三峡水库蓄水前后长江口及其邻近海域浮游植物群落结构的变化及原因分析
2014-08-14贾海波邵君波胡颢琰王益鸣魏娜胡序朋
贾海波,邵君波,胡颢琰,王益鸣,魏娜,胡序朋
(浙江省舟山海洋生态环境监测站,浙江 舟山 316004)
长江是中国第一、世界第二大河,也是注入西太平洋最大的河流。长江三峡水利枢纽工程是中国,也是世界上最大的水利枢纽工程。1994年三峡工程正式开工,2003年6月首次蓄水,2006年5月大坝全线建成,成功蓄水至156m水位。虽然在防洪、发电、通航等方面发挥着巨大效益,但三峡工程的建立也可能对长江口及其邻近海域的生态环境产生负面影响(Milliman,1997;Humborg et al,1997;Gong etal,2006),甚至改变东中国海的生态系统结构 (Wang et al,2008;Xu et al,2009;余立华等,2006)。
浮游植物是海洋主要的初级生产者,处于海洋生态系统食物链的关键环节,是海洋中浮游动物及游泳动物等的基本饵料。浮游植物的分布与海洋环境之间有着十分密切的关系,浮游植物物种分布的变化对环境的变化具有指示作用,同时环境条件的改变也直接或间接地影响到浮游植物的群落结构。
虽然,针对三峡水库截流后长江口及其邻近海域浮游植物的变化已进行了一些研究(Gong etal,2006;赵冉,2009;孙军等,2011a,2011),但由于调查航次不连续、浮游植物采样方法不一致、环境参数缺失等问题,尚缺少全面、系统的分析。本研究根据2002-2006年共13个航次的调查结果,系统比较了三峡蓄水前后长江口及其邻近海域浮游植物群落结构的变化,并结合环境因子调查数据,对浮游植物群落演替原因进行了初步分析,从而为了解长江口及其邻近海域生态系统的演变提供科学的依据。
1 材料和方法
1.1 调查站位与时间
2002年秋季至2006年秋季(每年春、夏、秋3个航次)对长江口及其邻近海域 (29.6°-31.8°N,121°-123°E)进行了 13 个航次的综合调查。其中,2002年秋季、2003年春季代表三峡水库蓄水前;2003年夏季至2006年春季代表三峡水库蓄水过程中;2006年夏季、秋季代表三峡水库蓄水后。调查共设35个站位(图1),调查船为“浙海环监”号。

图1 长江口及其邻近海域调查站位
1.2 采样与分析方法
浮游植物样品分为拖网(定性)和采水(定量),拖网样品采集使用Ⅲ型浮游生物网(网口面积0.1m2,筛绢孔径76μm),距离海底2m处至海面垂直拖网(水深≥27m处均取绳长25m),并用5%甲醛固定保存。采水样品采集使用GOFLOW采水器采集表层水,然后按每升水样加入6~8mL鲁哥氏液固定。调查涉及的环境指标有盐度(SAL)、悬浮物浓度(SS)、硅酸盐(DSi)、活性磷酸盐(DIP)、无机氮(DIN)。样品的采集、预处理、分析及鉴定均按《海洋监测规范》 (GB 17378-2007)和相关标准方法进行。
优势度指数的计算公式为:Y=nifi/N
式中:ni为站位中第i种的个体数,fi为第i种在各站位中出现的频率,N为各站位的总个体数。
1.3 统计分析
应用MAKESENS 1.0,采用曼-肯德尔(Mann-Kendall)检验法对调查海域浮游植物和环境因子数据的变化趋势进行分析(Salmi et al,2002)。曼-肯德尔法是一种非参数方法,其优点是不需要样本遵从一定的正态分布,允许缺失数据的存在,也不受少数异常值的干扰,更适用于类型和顺序变量,计算也比较简便(Gilbert,1987)。
应用SPSS 19.0软件,采用spearman相关系数对浮游植物的种类数、硅藻种类比例、甲藻种类比例、丰度、硅藻丰度比例、甲藻丰度比例与主要环境因子的相关性进行双变量相关分析(Bivariate)。
采用典范对应分析(Canonical Correspondence Analysis,CCA)对长江口及其邻近海域浮游植物群落结构与环境因子的关系进行统计分析。CCA是一种非线性多元直接梯度分析方法。它把对应分析与多元回归结合起来,每一步计算结果都与环境因子进行回归,而详细地研究物种与环境的关系(Ter,1986)。CCA要求两个数据矩阵:一个是物种数据矩阵,一个是环境数据矩阵。物种数据矩阵和环境数据矩阵经过log(x+1)转换后,应用Canoco 4.5软件进行典范对应分析 (Ter et al,1998;Ng etal,2003)。
2 结果与分析
2.1 浮游植物群落结构变化
三峡水库蓄水前后长江口及其邻近海域浮游植物群落结构的变化趋势见图2。环境因子和浮游植物群落变化趋势的Manne-Kendall检验结果见表1。调查海域浮游植物种类数量变化范围在172~263之间,最小值出现在2003年春季,最大值出现在2005年秋季。浮游植物种类数量呈现一定上升的趋势(图2),但Mann-Kendall检验结果未达到显著水平 (Z=1.40,P>0.05)(表 1)。
硅藻种类比例呈现随季节波动中逐渐下降的趋势,并且经Mann-Kendall检验达到了显著水平(Z=-2.01,P<0.05)。硅藻种类比例由2002年秋季的77.09%,逐渐下降至2006年秋季的71.43%。各季节间,春季硅藻种类比例始终高于夏秋两季。
甲藻种类比例则呈现随季节波动中逐渐上升的趋势,经Mann-Kendall检验已达到了显著水平(Z=2.14,P<0.05)。甲藻种类比例由2002年秋季的17.32%,逐渐上升至2006年秋季的22.45%。各季节间,春季甲藻种类比例一般低于夏秋两季。结合各航次资料,可以得出调查海域浮游植物种类数量的上升主要是由于甲藻种类数量的增多,尤其是大洋性与暖水性小个体甲藻种类的增多。
长江口及其邻近海域是赤潮多发区,2003年夏季及2004年春季调查遇到大面积赤潮,导致丰度值异常偏高及硅、甲藻丰度比例的异常。因此,未将2003年夏季及2004年春季的丰度及硅、甲藻丰度比例数据纳入统计之中。
调查海域浮游植物丰度变化范围在1.30×104~2.67×105个/L之间,最小值出现在2004年夏季,最大值出现在2006年夏季,各年、季节之间丰度的波动较大。整体上浮游植物丰度呈现一定上升的趋势(图2),但Mann-Kendall检验结果未达到显著水平 (Z=0.78,P>0.05)(表1)。硅藻占细胞丰度的比例呈现随季节波动中逐渐下降的趋势,甲藻丰度比例则呈现随季节波动的上升趋势,但经Mann-Kendall检验两者的变化趋势均未达到显著水平(表1)。
采用优势度指数(Y),将Y>0.02的种类确定为调查海域的优势种。调查海域浮游植物优势种的变化情况见表2。从表2中可以看出,除2004年春季外(调查期间发生了具齿原甲藻(Prorocentrum dentatum)赤潮),中肋骨条藻(Skeletonema costatum)始终是调查海域的第一优势种。中肋骨条藻是典型的广温广盐广布种,在近岸低盐水及河口海域尤为繁茂,是沿岸流的良好指示种,也是影响调查海域浮游植物数量的关键种。虽然中肋骨条藻的优势度呈现一定的波动,但其优势地位十分明显。
综上所述,虽然硅藻依然是调查海域的第一优势类群,中肋骨条藻始终是第一优势种,但硅藻种类比例已显著下降,硅藻丰度比例呈现随季节波动中逐渐下降的趋势,甲藻种类比例显著上升,甲藻丰度比例随季节波动中呈现一定的上升趋势。因此可以得出,三峡水库蓄水后,长江口及其邻近海域浮游植物群落结构已发生了显著的变化。相同的结论,也在其它的调查研究中得到证实(Gong etal,2006;章飞燕,2009;赵冉,2009)。
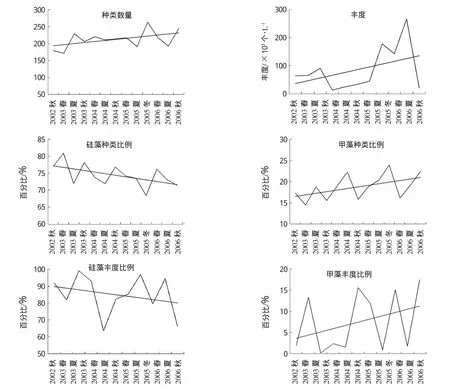
图2 长江口及其邻近海域浮游植物群落结构变化趋势
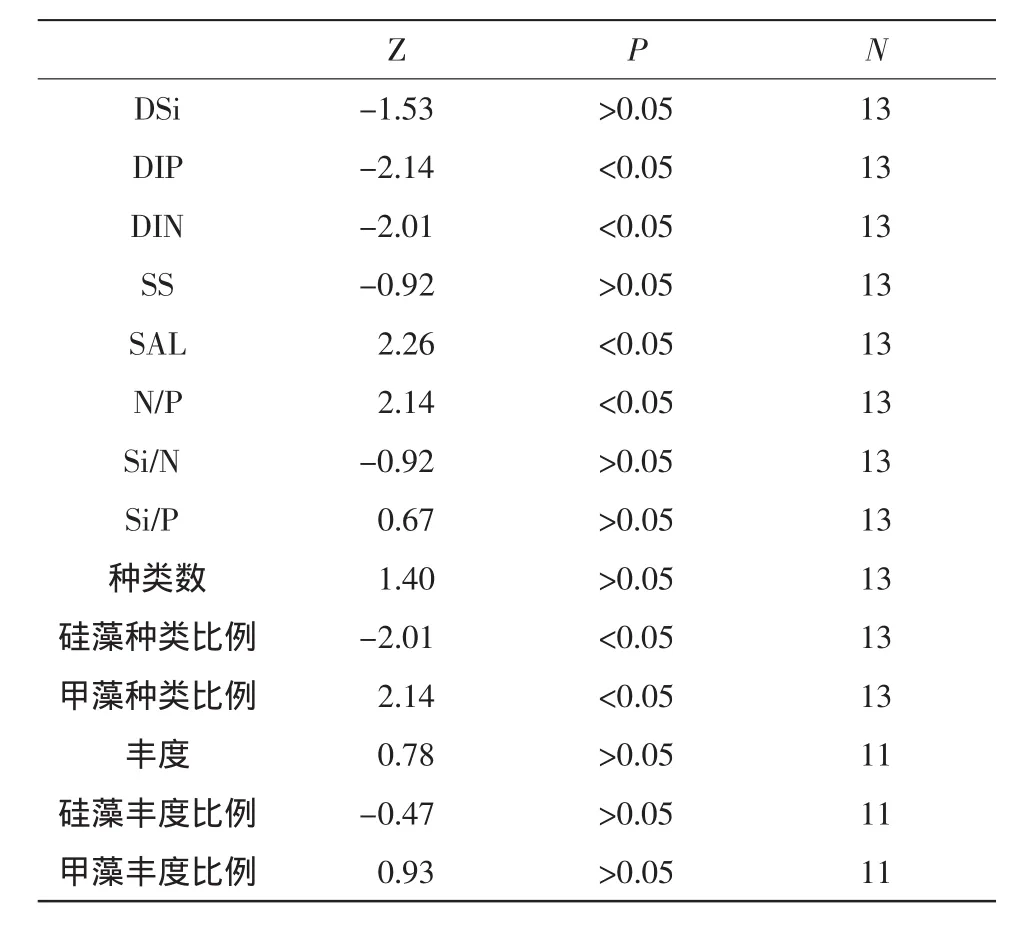
表1 环境因子和浮游植物群落变化趋势的M anne-Kendall检验结果(Z检验统计量,P显著性,n统计样本数)
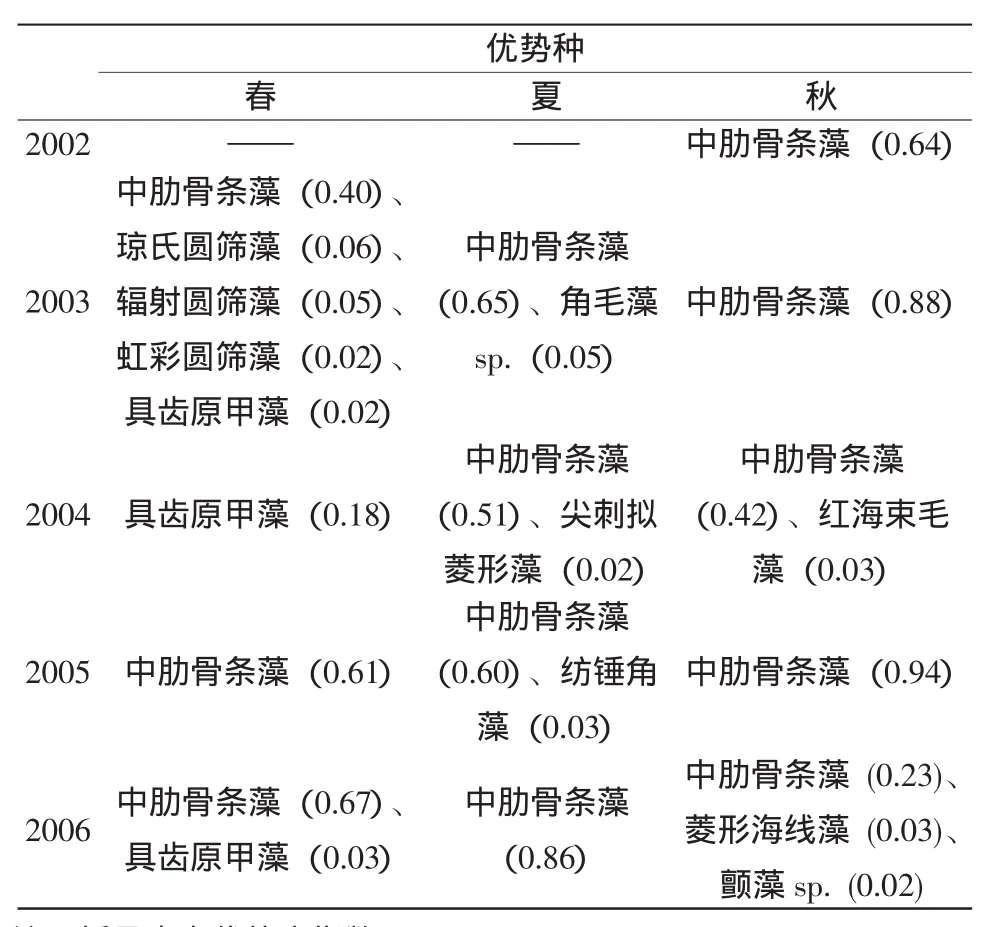
表2 长江口及其邻近海域浮游植物优势种的变化
2.2 环境因子的变化
2.2.1 营养盐含量及结构的变化
三峡水库蓄水前后长江口及其邻近海域营养盐含量及结构的变化趋势见图3。由图3可知,调查海域从2002年秋季开始,随着三峡水库的蓄水,无机氮 (DIN)、活性磷酸盐 (DIP)、硅酸盐(DS)i浓度均呈现下降的趋势,经Mann-Kendall检验,DIN(Z=-2.01,P<0.05)、DIP(Z=-2.14,P<0.05)浓度的变化趋势达到了显著水平,而DSi(Z=-1.53,P>0.05)未达到显著水平 (表1)。DSi浓度的变化趋势未达到显著水平,这主要是DSi浓度的季节变化所致(由春季至夏季、秋季逐渐升高,至下一年春季又大幅降低),进而影响到Mann-Kendall检验结果。但单一季节的DSi浓度变化均呈现下降的趋势,并且结合调查海域其他相关研究,可以判定三峡水库蓄水前后调查海域的DSi浓度也呈现显著的下降趋势 (Gong et al,2006;余立华等,2006;Caietal,2009)。而且,DSi的下降幅度最为明显,从2002年秋季的83.14μmol/L下降至2006年秋季的42.83μmol/L;DIP从 1.40μmol/L下降至 0.89μmol/L;DIN从56.80μmol/L下降至49.18μmol/L。
营养盐结构的变化趋势中,N/P比值呈现显著的上升趋势(Z=2.14,P<0.05),Si/N比值和Si/P比值的变化趋势不明显,Mann-Kendall检验结果也未达到显著水平。N/P比值从2002年秋季的39.86上升至 2006年秋季的 81.27,增幅达103.9%。特别是夏季和秋季,N/P比值的增大趋势较春季更为明显。
Justic等(1995)提出一个系统地评估每一种营养盐化学计量限制标准:(1)若Si/P>22,N/P>22则磷酸盐为限制因子;(2)若N/P>10,Si/N>1则溶解无机氮为限制因子;(3)若Si/P>10,Si/N>l则溶解无机硅为限制因子。根据此判定标准,长江口及其邻近海域2006秋的N/P比值为81.27、Si/N比值为0.93、Si/P比值为68.18,磷酸盐为调查海域浮游植物生长的主要限制因子。Cai等(2009)认为三峡工程蓄水后,长江口海域潜在的磷限制更为严重。这与本研究结果基本一致。三峡工程蓄水后,长江口海域营养盐浓度及结构的显著变化也在其他相关研究中得到证实(Gong etal,2006;余立华等,2006)。相同的磷限制现象也在尼罗河阿斯旺大坝建成后的东地中海的研究中被发现 (Turley,1999)。
海洋中硅酸盐主要是由河流输送。硅酸盐浓度对硅藻生物量和其优势种群的持续时间起着决定作用,而且在浮游植物群落由硅藻型向甲藻型演替过程中起着十分重要的作用。如果硅藻生长所需的最低限度的Si/N比值达不到,则浮游植物群落产生变化而非硅藻成优势种完全有可能发生(Officeret al,1980)。
2.2.2 悬浮物浓度及盐度的变化
悬浮物浓度和盐度也是影响长江口浮游植物分布的重要因素。三峡水库蓄水前后长江口及其邻近海域悬浮物浓度及盐度的变化趋势见图4。由图4可知,随着三峡水库的蓄水,调查海域悬浮物浓度呈现一定的下降趋势,而且悬浮物浓度的季节间变化幅度较大,但Mann-Kendall检验结果显示悬浮物浓度的变化趋势未达到显著水平(Z=-0.92,P>0.05)。调查海域盐度则呈现出显著的上升趋势(Z=2.26,P<0.05),从2002年秋季的19.44上升至2006年秋季的25.42。各季节间,春、夏季的盐度值高于秋季。
2.3 环境因子对浮游植物群落的影响
2.3.1 相关性分析
采用spearman相关性系数对浮游植物群落与主要环境因子的相关性进行双变量相关分析(Bivariate)。结果表明:DIN与浮游植物群落种类数、硅藻种类比例等各因子均为呈现显著相关性;DIP与硅藻种类比例(r=0.665,P<0.05)呈现显著正相关,与甲藻种类比例(r=-0.725,P<0.01)、甲藻丰度比例(r=-0.609,P<0.05)呈现显著负相关;DSi与甲藻丰度比例 (r=-0.855,P<0.01)呈现显著负相关;N/P比值与硅藻种类比例(r=-0.560,P<0.05)呈现显著负相关,与甲藻种类比例(r=0.593,P<0.05)呈现显著正相关;Si/N比值与甲藻丰度比例(r=-0.755,P<0.01)呈现显著负相关;Si/P比值与甲藻种类比例(r=0.621,P<0.05)呈现显著正相关;悬浮物浓度与硅藻种类比例(r=0.725,P<0.01)呈现显著正相关,与甲藻种类比例(r=-0.632,P<0.05)呈现显著负相关;盐度与甲藻丰度比例(r=0.673,P<0.05)呈现显著正相关(表3)。
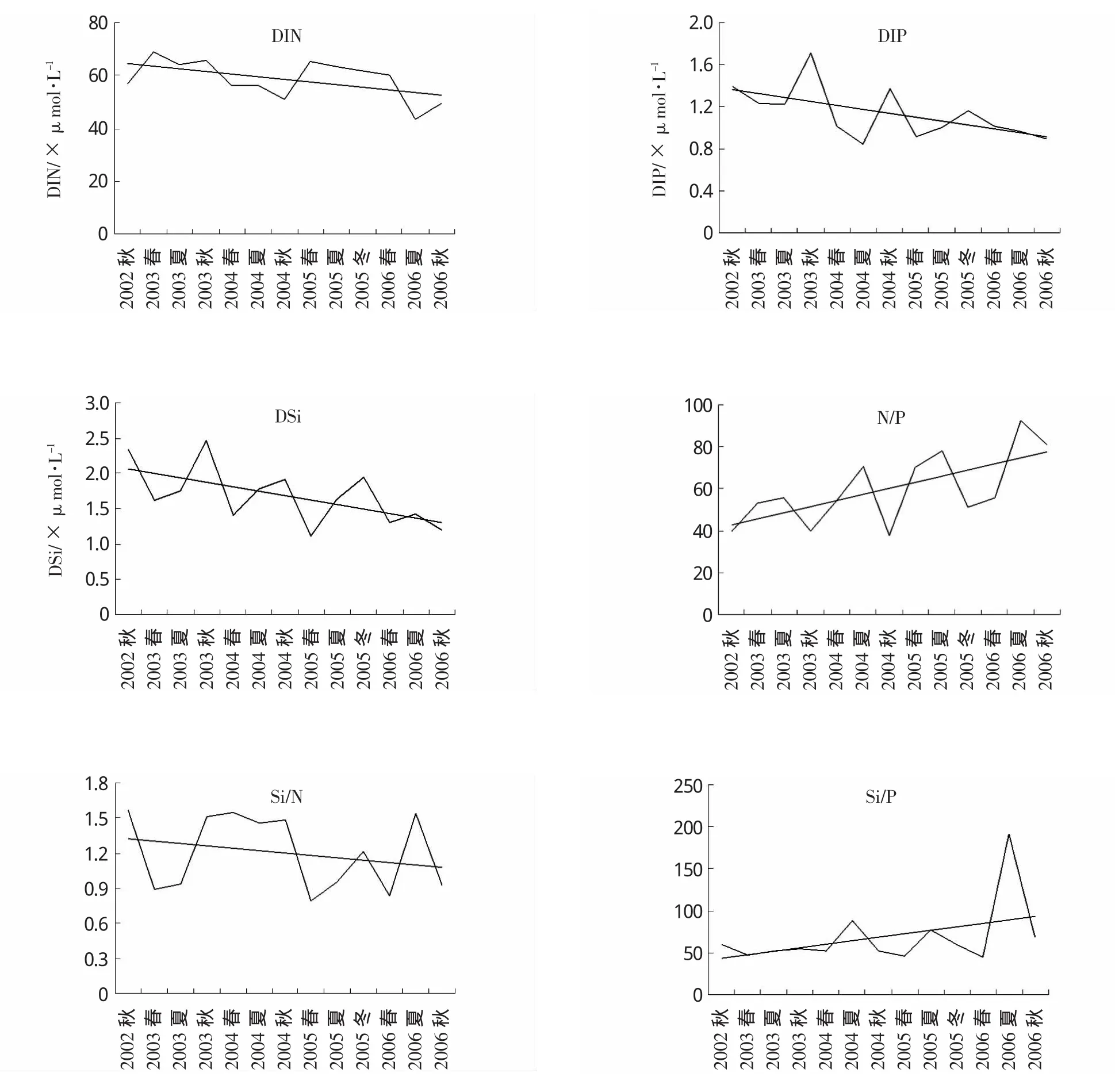
图3 长江口及其邻近海域营养盐含量及结构变化趋势
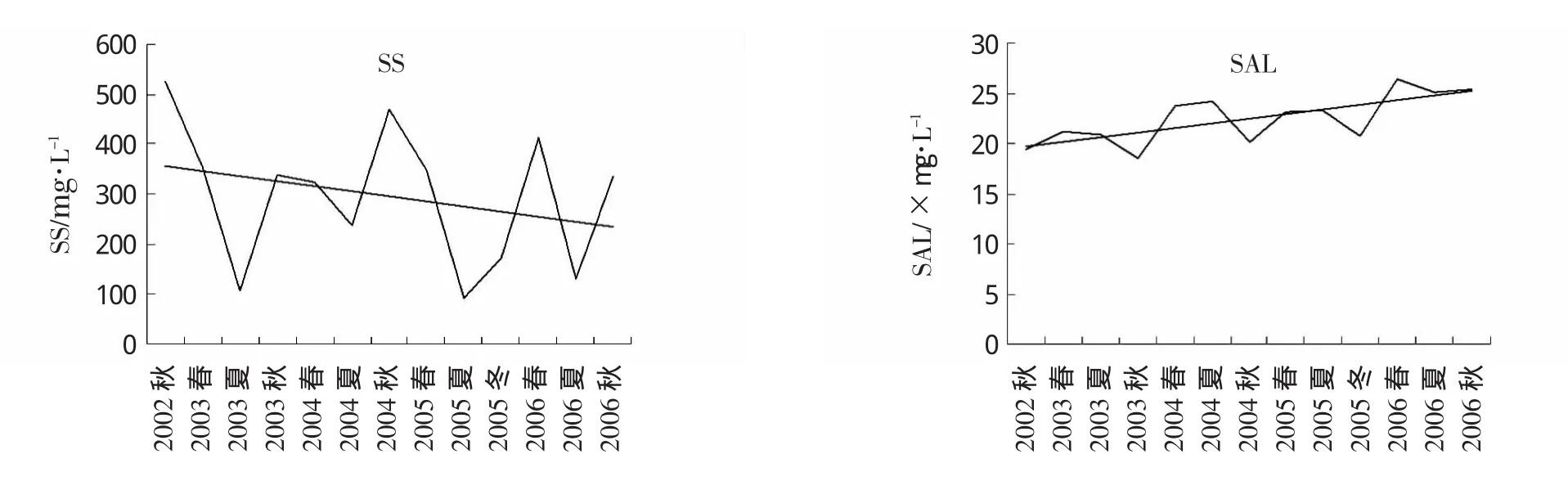
图4 长江口及其邻近海域悬浮物浓度及盐度变化趋势

表3 环境因子与浮游植物群落的spearm an相关性系数(r)
相关性分析表明,三峡蓄水后盐度、N/P比值的升高以及DIP、DSi和悬浮物浓度的降低,均对调查海域浮游植物群落结构变化起了显著的促进作用。
2.3.2 典范对应分析
应用CANOCO4.0软件对长江口及其邻近海域2002年秋季至2006年秋季浮游植物丰度与环境因子数据进行了典范对应分析(CCA),以探讨环境因子对浮游植物主要种类的丰度产生的影响。用于CCA排序的浮游植物种名录见表4。

表4 CCA分析中包含的浮游植物种名录
在CCA排序图中,盐度、悬浮物浓度、DSi、DIP及DIN含量与第一物种排序轴的相关系数分别达到了 0.5250、0.5425、-0.6139、-0.4104和0.072 6;与第一物种排序轴的相关系数分别为-0.468 0、0.6119、0.528 8、0.493 8 和 0.217 2。说明浮游植物物种的分布主要受到盐度、悬浮物浓度、DSi和DIP含量的分布特征和变化趋势的影响,而DIN含量对物种分布的影响相对较小。
调查海域浮游植物物种与环境因子之间的CCA分析发现,甲藻种类主要集中于排序图的右下方(图5),表现出与盐度显著的正相关,与DSi、DIP含量显著的负相关。表明,甲藻物种更适宜生活在较高盐度的海水中,对营养盐的要求较低。而与甲藻相比,硅藻种类分布更偏向于排序图的左上方(图5),呈现出与DSi、DIP含量较强的正相关,与盐度呈显著的负相关。中肋骨条藻(图5中数字1表示)作为调查海域的绝对优势种,在CCA分析图中位于原点附近,受各种环境因子的影响均较小,表现出较强的适应性,在调查海域群落中的优势地位比较稳定。
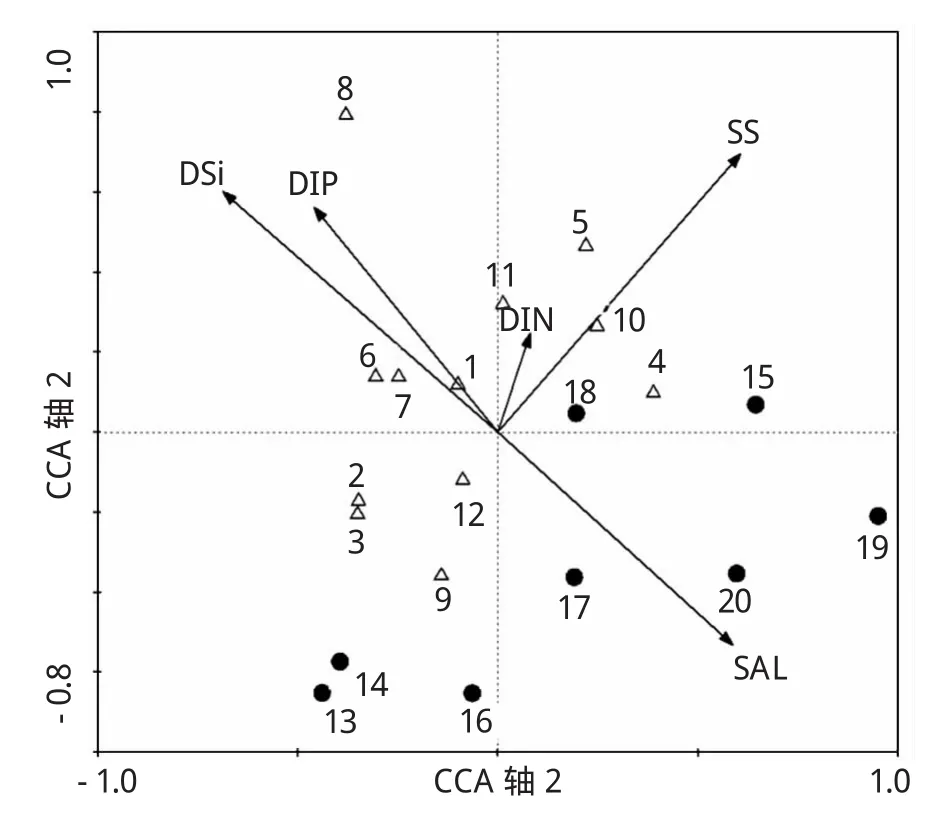
图5 长江口及其邻近海域浮游植物种类与环境因子的CCA分析
将CCA分析与spearman相关性系数统计相比较,两者的分析结果基本一致:调查海域浮游植物群落主要受到盐度、N/P比值、DSi和DIP含量的影响,而受DIN的影响较小。
3 讨论
现场调查和模拟实验均表明,营养盐输入的改变会对浮游植物的细胞丰度和群落结构产生影响(Domingues et al, 2005; Furnas et al, 2005;Paerl,2006)。研究证明,适合硅藻生长的Si、N、P的比例为16∶16∶1(Redfield,1958)。由于较快的生长速度,在高营养盐的海域中硅藻更易成为浮游植物群落的优势类群(Egge,1998)。而硅藻较大的表面积体积比决定了硅藻生长需要较高的营养盐条件(Falkowskietal,1992)。与甲藻相比,硅藻受到P限制的影响更为显著(Egge,1998)。研究表明,当P浓度低于0.3μmol/L时,中肋骨条藻的生长将受到限制(Lietal,2003)。而甲藻赤潮的暴发时则常伴随着高N/P比值和P缺乏的特征(Egge,1998)。其他相关研究也认为,近年来长江口水体N/P比值不断升高,限制硅藻、促进甲藻的生长,这是造成长江口浮游植物种群变化的重要原因(Zhou etal,2008;王江涛 等,2012;李俊龙等,2013)。
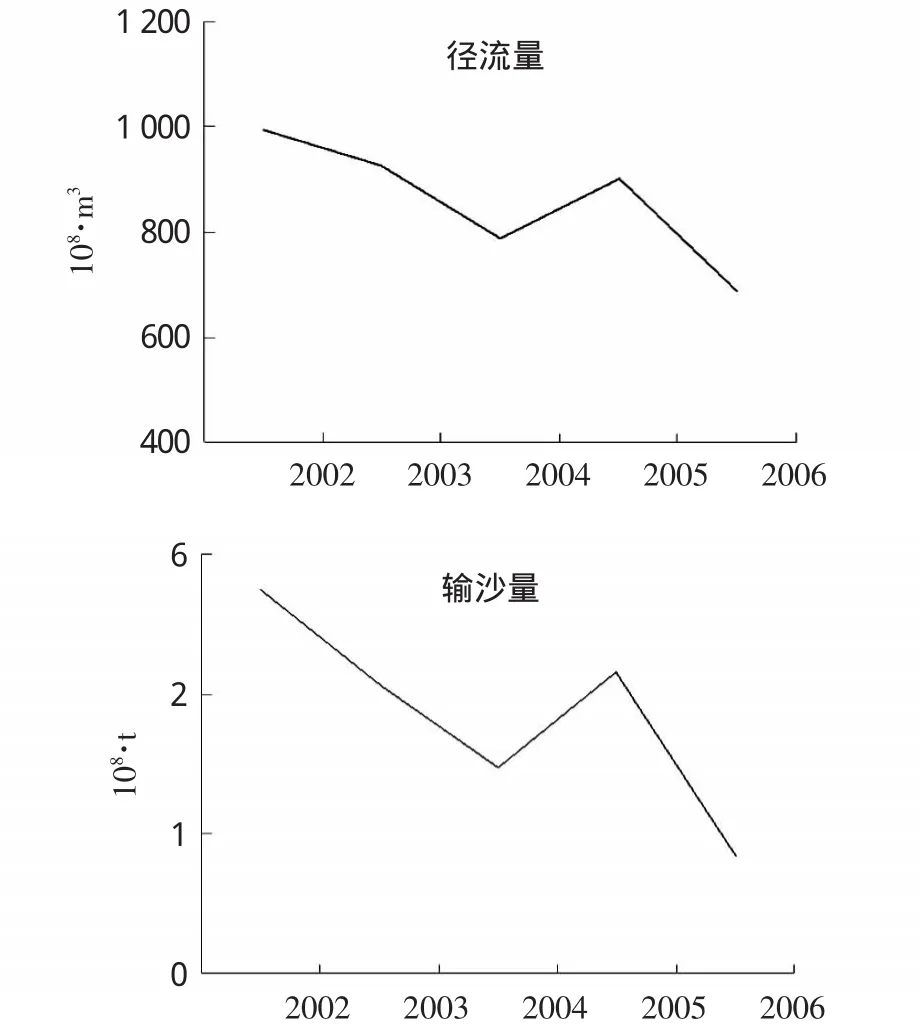
图6 长江大通断面年径流量和输沙量年际统计
长江径流输入是调查海域陆源物质最主要的来源,随径流输入海域的营养物质和泥沙在维持河口生态系统稳定中起着重要作用(余立华等,2006)。由于三峡大坝对长江冲淡水的截流作用,水体流速减慢,加快了泥沙的沉积,使得长江口由冲淡水携带的悬浮物相应减少 (陈慧敏等,2011)。据《长江泥沙公报》统计,与2002年相比,三峡蓄水后2006年长江大通断面年径流量和输沙量分别减少了22.34%和69.16%。长江大通断面年径流量和输沙量变化情况见图6。研究表明,长江口一年中浮游植物的数量与径流量呈正相关的变化趋势,而在年际之间,丰水期浮游植物总量与径流量也呈正相关的趋势(郭玉洁等,1992)。径流量和输沙量的减少以及径流季节分配的改变,一方面造成长江口海域营养盐浓度及结构发生改变,进而对浮游植物群落产生影响。另一方面,三峡蓄水后径流量减小,造成长江口海域盐度升高,大洋性物种与暖水性物种入侵,河口区暖水性物种分布更接近口门,引起浮游植物群落结构发生改变(赵冉,2009)。
甲藻类尤其是有毒有害甲藻的增加,导致海域赤潮发生的风险增加,对海洋生物及人类健康造成危害。同时,由于硅藻主要被中型浮游动物摄食(Harrisetal,2000),硅藻比例的下降,直接导致饵料浮游动物数量的减少,渔业资源也会受到损害。河口的生物种群大多数为季节性洄游种类,其生物量的高低与长江径流量的大小密切相关。径流量减小,冲淡水与海水的交汇区上移,冲淡水的面积变小,水生生物的生活空间将被压缩(姜翠玲等,2003)。刘淑德等(2010)发现三峡水库蓄水后长江口春季鱼类的丰度和群落多样性均显著下降。由于入海径流量的减少而对海域生态系统产生影响,也在尼罗河口、黑海、黄河口、渤海等海域的调查研究中被发现(Wahbyetal,1980;Humborg etal,1997;Ningetal,2010;马媛,2006)。
总之,河流径流通过对河口海域盐度及营养盐结构等的改变,对浮游植物产生影响,进而通过食物链改变浮游生物、鱼类等的群落结构,最终影响到整个海域生态系统。对于三峡工程对长江口及东中国海生态系统的影响仍需要长期的深入研究。
4 结论
(1)三峡水库蓄水后,长江口及其邻近海域浮游植物群落结构发生显著变化。硅藻种类比例显著下降,由2002年秋季的77.09%下降至2006年秋季的71.43%;甲藻种类比例显著上升,由17.32%上升至22.45%。
(2)盐度、N/P比值显著上升,DIP、DSi、DIN浓度显著下降。DIP、DSi、DIN浓度分别从2002 年 秋 季 的 1.40 μmol/L、83.14 μmol/L、56.80μmol/L下降至2006年秋季的0.89μmol/L、42.83 μmol/L、49.18 μmol/L。 而 N/P 比 值 从39.86上升至81.27;盐度则从19.44上升至25.4。
(3)CCA及相关性分析表明,盐度、N/P比值的升高,以及DIP、DSi浓度的降低,均对浮游植物群落结构变化产生了显著的促进作用。
(4)三峡大坝截流蓄水后,长江径流量、输沙量的减少以及径流季节分配的改变是造成长江口海域浮游植物群落结构发生改变的根本原因。
Chai C,Yu Z M,Shen Z L,et al,2009.Nutrient characteristics in the Yangtze River Estuary and the adjacent East China Sea before and after impoundment ofthe Three Gorges Dam.Science ofthe Total Environment,407(16):4687-4695.
Domingues R B,Barbosa A,Galvao H,2005.Nutrients,lightand phytoplankton succession in a temperate estuary(the Guadiana,southwestern Iberia).Estuarine,Coastaland Shelf Science,64(2):249-260.
Egge J K,1998.Are diatoms poor competitors atlow phosphate concentrations.Journalof Marine Systems,16,191-198.
Falkowski P G,Woodhead A D,1992.Primary Production and BiogeochemicalCycles in the sea.Plenum,New-York.
Furnas M,Mitchell A,Skuza M,et al,2005.In the other 90%:phytoplankton responses to enhanced nutrient availability in the Great Barrier ReefLagoon.Marine Pollution Bulletin,51,253-265.
GilbertR O,1987.StatisticalMethods for EnvironmentalPollution Monitoring.Van Nostrand Reinhold Co.,New York.
Gong G C,Chang J,Chiang K P,et al,2006.Reduction of primary production and changing ofnutrient ratio in the East China Sea:Effect of the Three Gorges Dam.Geophysical Research Letters,(33):L07610.
Harris R P,Irigoien X,Head R N,etal,2000.Feeding,growth and reproduction in the genus Calanus.ICES Journalof Marine Science,(57):1708-1726.
Humborg C V,Ittekkot A C,Bodungen B V,1997.Effectof Danube River dam on Black Sea biogeochemistry and ecosystem structure.Nature,(386):385-388.
Justic D,Rabalais N N,Turner R E,et al,1995.Changes in nutrient structure of river-dominated coastal waters:stoichiometric nutrient balance and its consequences.Estuarine,Coastaland Shelf Science,(40):339-356.
Li,R X,Zhu M Y,Wang Z L,etal,2003.Mesocosm experimenton competition between two HAB species in East China Sea.Chinese JournalofApplied Ecology,14(7),1049-1054.
Milliman J,1997.Blessed dams or damned dams.Nature,(386):325-327.
Ng S L,Sin F S,2003.A diatom model for inferring sea level change in the coastal waters of Hong Kong.Journal of Paleoliminology,(30):427-440.
Ning X R,Lin C L,Su J L,etal,2010.Long-term environmentalchanges and the responses ofthe ecosystems in the Bohai Sea during 1960-1996.Deep-Sea Research II,57:1079-1091.
Officer C B,Ryther J H,1980.The possible importance ofsilicon in marine eutrophication.Marine Ecology Progress Series,(3):383-391.
PaerlH W,2006.Assessing and managing nutrient-enhanced eutrophication in estuarine and coastalwaters:interactive effects ofhuman and climatic perturbations.Ecological Engineering,26,40-54.
Redfield A C,1958.The biologicalcontrolofchemicalfactors in the environment.American Science,46:205-221.
Salmi T,Maatta A,Anttila P,et al,2002.Detecting Trends of Annual Values of Atmospheric Pollutants by the Mann-Kendall Test and Sen's Slope Estimates-the Excel Template Application MAKESENS.Finnish Meteorological Institute,Helsinki,Publications on Air Quality.
Ter B C,Smilauer P,1998.CANOCO reference manualand user's guide to Canoco for Windows:software for Canonicalcommunity ordinatio.Microcomputer Power.Ithaca,New York.
Ter B C,1986.Canonical correspondence analysis:a new eigenvector method formultivariate directanalysis.Ecology,67(5):1167-1179.
Turley C M,1999.The changing Mediterranean sea-a sensitive ecosystem.Progress in Oceanography,(44):387-400.
Wahby S D,Bishara N F,1980.The effectofthe River Nile on Mediterranean water,before and after the construction of the High Dam at Aswan.River Inputs to Ocean Systems,311-318.
Wang B D,Brockiviann U,2008.Potentialimpacts of Three Gorges Dam China on the ecosystem of EastChna Sea.Acta Oceanologica Sinica,27(1):67-76.
Xu K H,Milliman J D,2009.Seasonal variations of sediment discharge from the Yangtze River before and after impoundment of the Three Gorges Dam.Geomorphology,(104):276-283.
Zhou M J,Shen Z L,Yu R C,2008.Responses ofa coastalphytoplankton community to increased nutrient input from the Yangtze(Yangtze)River.ContinentalShelfResearch,28:1483-1489.
陈慧敏,孙承兴,仵彦卿,2011.近23 a来长江口及其邻近海域营养盐结构的变化趋势和影响因素分析.海洋环境科学,30(4),551-553.
郭玉洁,杨则禹,1992.长江日区浮游植物的数量变动及生态分析.海洋科学集刊,33,167-188.
国家海洋局,2007.GB 17378-2007海洋监测规范第7部分.北京:中国标准出版社.
姜翠玲,严以新,2003.水利工程对长江河口生态环境的影响.长江流域资源与环境,12(6):547-551.
李俊龙,郑丙辉,刘录三,等,2013.长江口浮游植物群落特征及其与环境的响应关系.环境科学研究,26(4),403-409.
林更铭,杨清良,林维含,等,2012.台湾海峡浮游甲藻的分布特征和变化趋势.海洋通报,14(1),68-79.
刘淑德,线薇微,2010.三峡水库蓄水前后春季长江口鱼类浮游生物群落结构特征.长江科学院院报,27(10):82-87.
马媛,2006.黄河入海径流量变化对河口及邻近海域生态环境影响研究.青岛:中国海洋大学.
孙军,田伟,2011a.2009年春季长江口及其邻近水域浮游植物——物种组成与粒级叶绿素a.应用生态学报,22(1):235-242.
孙军,赵冉,张利永,2011b.春末三峡大坝首次蓄水期长江口浮游植物群集.应用生态学报,22(4):1045-1052.
王江涛,曹婧,2012.长江口海域近50 a来营养盐的变化及其对浮游植物群落演替的影响.海洋环境科学,31(3):310-315.
王金辉,黄秀清,刘阿成,等,2004.长江口及邻近水域的生物多样性变化趋势分析.海洋通报,23(1):32-39.
余立华,李道季,方涛,等,2006.三峡水库蓄水前后长江口水域夏季硅酸盐、溶解无机氮分布及硅氮比值的变化.生态学报,26(9):2817-2826.
章飞燕,2009.长江口及邻近海域浮游植物群落变化的历史对比及其环境因子研究.华东师范大学.
赵冉,2009.三峡大坝截流前后长江口及其邻近水域浮游植物群集对比研究.中国海洋大学.
中华人民共和国水利部,2001-2010.长江泥沙公报2001-2010.北京:水利部长江水利委员会.
