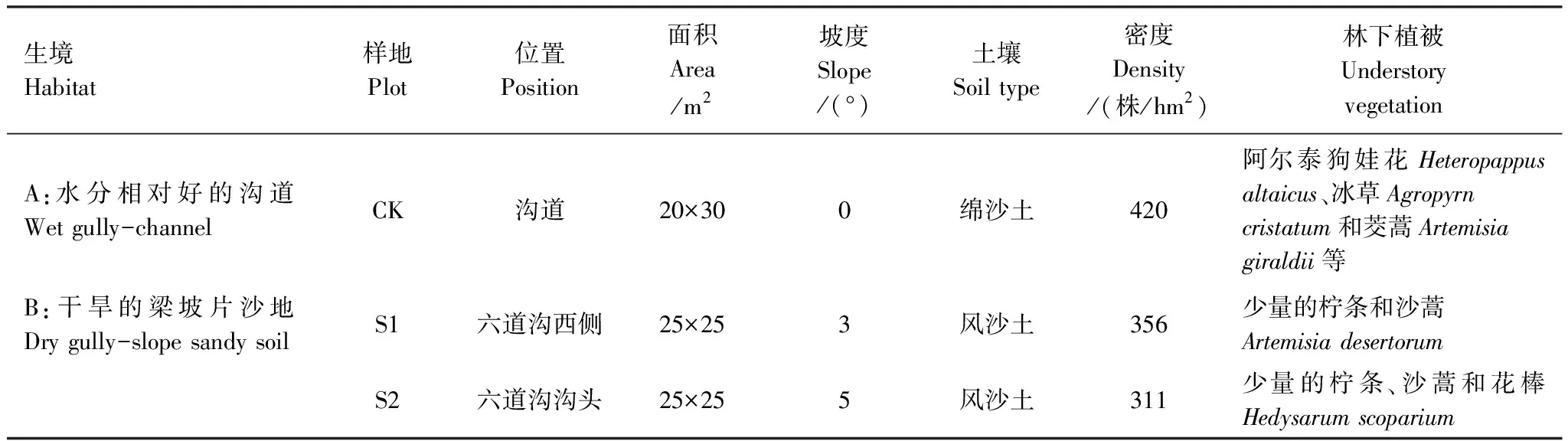
表1 样地基本情况
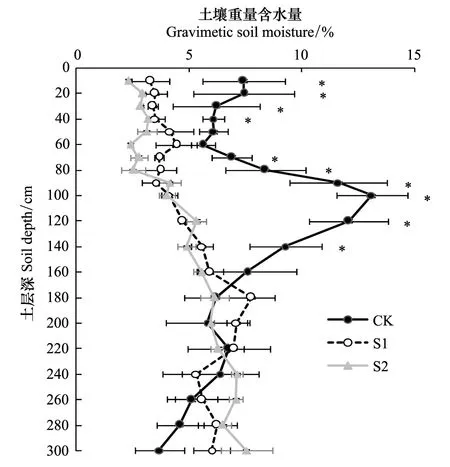
图1 各样地土壤含水量随深度的变化
1.3 测定项目及方法
(1)生长
树高、枯枝起始高度、枯枝结束高度均用测高仪(哈尔滨光学仪器厂,CGQ-1型)测定,枯枝结束高度与起始高度之差为顶端枯枝长度。由于沙地树木在1.3 m以下已分叉形成树冠,故本研究用地径和1 m树高处直径来说明生长状况。每个样地随机调查20棵树。
(2)净光合速率(Pn)、气孔导度(gs)和潜在水分利用效率(WUEi)
每块样地随机选6棵树,每棵树选取树冠南部充分照光的健康叶3片,在晴天10:00—12:00用光合仪(美国CID公司,CI-340型)测量其净光合速率和气孔导度,测定时光强为1000 μmol m-2s-1,CO2浓度为380—400 μmol/mol,温度为28—35 ℃,湿度为30%—40%。其中沙地小叶杨为原位测量,而沟道小叶杨用修枝剪剪取枝条后,在水中剪去末端20 cm 左右,然后放入水中,测量离体光合,5 min内完成。潜在WUEi为净光合速率与气孔导度之比。
(3)黎明前和正午叶水势、正午木质部水势
每块样地随机选6棵树,每棵树随机选取树冠南部健康叶9片,分别用作黎明前叶水势、正午叶水势、正午木质部水势(各3片叶)的测定。黎明前叶水势和正午叶水势分别在4:30—5:30和12:00—14:00用压力室(美国PMS仪器公司,Model 600型)测定;在木质部水势测定前一天下午,用锡箔纸包裹叶,以使叶水势和木质部水势平衡,第2天12:00—14:00测定包裹叶的水势来代表木质部水势。
(4)枝干原位水分传输效率
每块样地随机选5棵树,每棵树在树冠南侧选取3个2—3年生枝条,水中修剪多余枝叶后用移液管法[14]在晴天12:00—14:00测定枝干的导水率(Kh),边材面积(As)用游标卡尺测量去皮枝干直径,然后求得面积(枝干心材面积太小故忽略);叶用扫描仪扫描后,用DT-scan软件计算叶面积(Al)。导水率与边材面积之比为比导水率(Ks),与叶面积之比为比叶导水率(Kl)。Huber值为边材面积与叶面积之比。
(5)脆弱性曲线
采样株数和枝条数与测定枝干原位水分传输效率的相同。采集的枝条用湿报纸包裹后带回实验室,水中修剪多余的枝叶后取25 cm左右长枝干,用空气注入法测定脆弱性曲线[15-16]。所用溶液为20 mmol/L KCl+1 mmol/L CaCl2的混合液。
脆弱性曲线用S型函数
PLC=100/(1+ea(P+b))[17]拟合
式中,PLC为导水率损失百分数(代表了枝干的栓塞程度),P为所用压力,a与曲线斜率有关的常数,b为导水率损失50%所对应的茎木质部水势(P50)。
(6)气孔导度对叶水势的反应曲线
每个样地随机选5棵树。降雨过后的晴天,当一日中气孔导度达到最大时(10:00左右)每棵树砍下3个枝条,砍下的枝条置于田间原位条件下离体慢慢失水,失水过程中同时测定叶水势和气孔导度,直至气孔完全关闭。测定用的水势仪同上,气孔导度用光合仪(美国Li-cor公司,Li-6400型)测定。
1.4 统计分析
不同样地小叶杨生长、光合、水力特性等的差异均用单因素方差分析进行,当差异达到显著(P<0.05)或极显著(P<0.01)时,使用LSD法进行多重比较。所用软件为SPSS 12.0。
2 结果
2.1 生境A和生境B小叶杨的生长与光合
生境B小叶杨的生长状况明显不如生境A。其地径和1 m树高处直径只有生境A小叶杨的1/2左右,其树高仅为1/3左右。生境B小叶杨在1.2—1.5 m就开始有枯稍出现,且其枯稍长度长于生境A,表现出明显的“小老树”特征(表2),但生境B的两块样地小叶杨生长差异不大。
生境B小叶杨的净光合速率显著低于生境A,气孔导度也表现出同样的趋势;尽管生境B小叶杨的潜在WUEi稍高,但它们之间的差异并不达到显著(表3)。生境B中S1样地小叶杨净光合速率高于S2样地,但二者的气孔导度和潜在WUEi差异并不达到显著(表3)。
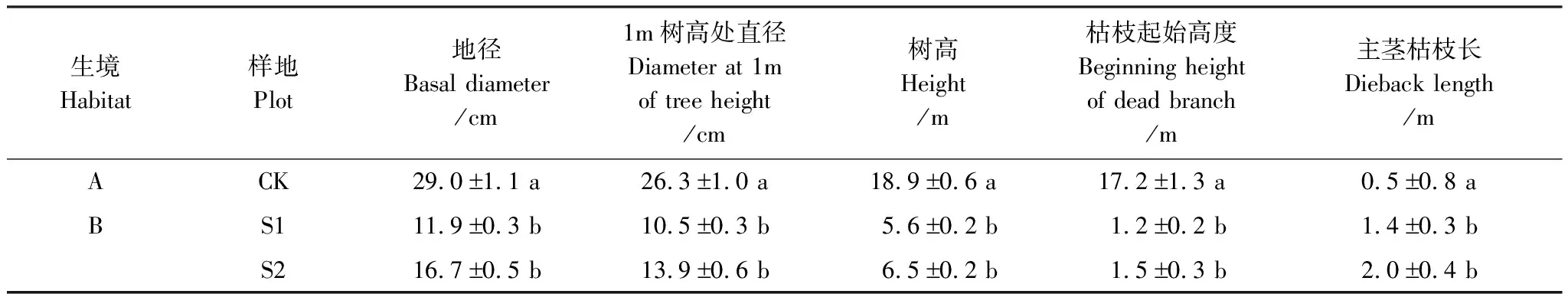
表2 生境A和生境B小叶杨的生长状况
表中数据为平均值标准误差(n=20); 同一列中的小写字母表示样地间差异达到显著水平(P<0.05)
2.2 生境A和生境B小叶杨的叶水势及枝干水分传输效率
生境B小叶杨的黎明前叶水势、正午叶水势和生境A相比无显著差异(图2)。3个样地黎明前叶水势为-0.10 — -0.14 MPa,正午叶水势为-1.64—-1.96 MPa。生境B的 S2样地正午叶水势低于S1样地,并且差异显著。
表4为不同样地上小叶杨田间原位正午的水分传输效率。生境A小叶杨枝干比导水率(Ks)显著高于生境B,但比叶导水率(Kl)并无显著差异,生境B小叶杨的Huber值明显高于生境A。生境B的两块样地之间比导水率、比叶导水率和Huber值并无显著差异。

表3 生境A和生境B小叶杨的光合
表中数据为平均值标准误差(n=6);同一列中的小写字母表示样地间差异达到显著水平(P<0.05)

表4 生境A和生境B小叶杨正午时原位的水分传输效率
表中数据为平均值标准误差(n=5);同一列中的小写字母表示样地间差异达到显著水平(P<0.05)
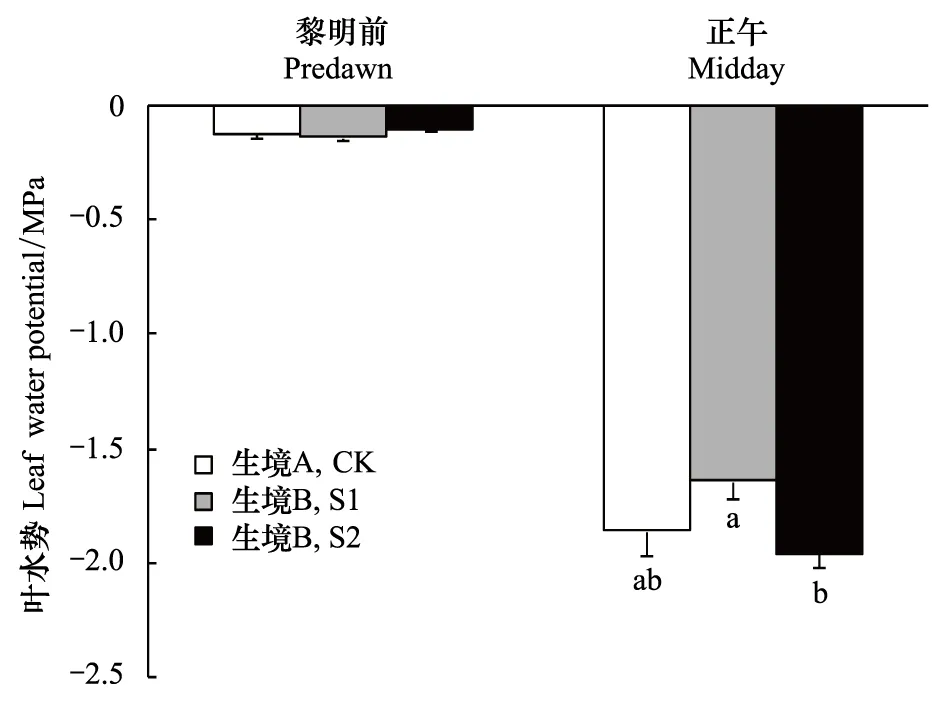
图2 生境A和生境B小叶杨黎明前和正午叶水势
2.3 生境A和生境B小叶杨枝干对栓塞的脆弱性
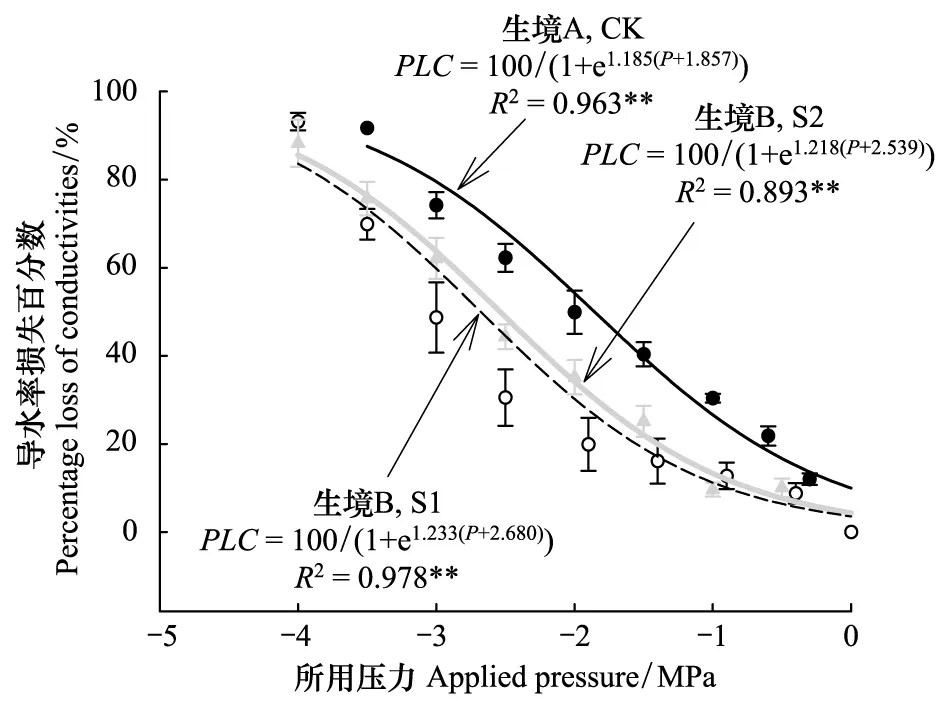
图3 生境A和生境B小叶杨枝干的脆弱性曲线
枝干水分传输能力的降低可能与栓塞形成有关,图3为不同样地小叶杨的脆弱性曲线。从中可看出:S型曲线PLC=100/(1+ea(P+b))较好地模拟了小叶杨的脆弱性曲线,拟合方程都达到了极显著水平。利用模拟函数可求得各样地小叶杨的P50。生境B的两块样地S1和S2的P50分别为-2.68和-2.54 MPa,明显低于生境A的CK样地(-1.85 MPa),表明干旱生境导致小叶杨枝干抵抗空穴化形成能力明显增加,但生境B两个样地之间的P50并无显著差异。将测定的正午木质部水势代入脆弱性曲线模拟方程中,可求得各样地小叶杨正午导水率损失程度。样地CK、S1和S2小叶杨正午枝干导水率损失百分数分别为18.6%、9.2%和8.4%,表明小叶杨枝干正午时的栓塞程度并不高。
2.4 生境A和生境B小叶杨的气孔调节及水分传输安全距离
气孔导度对叶水势的反应曲线用S型函数:
式中,a为最大气孔导度;b与曲线的斜率有关,其值越小,气孔导度下降越快;x0为气孔导度下降50%所对应的叶水势)拟合效果较好,R2均在0.89以上(图4)。生境B小叶杨的气孔导度明显低于生境A(a值,图4)。生境A小叶杨气孔导度的下降速率要超过生境B(b值,图4),表明其气孔对干旱的敏感性要高于生境B。不同生境小叶杨气孔导度下降50%所对应的叶水势无明显差异,均在-1 MPa左右(x0,图4)。生境A的CK样地小叶杨气孔关闭的水势为-1.31 MPa,而生境B的S1和S2样地小叶杨气孔关闭的水势分别为-1.51和-1.54 MPa,生境A小叶杨气孔关闭的水势要早于生境B,但差异只有0.2 MPa左右。样地CK、S1和S2小叶杨的水分传输安全范围(ψsc-P50)分别为0.54、1.17和1.00 MPa(图4),生境B小叶杨水分传输的安全性要明显高于生境A。
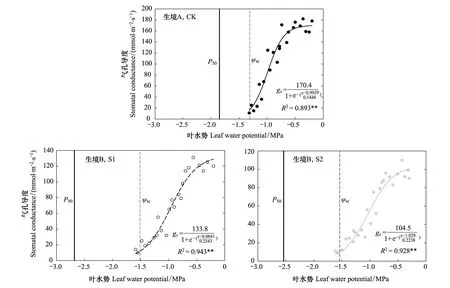
图4 生境A和生境B小叶杨叶气孔导度随叶水势的变化及水分传输安全距离
3 讨论
干旱生境中的小叶杨“小老树”其黎明前叶水势和正午叶水势与正常水分生境中的并无差异,反映了其叶水分状况处于相对供需平衡状态。田间原位水力学性质测定表明:尽管干旱生境中茎水分传输能力降低,但叶水分传输能力并未受到限制,干旱生境中小叶杨有高的Huber值。Huber值反映了承载单位叶面积水分供给的枝干组织量,高的Huber值增加了枝干向叶的水分传输能力,有助于干旱生境中小叶杨叶维持水分平衡,在澳洲生长的桉树上也发现如此[20];另一方面,增加的Huber值有助于降低栓塞的发生程度[21],干旱生境中小叶杨正午时的栓塞程度低于水分好的生境可能也与其高Huber值有关。此外,干旱生境下其它形态学特征,如树高降低、直径减少、冠层上部枯稍增多等均可能有助于维持小叶杨的水分平衡。通过一系列实验证实:水滨边生长的杨树P.deltoids和P.fremontii茎空穴化是导致其枯稍出现的重要原因,并认为枯稍的出现有助于降低蒸腾水分损失,从而使剩余树枝保持较好的水分关系[10]。由于本研究并未单独测定枯枝的水分关系,尚不能证明空穴化是导致枯枝出现的原因。但从正常枝的测定结果看,其水分关系确实并未受到影响。
干旱生境中的物种通常有高的抵抗空穴化能力[22-23],但同一物种干旱后其抵抗空穴化能力的变化常因物种不同而异。对干旱敏感的柳树品种受旱后抵抗空穴化能力稍有增加[24],3个地中海雪松品种中的2个其抵抗空穴化能力对干旱并无显著的反应[25],一些物种如P.angustifolia、P.tremuloides反复干旱后其抵抗空穴化能力明显降低,发生所谓“空穴化疲惫”现象(cavitation fatigue)[26],干旱生境中的假山毛榉属物种茎和叶抵抗空穴化能力均弱于湿润生境中的同种植物[27]。本研究表明:干旱导致小叶杨P50显著降低,这与在其它杨树(毛果杨P.trichocarpa和一种杂交杨P.tremula×P.alba) 上[11,28]的结果一致。生长在沙地上的小叶杨尽管经历反复干旱,但并未出现”空穴化疲惫“现象,可能与小叶杨纹孔膜的微纤丝经受干旱后不易拉伸或断裂从而不易被破坏有关。干旱生境中小叶杨枝干抵抗空穴化能力的增加可能主要与木质部导管大小、密度、导管壁及纹孔膜性质等的改变有关。
除长期的形态结构改变来适应干旱生境外,植物在短时段内常常通过气孔调节来适应干旱。干旱生境中小叶杨气孔导度明显小于沟道小叶杨,在正午叶水势时,两个干旱生境中小叶杨气孔导度仅为沟道小叶杨的17%和4%,气孔关闭导致其光合速率明显降低。本研究中不同生境小叶杨气孔关闭的水势高于木质部发生大量栓塞的水势,表明小叶杨具有较强的气孔调节能力来避免发生大量栓塞。干旱生境中的植物通常有大的水分传输安全距离[19,29],这种大的水分传输安全距离为缓和气孔关闭后水势的进一步下降所必需。本研究中干旱生境的小叶杨也如此,但干旱生境中小叶杨气孔关闭水势比水分生境好的沟道小叶杨仅降低0.2 MPa,表明不同生境中小叶杨气孔调节能力有限,小叶杨主要通过增加木质部抵抗空穴化能力来保证其水分传输的安全性,大的水分传输距离保证了水分传输的安全性,导致光合同化碳更多用来建造抵抗空穴化能力强的木质部,降低了光合碳的投资效率,从而影响生长。因此,从本研究结果看,低的光合速率和为保证水分传输安全性所需的木质部碳投资成本增加是影响小叶杨生长的重要原因。
除木质部水分传输功能和气孔调节外,植物也可能采取其他策略,如落叶、水分贮存、根与茎生物量分配比例的改变、根系吸收深层水比例增加和栓塞修复等来适应干旱,因而有必要对这些策略进行进一步研究,以全面揭示小叶杨“小老树”形成机制。
:
[1]Hou Q C, Huang X, Han S F, Zhang X Z. Study on the forming of “small olded-tree” and the transforming way in the Loess Plateau Ⅰ. the distribution and growth nature of small olded-tree. Acta Conservationis Soli Et Aquae Sinica, 1991, 5(1): 64-72.
[2]Hou Q C, Huang X, Han S F, Zhang X Z. Study on the forming of “small olded-tree” and the transforming way in the Loess Plateau Ⅱ. The status of soil moistures and nutrients in small-old-tree stands and impact on tree growth. Acta Conservationis Soli Et Aquae Sinica, 1991, 5(2): 75-83.
[3]Wang L, Shao M A, Wang Q J, Jia Z K, Li J. Review of research on soil desiccation in the Loess Plateau. Transactions of the Chinese Society of Agricultural Engineering, 2004, 20(5): 27-32.
[4]Hubbard R M, Stiller V, Ryan M G, Sperry J S. Stomatal conductance and photosynthesis vary linearly with plant hydraulic conductance in ponderosa pine. Plant Cell and Environment, 2001, 24(1): 113-121.
[5]Brodribb T J, Field T S. Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests. Plant Cell and Environment, 2000, 23(12): 1381-1388.
[6]Han R L, Hou Q C. Water metabolism characters and photosynthesis capacity of old smallPopulustrees in growth season. Journal of Northwest Forestry College, 1996, 11(3): 36-40.
[7]Tanaka-Oda A, Kenzo T, Koretsune S, Sasaki H, Fukuda K. Ontogenetic changes in water-use efficiency (δ13C) and leaf traits differ among tree species growing in a semiarid region of the Loess Plateau, China. Forest Ecology and Management, 2010, 259(5): 953-957.
[8]Tyree M T, Kolb K J, Rood S B, Patiňo S. Vulnerability to drought-induced cavitation of riparian cottonwoods in Alberta: a possible factor in the decline of the ecosystem? Tree Physiology, 1994, 14(5): 455-466.
[9]Cochard H, Casella E, Mencuccini M. Xylem vulnerability to cavitation varies among poplar and willow clones and correlates with yield. Tree Physiology, 2007, 27(12): 1761-1767.
[10]Rood S B, Patiňo S, Coombs K, Tyree M T. Branch sacrifice: cavitation-associated drought adaptation of riparian cottonwoods. Trees, 2000, 14(5): 248-257.
[11]Sparks J P, Black R W. Regulation of water loss in populations ofPopulustrichocarpa: the role of stomatal control in preventing xylem cavitation. Tree Physiology, 1999, 19(7): 453-459.
[12]Sperry J S, Hacke U G. Desert shrub water relations with respect to soil characteristics and plant functional type. Functional Ecology, 2002, 16(3): 367-378.
[13]Wang J G, Fan J, Wang Q J, Wang L. Vegetation above-ground biomass and its affecting factors in water/wind erosion crisscross region on Loess Plateau. Chinese Journal of Applied Ecology, 2011, 22(3): 556-564.
[14]Melcher P J, Holbrook N M, Burns M J, Zwieniecki M A, Cobb A R, Brodribb T J, Choat B, Sack L. Measurements of stem xylem hydraulic conductivity in the laboratory and field. Methods in Ecology and Evolution, 2012, 3(4): 685-694.
[15]Sperry J S, Saliendra N Z. Intra-and Inter-plant variation in xylem cavitation inBetulaoccidentalis. Plant, Cell and Environment, 1994, 17(11): 1233-1241.
[16]Zuo L X, Li J H, Li Y Y, Zhao L M. Comparison of hydraulic traits in branches and leaves of diffuse-and ring-porous species. Acta Ecologica Sinica, 2012, 32(16): 5087-5094.
[17]Pammenter N W, Willigen C V. A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation. Tree Physiology, 1998, 18(8/9): 589-593.
[18]Brodribb T, Hill R S. The importance of xylem constraints in the distribution of conifer species. New Phytologist, 1999, 143(2): 365-372.
[19]Mayr S, Beikircher B, Obkircher M A, Schmid P. Hydraulic plasticity and limitations of alpineRhododendronspecies. Oecologia, 2010, 164(2): 321-330.
[20]Carter J L, White D A. Plasticity in the Huber value contributes to homeostasis in leaf water relations of a mallee Eucalypt with variation to groundwater depth. Tree Physiology, 2009, 29(11): 1407-1418.
[21]DeLucia E H, Maherali H, Carey E V. Climate-driven changes in biomass allocation in pines. Global Change Biology, 2000, 6(5): 587-593.
[22]Pockman W T, Sperry J S. Vulnerability to xylem cavitation and the distribution of Sonoran desert vegetation. American Journal of Botany, 2000, 87(9): 1287-1299.
[23]Maherali H, Pockman W T, Jackson R B. Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology, 2004, 85(8): 2184-2199.
[24]Wikberg J,gren E. Variation in drought resistance, drought acclimation and water conservation in four willow cultivars used for biomass production Tree Physiology, 2007, 27(9): 1339-1346.
[25]Ladjal M, Huc R, Ducrey M. Drought effects on hydraulic conductivity and xylem vulnerability to embolism in diverse species and provenances of Mediterranean cedars. Tree Physiology, 2005, 25(9): 1109-1117.
[26]Hacke U G, Stiller V, Sperry J S, Pittermann J, McCulloh K A. Cavitation fatigue: embolism and refilling cycles can weaken the cavitation resistance of xylem. Plant Physiology, 2001, 125(2): 779-786.
[27]Bucci S J, Scholz F G, Campanello P I, Montti L, Jimenez-Castillo M, Rockwell F A, Manna L L, Guerra P, Bernal P L, Troncoso O, Enricci J, Holbrook M N. Hydraulic differences along the water transport system of South AmericanNothofagusspecies: do leaves protect the stem functionality? Tree Physiology, 2012, 32(7): 880-893.
[28]Awad H, Barigah T, Badel E, Cochard H, Herbette S. Poplar vulnerability to xylem cavitation acclimates to drier soil conditions. Physiologia Plantarum, 2010, 139(3): 280-288.
[29]Holloway-Phillips M M, Brodribb T J. Contrasting hydraulic regulation in closely related forage grasses: implication for plant water use. Functional Plant Biology, 2011, 38(7): 594-605.
参考文献:
[1]候庆春, 黄旭, 韩仕峰, 张孝中. 黄土高原地区小老树成因及其改造途径的研究. Ⅰ 小老树的分布及其生长特点. 水土保持学报, 1991, 5(1): 64-72.
[2]候庆春, 黄旭, 韩仕峰, 张孝中. 黄土高原地区小老树成因及其改造途径的研究. Ⅱ 土壤水分和养分状况及其与小老树生长的关系. 水土保持学报, 1991, 5(2): 75-83.
[3]王力, 邵明安, 王全九, 贾志宽,李军. 黄土区土壤干化研究进展. 农业工程学报, 2004, 20(5): 27-32.
[6]韩蕊莲, 候庆春. 小叶杨“小老树”生长季内水分特征及光合能力. 西北林学院学报, 1996, 11(3): 36-40.
[13]王建国, 樊军, 王全九, 王力. 黄土高原水蚀风蚀交错区植被地上生物量及其影响因素. 应用生态学报, 2011, 22(3): 556-564.
[16]左力翔, 李俊辉, 李秧秧, 赵丽敏. 散孔材与环孔材树种枝干、叶水力学特性的比较研究. 生态学报, 2012, 32(16): 5087-5094.
