连续单一除草剂应用情况下的转基因直播稻田杂草群落动态
2014-08-30张晶旭戴伟民
张晶旭, 戴伟民, 强 胜
南京农业大学杂草研究室,江苏 南京 210095
水稻是世界上最重要的粮食作物之一,全球人均1/4的食物热量来自水稻(Macleanetal., 2003)。中国则是世界最大的水稻生产国,2012年水稻种植面积超过3000万hm2,产量达到20423万t(中华人民共和国国家统计局,2013)。然而水稻生产一直受到杂草的威胁,每年因杂草危害造成的产量损失超过1000万t稻谷(强胜,2010;于改莲,2001)。化学除草剂是稻田杂草防除最主要的技术手段(强胜,2009),轻型栽培模式下的水稻,需要使用2~3次选择性除草剂才能免除杂草危害,以确保稳产和高产。但是除草剂的使用无疑给农民带来生产成本的增加,其费用甚至高达每公顷上千元,同时也增加了环境污染的压力和人们对食品安全的担忧(强胜和马波,2004)。通过转基因技术培育抗除草剂水稻,可在水稻生长期间使用灭生性除草剂如草甘膦、草铵膦等,从而扩大除草剂的杀草谱,减少或免去针对类似禾本科恶性杂草的专门选择性除草剂的应用。因此种植转基因作物有降低生产成本,改善产品品质及降低生态环境污染等优点。自转基因作物1996年商业化以来种植面积不断扩大,2013年全球的种植面积已经达到1.75亿hm2,其中抗除草剂转基因作物及包含除草剂抗性的复合性状转基因作物的种植面积比例之和超过80%(James,2014)。
抗除草剂转基因水稻的商业化种植势必会导致除草剂的使用更加单一,这种单一除草剂的长期使用更有可能导致抗/耐药性杂草的演化(卢宝荣等,2008; 左娇等,2013; Rajguruetal.,2005; Gainesetal.,2010)。抗性杂草的形成将导致杂草群落演替,这已成为我国杂草成灾的主要原因之一(强胜,2010),其最终结果将可能导致除草剂技术体系的失效(强胜,2001),这已成为转基因抗除草剂作物发展的重要生物安全性问题。由于转基因抗除草剂水稻还未商业化,种植过程中灭生性除草剂使用后的杂草群落结构动态的研究也较少,更没有在长期连续使用下群落动态的报道(余柳青等,2005)。为了明确在转基因抗除草剂水稻栽培中长期单一使用灭生性除草剂对稻田杂草群落动态的影响,自2011~2013年,连续3年在种植抗草铵膦转基因水稻Bar68-1的田中使用灭生性除草剂草铵膦以及常规选择性除草剂丙草胺—苄嘧磺隆,比较观察稻田杂草群落结构,以期揭示单一灭生性除草剂应用情况下稻田杂草群落的动态变化规律,为转基因抗除草剂水稻的产业化提供生物安全的理论依据。
1 材料和方法
1.1 试验材料
抗草铵膦转基因水稻Bar68-1,以基因枪法将草铵膦抗性基因Bar转入受体材料D68得来,对叶面喷施草铵膦表现出抗性。中国科学院亚热带农业生态研究所提供;
草铵膦(Glufosinate)(有效浓度为200 g·L-1的水剂);
丙草胺—苄嘧磺隆(Bensulfuron-methyl+Pretilachlo,后简写为丙·苄,BMP)(35%的可湿性粉剂,其中苄嘧磺隆含量2%,丙草胺含量33%)。
1.2 试验设计
于2011~2013年在南京农业大学杂草研究室转基因水稻安全评估试验基地进行。采用水直播栽培方式种植转基因水稻Bar68-1。试验设置常规除草剂丙·苄处理(水直播10 d后用药,有效浓度236.25 g·hm-2)、灭生性除草剂草铵膦处理(水直播40 d后用药,有效浓度450 g·hm-2)和空白对照(不使用任何除草措施)。每种处理4个重复,每个小区面积50 m2。
1.3 调查方法及数据分析
1.3.1 杂草调查方法 使用样框法调查杂草群落,每小区对角线5点取样,样框大小0.25 m2,记录每个样框中的杂草种类及每种杂草数量。水稻水直播10 d后进行第1次调查,之后每隔10 d调查1次,每年共调查7次。每年的调查结果用作反映当年2类除草剂对杂草的防除效果。每年最后一次杂草调查结果作为每年的终杂草群落数据。综合3年终杂草群落的数据反映长期使用单一除草剂对杂草群落演替的影响。水稻成熟后将全部水稻收割脱粒测产,随后焚烧销毁。
1.3.2 数据处理分析 杂草密度为杂草株数·m-2;物种丰富度即样方中包含的所有杂草种类数;相对优势度RI=(RD+RF)/2;
其中,RD为相对密度,即某杂草的密度占总杂草密度的比例,RF为相对频度,即某小区杂草出现的样方数占总样方数的比例(Cardinaetal.,2002; Shresthaetal.,2002);
物种多样性指数包括Margalef丰富度指数(Margalef,1958)、Shannon-Wiener指数(Putman & Wratten,1984)、Simpson优势度指数(Parishetal.,1994)及Pielou均匀度指数(Hill,1973)。
Margalef丰富度指数:R=(S-1)/lnN;
Shannon-Wiener指数:H′=-∑Pi·lnPi;
Simpson优势度指数:D=1-∑Pi2;
Pielou均匀度指数:E=H′/lnS。
上述公式中的S为杂草种类总数,Pi=Ni/N,N为样方中个体总数,Ni为样方中第i物种的个体数。
群落相似性指数用Bray-Curtis指数(Bray & Curtis, 1957)及Jaccard相似指数测定(Milneretal.,2008)。
其中xik指第k个物种在年份i中的个体数目,xjk含义与之类似,P指的是样方中的物种总数。
Jaccard相似性指数:Cj=c/(a+b-c);
其中a为群落A含有的全部物种数,b为群落B含有的全部物种数,c为两群落共有的物种数。
使用Excel,Origin进行数据处理、绘图,并使用软件SPSS、软件R进行统计分析并检验各处理间的差异显著性。
2 结果与分析
2.1 草铵膦和丙·苄的除草效果及对产量影响
如图1所示,使用2种除草剂处理小区的总杂草密度均显著小于不除草CK处理小区。使用常规除草剂丙·苄可明显的抑制杂草发生,总杂草发生量明显减少。但是随着使用年份增加,用药后期杂草发生量有上升趋势,终总杂草密度最高超过500株·m-2。稻田中总杂草密度在水直播后30~40 d时达到最高峰。在杂草密度达到最高峰后使用灭生性除草剂草铵膦,能达到理想的除草效果,且后期持续维持这种状况,最终总杂草密度显著下降到最低水平108株·m-2。3年结果比较,年际间趋势差别不大。
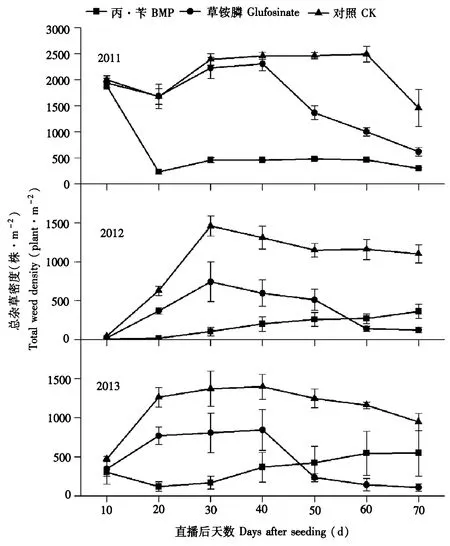
图1 不同除草剂处理对总杂草发生密度的影响Fig.1 Effects of the 2 herbicide treatmenst on the density of weeds
如图2所示,2种除草剂处理小区在2011~2013年的水稻产量均显著高于不除草CK处理小区。2011年草铵膦和丙·苄处理小区的水稻产量无显著差异,而2012、2013年草铵膦处理的水稻产量显著高于丙·苄处理组。该结果与总杂草密度显示的结果相对应。说明连续使用草铵膦可持续达到较好的除草效果,确保水稻产量正常,而使用常规选择性除草剂丙·苄,则随着用药年限的增加,由于后期多年生杂草发生量增加而对水稻产量的影响变大。
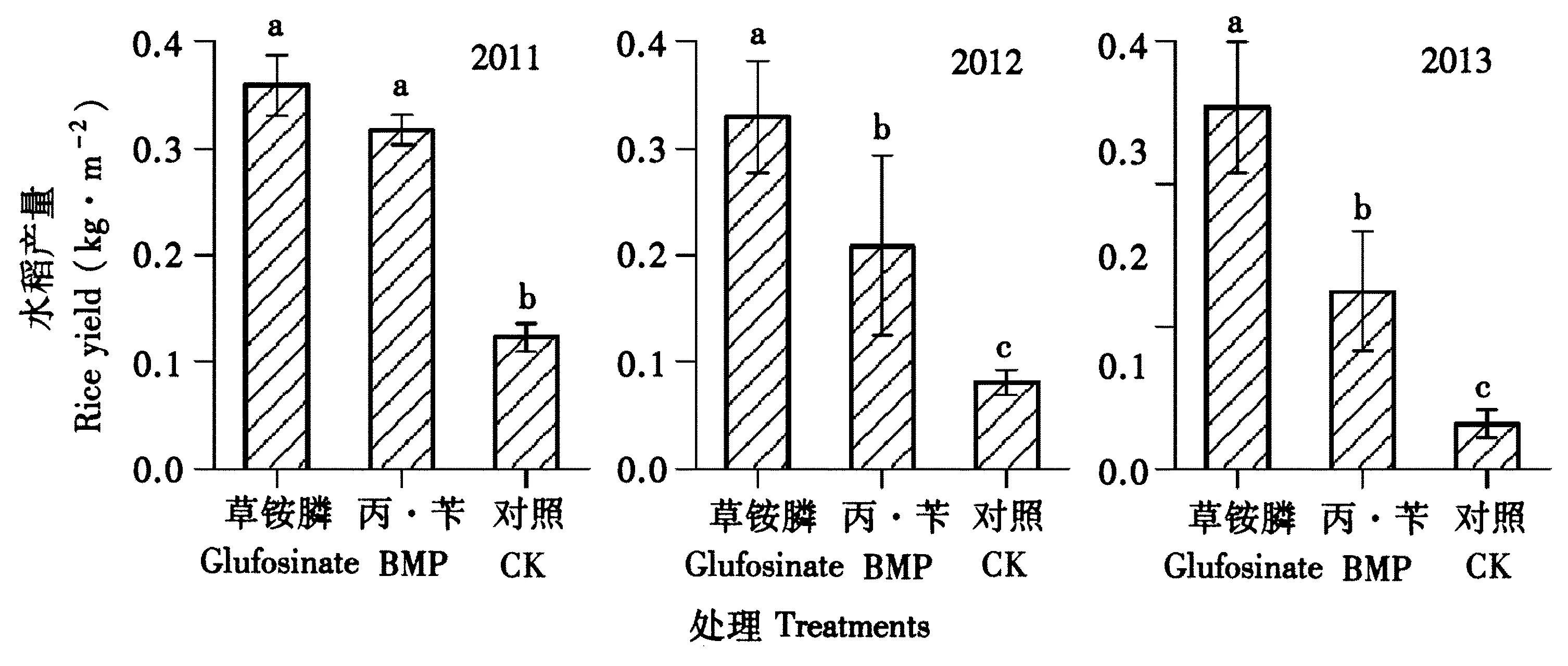
图2 不同除草剂处理小区水稻产量Fig.2 Rice yields exposed to different herbicide treatments
2.2 单一除草剂连续使用对物种丰富度和总杂草密度的影响
如图3所示,经过连续3年的单一使用灭生性除草剂草铵膦或常规除草剂丙·苄都使田间杂草物种丰富度呈下降趋势,而不使用任何除草措施的CK组杂草物种丰富度相对稳定。2种除草剂处理的终总杂草密度均显著低于CK,其中草铵膦处理和CK组呈逐年降低趋势,而丙·苄处理呈逐年上升趋势。
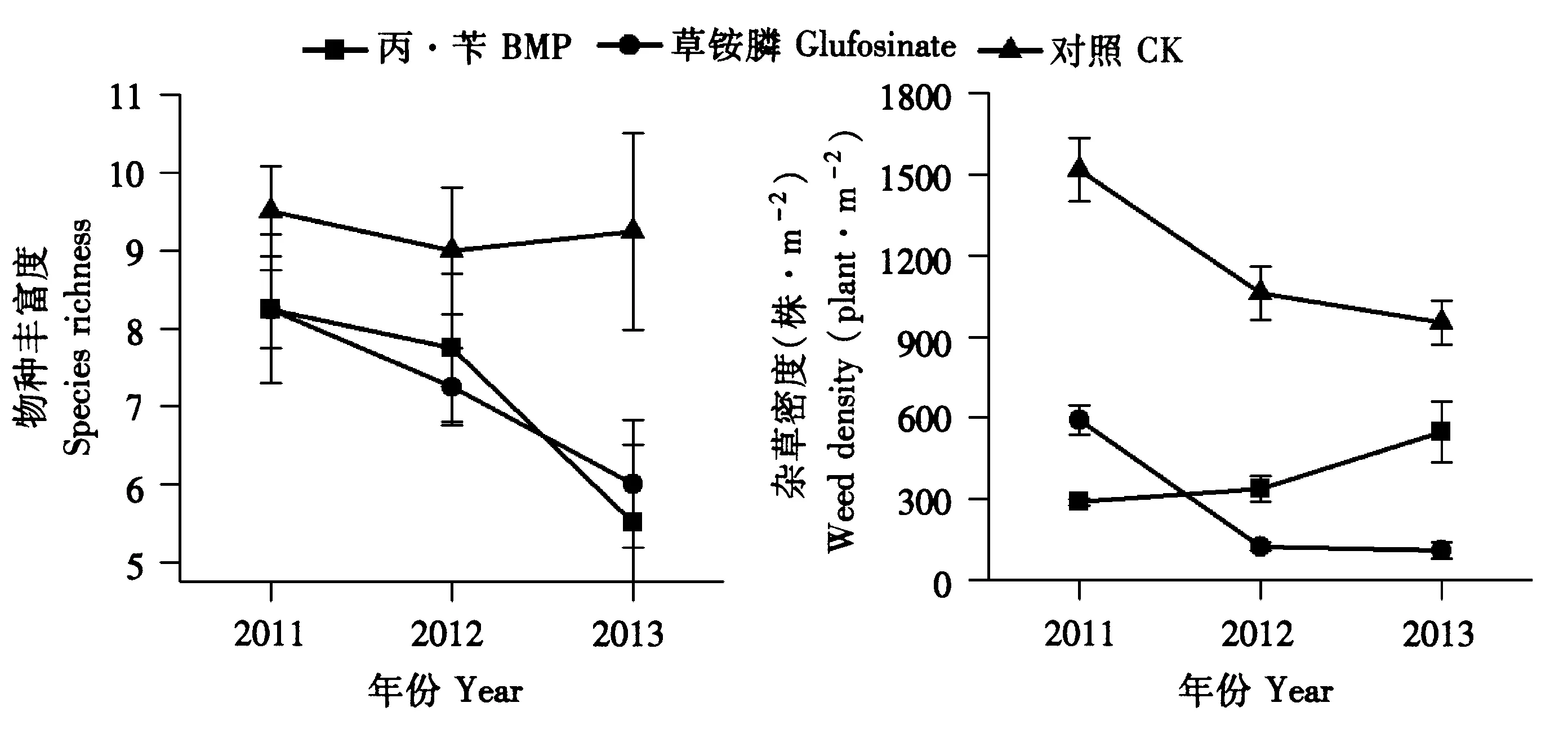
图3 长期使用单一除草剂对物种丰富度和总杂草密度的影响Fig.3 Effects of single herbicide for long-term usage on the species richness and total weed density
2.3 杂草组成结构以及各种杂草的相对重要值变化情况
将杂草归为莎草科杂草、禾本科杂草和阔叶杂草3类进行统计,综合2011~2013年终杂草群落的组成结构可以看出,2种除草剂处理以及不除草CK处理的杂草群落组成结构都发生了较明显的变化,其共同点是禾本科杂草比例均增加,莎草科杂草比例均减少。但从变化幅度上看,以使用常规选择性除草剂丙·苄的处理变化幅度最大,禾本科杂草已占到总杂草比例的96.8%,而莎草科杂草几近绝迹(图4)。结合终杂草总密度可以看出,CK组终杂草密度有缓慢下降趋势;草铵膦处理组终杂草密度变得很低;而丙·苄处理组却呈现增长的趋势,且增长的动力主要来自于禾本科杂草的显著增加(图4)。该结果表明,相对于灭生性除草剂草铵膦,选择性除草剂由于对禾本科杂草防除效果较差而使其占总杂草比例显著增加,成为绝对的优势杂草。
单一使用丙·苄处理使各种杂草的相对重要值发生了显著变化,除双穗雀稗Paspalumpaspaloides(Michx.) Scribn.的重要值增加、稗Echinochloacrusgalli(L.) Beauv.重要值波动较小外,其他杂草的相对重要值均显著降低,特别是水虱草Fimbristylismiliacea(L.) Vahl从优势种降低到完全消失。主要优势杂草由最初的水花生Alternantheraphiloxeroides(Mart.) Griseb.+水虱草+双穗雀稗+稗变成双穗雀稗+稗,说明双穗雀稗已成为最主要的优势杂草(图5)。长期单一使用草铵膦处理最明显的变化是双穗雀稗的优势度有所上升,水虱草优势度显著下降,主要优势杂草从最初的水虱草+异型莎草CyperusdifformisL.+稗+水花生变成了异型莎草+双穗雀稗+稗+水花生(图5)。CK处理组显示出莎草科杂草优势度下降和禾本科杂草优势度上升的趋势。主要优势杂草从最初的水虱草+异型莎草+稗+水花生变为异型莎草+碎米莎草CyperusiriaL.+双穗雀稗+稗(图5)。
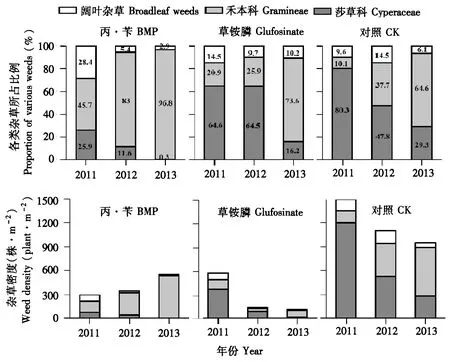
图4 不同除草剂处理对杂草组成结构的影响Fig.4 Effect of different herbicide treatments on weed species community and density
2.4 杂草群落物种多样性的变化
如图6所示,常规选择性除草剂丙·苄处理连续3年Shannon-Wiener、Simpson多样性指数以及Margalef丰富度指数、Pielou均匀度指数均显示出下降趋势,说明物种多样性下降并且出现明显的优势物种。灭生性除草剂草铵膦处理和CK处理的各项指数相对稳定,其原因在于草铵膦的灭生性特点,在没有抗/耐药性杂草出现之前,杂草群落不易形成优势物种。
2.5 杂草群落相似性的变化
2种除草剂处理以及CK都表明,随着连续处理年份的增加杂草群落相似性降低,趋异性(距离)增大(表1)。通过Bray-Curtis距离系数矩阵绘制的聚类图,显示3年的CK组杂草群落最为相似聚在一起,表明杂草群落结构的变化较小。2012和2013年丙·苄处理和草铵膦处理分别聚在一起,2011年的丙·苄与草铵膦处理聚在一起(图7)。这说明2种除草剂在使用初期,由于对杂草的防除作用使杂草密度减小,导致杂草群落较为相似。但随着各自使用时间的增加,显现出不同的除草机理,从而对杂草群落产生不同影响,导致异化。
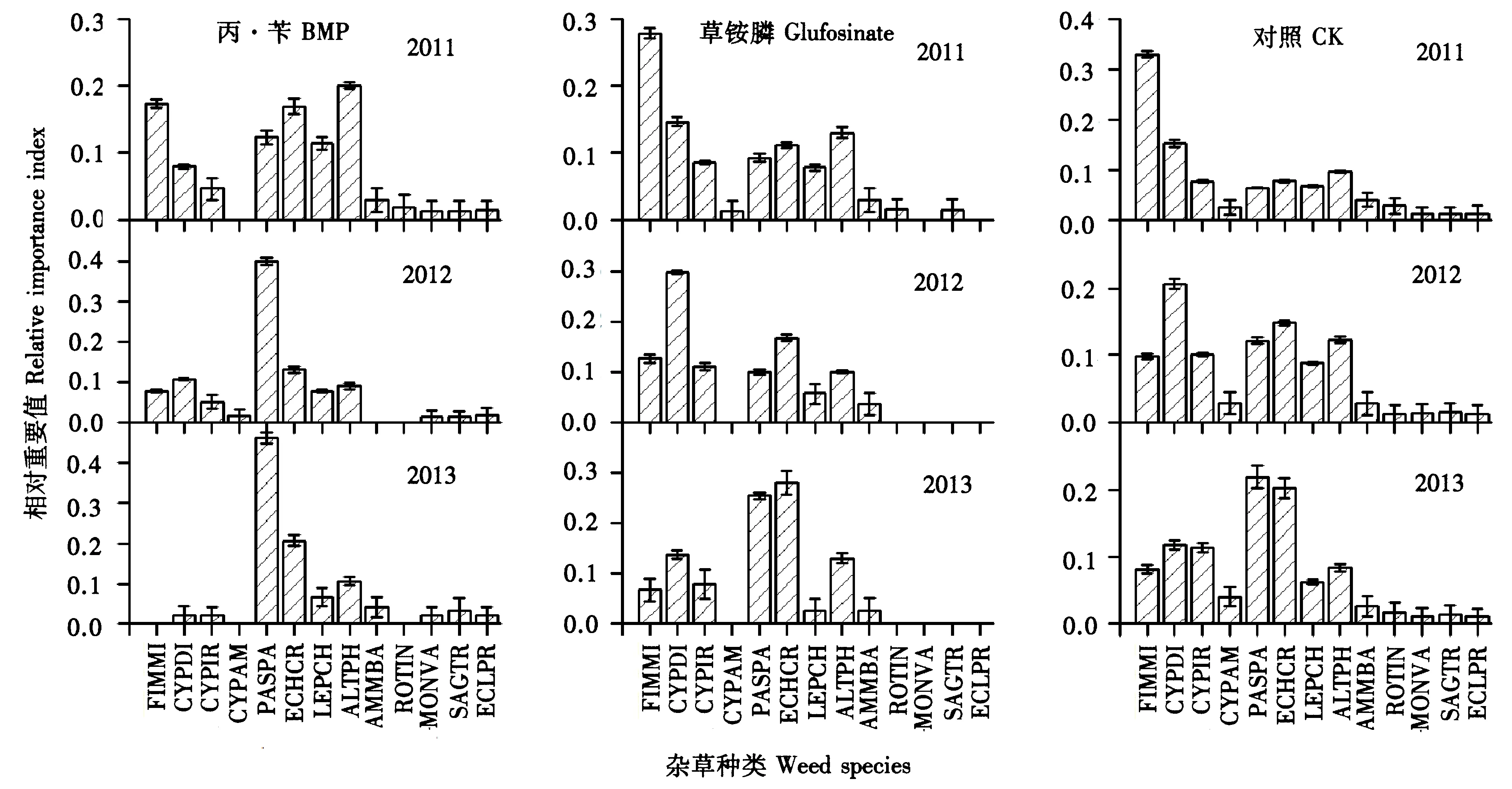
图5 各处理在2011~2013年各种杂草相对重要值的比较Fig.5 Relative importance of different weed species under the 3 different herbicide treatments from 2011~2013 FIMMI, 水虱草; CYPDI, 异型莎; CYPIR, 碎米莎; CYPAM, 阿穆尔莎草; PASPA, 双穗雀稗; ECHCR, 稗; LEPCH, 千金; ALTPH, 水花生; AMMBA, 水苋菜; ROTIN, 节节菜; MONVA, 鸭舌草; SAGTR, 野慈姑; ECLPR, 鳢肠。
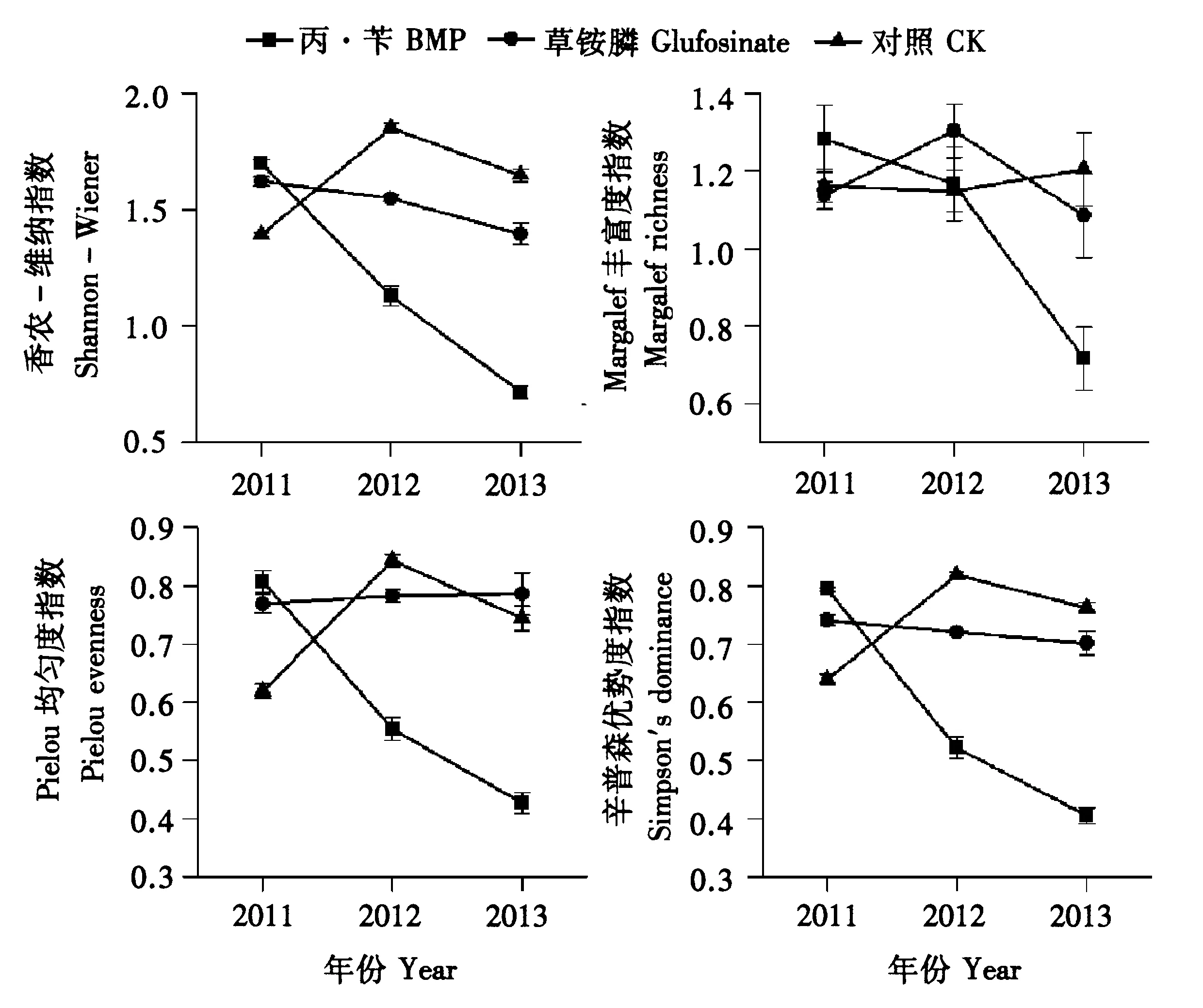
图6 长期使用单一除草剂对杂草物种多样性影响Fig.6 Effect of single herbicide for long-term usage on species diversity indices
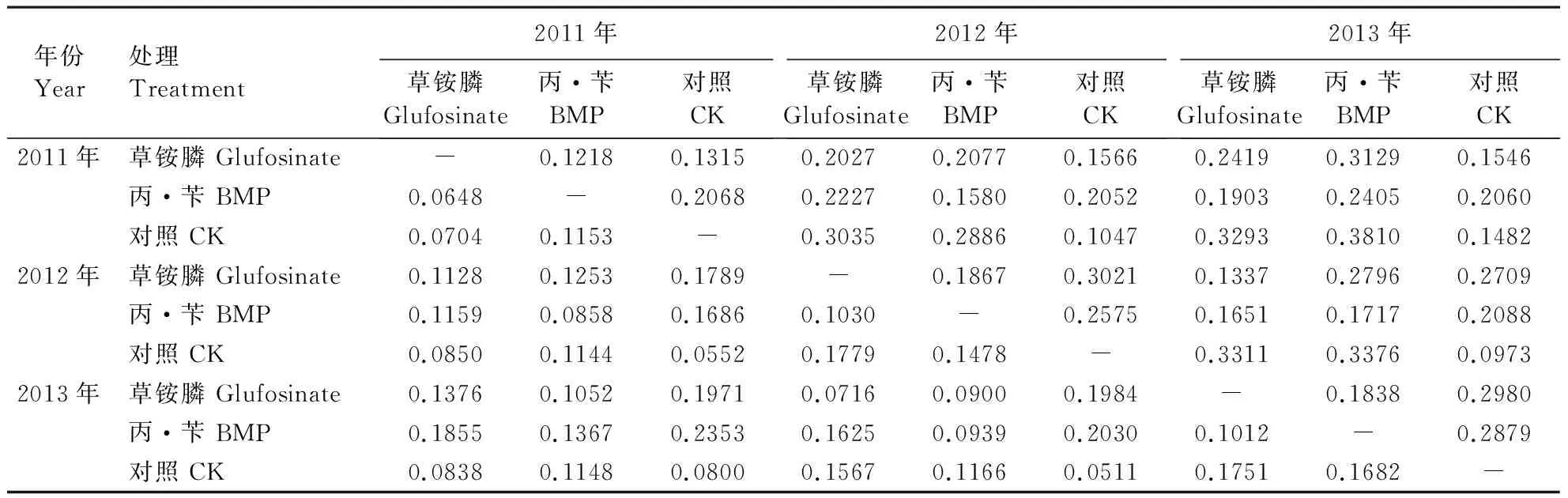
年份Year处理Treatment2011年草铵膦Glufosinate丙·苄BMP对照CK2012年草铵膦Glufosinate丙·苄BMP对照CK2013年草铵膦Glufosinate丙·苄BMP对照CK2011年草铵膦 Glufosinate-0.12180.13150.20270.20770.15660.24190.31290.1546丙·苄 BMP0.0648-0.20680.22270.15800.20520.19030.24050.2060对照 CK0.07040.1153-0.30350.28860.10470.32930.38100.14822012年草铵膦 Glufosinate0.11280.12530.1789-0.18670.30210.13370.27960.2709丙·苄 BMP0.11590.08580.16860.1030-0.25750.16510.17170.2088对照 CK0.08500.11440.05520.17790.1478-0.33110.33760.09732013年草铵膦 Glufosinate0.13760.10520.19710.07160.09000.1984-0.18380.2980丙·苄 BMP0.18550.13670.23530.16250.09390.20300.1012-0.2879对照 CK0.08380.11480.08000.15670.11660.05110.17510.1682-
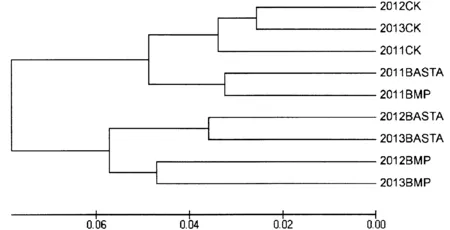
图7 不同除草剂处理2011~2013年杂草群落相似性聚类图Fig.7 Cluster analysis of weed community similarities under different herbicide treatments from 2011 to 2013
3 讨论
3.1 草铵膦和丙·苄的防除杂草效果
丙草胺(pretilachlor)属酰胺类除草剂,常用作苗前处理;苄嘧磺隆(bensulfuron methyl)属磺酰脲类除草剂,既可用作苗前处理剂亦可用作杂草苗后早期使用(强胜,2009)。两者的复配适合土壤封闭处理,使用推荐剂量有效浓度236.25 g·hm-2。在本试验水直播稻田中,用药20 d内能有效抑制莎草科和禾本科杂草萌发,且在2011年试验中对杂草幼苗有良好药效。但是随着药效减弱,后期仍有杂草发生,特别是多年生杂草双穗雀稗在用药后期会发生大量匍匐茎。2011~2013年连续单一使用丙·苄,在用药初期具有良好的抑草效果,但随着双穗雀稗成为绝对优势杂草,用药后期杂草危害情况呈逐年加重趋势,终总杂草密度逐年升高。
草铵膦作为灭生性苗后除草剂,需要在杂草出草高峰期过后用药,以达到将各类杂草一次性杀灭的效果。通过对草铵膦在水稻田的合理用药时间的研究,在水直播稻田5~6叶期,杂草发生量大且生物量较小时用药,可达到好的效果,而用药过早仍会在后期发生大量杂草(孙光辉等,2014)。使用推荐剂量有效浓度450 g·hm-2,对各类杂草均能达到良好的除草效果。但由于用药时间在出草高峰后的水直播后40 d,某些禾本科杂草已经具有较大生物量,而草铵膦对这些杂草的药效较差。连续3年单一使用草铵膦,终总杂草密度呈下降趋势,说明在没有出现抗/耐药性杂草的情况下,长期使用草铵膦可发挥稳定药效,且由于其杀草谱广可避免出现优势杂草。综合3年的水稻产量结果可以看出,种植抗除草剂转基因水稻田,即使在仅使用一次除草剂的情况下,只要选择适当的用药时期,草铵膦处理小区便可保持较高的产量。而使用丙·苄处理的小区仅在试验首年与使用草铵膦小区产量无显著性差异,在随后2年,其小区产量显著低于草铵膦处理的小区。显然,在常规水稻种植中使用常规除草剂丙·苄,一次化除不能达到有效控草的目的,需要再施行茎叶处理除草剂一到两次,比较考虑用药和人工施药成本,常规除草剂除草成本大约增加1倍。此外,后期使用的茎叶处理剂还存在对有些恶性杂草不敏感的问题,实际除草效果也将打折扣。因此,种植抗除草剂转基因水稻可以降低除草成本,除草效果在较大程度上得到保障。
3.2 长期使用单一除草剂对杂草群落结构的影响
杂草是农田生态系统中除作物本身外重要的组成部分。除草剂的选择是引起稻田杂草群落变化的重要因素(张斌和董立尧,2009)。酰胺类和磺酰脲类除草剂是在水稻田中使用最多的除草剂类型(强胜和马波,2004),对两种除草剂复配后的长期单一使用能体现稻田杂草群落变化的一般趋势。本研究水直播稻田杂草的组成结构中,单一丙·苄处理组在第3年禾本科杂草已经成为绝对优势的杂草,组成比达到96.8%。这说明长期使用单一选择性除草剂易加快农田杂草种类演替。多年生杂草双穗雀稗的相对重要值在3年的试验中显著上升成为特优势种,而原先的优势种水虱草最后几乎消失,其原因应该是丙·苄对莎草科杂草具有良好的抑草和除草效果,而对于多年生杂草效果较差,加之后期药效的减弱而使之优势度迅速上升。各项物种多样性指数值的下降说明长期单一使用丙·苄后直播稻田杂草群落稳定性降低,是杂草群落对选择性压力的适应;同时新的优势杂草的出现,也为杂草的防除带来新问题。
草铵膦属于灭生性除草剂,具有宽广的杀草谱(苏少泉,2006)。但灭生性除草剂的杀草效果也由于不同种类杂草具有不同的生理结构、敏感程度以及同种杂草龄期的不同而存在差异(Steckeletal.,1997; Tharpetal.,1999)。本研究中,长期单一草铵膦处理组表现出的杂草组成结构中禾本科杂草比例显著提升,但提升比例低于丙·苄处理组。禾本科杂草比例的提升可能与草铵膦用药时期部分禾本科杂草已经拥有较高生物量而敏感度下降有关。从各种杂草相对重要值变化的结果可以看出主要优势杂草的变化较丙·苄处理组要小,异型莎草、稗、水花生仍是主要优势杂草,仅有水虱草被双穗雀稗替代。双穗雀稗优势度的提高原因可能是由于其具有大量的匍匐茎且淹没在水层中而避免了与草铵膦的直接接触。
由于草铵膦和丙·苄都具有良好的除草效果,原本出现频度较低的杂草种类随着用药年限的增长而消失,从而使2种处理的物种丰富度都有所降低。但是草铵膦处理组的各项物种多样性指数却并没有丙·苄处理组下降明显,其原因一方面是由于前者总杂草发生量降低而后者却是上升趋势;另一方面原因是由于试验田初始杂草物种丰富度较低。值得注意的是,不除草CK组的群落结构发生了禾本科杂草比例上升而莎草科杂草比例下降的变化,并且终总杂草密度也下降。原因应该与禾本科杂草本身生态适应性、抗逆性和生长势强有关(强胜,2009)。
灭生性除草剂草铵膦是茎叶处理剂,宜在杂草出草高峰后使用,这将导致水稻前期可能遭受杂草危害,特别是杂草密度较大的田块,情况更严重。因此,在这种情况下可以考虑和常规选择性土壤处理除草剂配合使用,以控制苗期杂草,待后期发生的杂草,再使用灭生性除草剂草铵膦。由于草铵膦的广谱特性,几乎可以灭除大多数主要稻田杂草,效果更佳。
综上所述,相对于单一使用常规选择性除草剂丙·苄,灭生性除草剂在短期内单一使用,并不会迅速导致抗性杂草演化。在抗性杂草演化之前不会因单一灭生性除草剂应用而导致杂草群落迅速朝不良方向演替。但是,究竟这种应用可以持续多长时间,是需要更长时间连续观察研究。
卢宝荣, 傅强, 沈志成. 2008. 我国转基因水稻商品化应用的潜在环境生物安全问题. 生物多样性, 16(5): 426-436.
强胜. 2001. 杂草科学面向生物科学时代的机遇与挑战 (上). 世界农业, (4): 37-39.
强胜. 2009. 杂草学(2版). 北京: 中国农业出版社.
强胜. 2010. 我国杂草学研究现状及其发展策略. 植物保护, 36(4): 1-5.
强胜, 马波. 2004. 综观以化学除草剂为主体的稻田杂草防治技术体系. 杂草科学,( 2): 1-4.
苏少泉. 2006. 草铵膦述评. 农药, 44(12): 529-532.
孙光辉, 强胜, 戴伟民, 宋小玲. 2014. 抗草铵膦转基因水稻‘Y0003’直播田除草剂施用技术研究.植物保护, 40(5): 176-180.
于改莲. 2001. 稻田除草剂的正确施用方法. 农药, 40(12): 43-45.
余柳青, 渠开山, 周勇军, 李迪, 刘小川, 张朝贤, 彭于发. 2005. 抗除草剂转基因水稻对稻田杂草种群的影响. 中国水稻科学, 19(1): 68-73.
张斌, 董立尧. 2009. 水稻田杂草群落演化原因及趋势浅析. 贵州农业科学, 37(2): 58-60.
中华人民共和国国家统计局. 2013. 中国统计年鉴. 北京: 中国统计出版社.
左娇, 郭运玲, 孔华, 周霞, 徐林, 郭安平. 2013. 转基因水稻环境安全评价研究进展. 热带作物学报, 34(12): 2521-2526.
Bray J R and Curtis J T. 1957. An ordination of the upland forest communities of southern Wisconsin.EcologicalMonographs, 27: 325-349.
Cardina J, Herms C P and Doohan D J. 2002. Crop rotation and tillage system effects on weed seedbanks.WeedScience, 50: 448-460.
Gaines T A, Zhang W, Wang D, Bukun B, Chisholm S T. Shaner D L, Nissen S J, Patzoldt W L, Tranel P J, Culpepper A S, Grey T L, Webster T M, Vencill W K, Sammons R D, Jiang J, Preston C, Leach J and Westra P. 2010. Gene amplification confers glyphosate resistance in Amaranthus palmeri.ProceedingsoftheNationalAcademyofSciences, 107: 1029-1034.
Hill M O. 1973. Diversity and evenness: a unifying notation and its consequences.Ecology, 54: 427-432.
James C. 2014.GlobalStatusofCommercializedBiotech/GMCrops.BriefNo. 46. Ithaca, NY: ISAAA.
Maclean J L, Dawe D C, Hardy B and Hettel G P. 2003. 水稻知识大全(3版). 杨仁崔, 汤圣祥,等,译. 福州: 福建科学技术出版社.
Margalef R. 1958.TemporalSuccessionandSpatialHetero-geneityinPhytoplankton. California: University of California Press.
Milner A M, Robertson A L, Monaghan K A, Veal A J and Flory E A. 2008. Colonization and development of an Alaskan stream community over 28 years.FrontiersinEcologyandtheEnvironment, 6: 413-419.
Parish T, Lakhani K H and Sparks T H. 1994. Modelling the relationship between bird population variables and hedgerow and other field margin attributes. I. Species richness of winter, summer and breeding birds.JournalofAppliedEcology, 31, 764-775.
Putman R J and Wratten S D. 1984.PrinciplesofEcology. California: University of California Press.
Rajguru S N, Burgos N R, Shivrain V K and Stewart J M D. 2005. Mutations in the red rice ALS gene associated with resistance to imazethapyr.WeedScience, 53: 567-577.
Shrestha A, Knezeric S Z, Roy R C, Ball-coelho B R and Swanton J. 2002. Effect of tillage, cover crop and crop rotation on the composition of weed flora in asandy soil.WeedResearch, 42: 76-87.
Steckel G J, Hart S E and Wax L M. 1997. Absorption and translocation of glufosinate on four weed species.WeedScience, 45: 378-381.
Tharp B E, Schabenberger O and Kells J J. 1999. Response of annual weed species to glufosinate and glyphosate.WeedTechnology, 13: 542-547.
