象山港特定海域浮游动物生物量和物种年际及季节变化
2014-08-14杜秀宁王云龙
杜秀宁,王云龙
(中国水产科学研究院东海水产研究所,上海 200090)
象山港是一个半封闭型港湾,处于副热带季风气候区,东北-西南走向,纵深约60 km,平均水深10m,最深约55m。港湾内的鱼虾蟹贝类资源丰富,是我国海产养殖业的重要基地之一。
浮游动物是关系象山港水域生态系统食物产出的关键饵料环节,其重要种群组成和生物量的变动必然反映到渔业资源的现状及发展趋势。有关象山港浮游动物的生态调查最早始于1980年全国海岸带海洋生物资源调查,此后相继在20世纪80年代末、90年代初及21世纪前10年开展了不同尺度的学科综合调查(柏怀萍,1984;徐兆礼等,1992;宁修仁等,2002;王春生等,2003;尤仲杰等,2011),各调查均提供了有价值的基础资料。
2010年象山港被选做聚鱼增殖型海洋牧场研究与示范执行海域,拟放流增殖中国对虾、黄姑鱼等经济物种。港湾内的浮游动物自然地响应牧场建设带来的生态环境改善,食物链上的各营养级的生态性能也能够得以提高,但认为低营养级的浮游动物长期建立的生态平衡体系更可能从根本上决定放流物种在短期内的种群繁殖能力,因而有必要从长期变化角度加深对浮游动物群落特征的了解。本文依据在当前象山港海洋牧场生态监测区的1988及1992年浮游动物的月度海上调查数据(未发表),结合近30年来的不同时期在此开展的其他较大尺度的浮游动物调查资料,探讨了该重要生物营养级的生物量及物种组成的变化。
1 材料和方法
数据资料有东海水产研究所对象山港浮游动物1988年逐月和1992年季度月的调查数据,将结果主要对比了柏怀萍(1984)对1980象山港浮游动物的调查分析、王春生等(2003)对2000年浮游动物4个季度月的调查数据、尤仲杰等(2011)对2007-2009年桡足类各年季度月的分析数据(表1,后文结果中对这些调查资料相关数据的引用为避免过多重复而省略)。
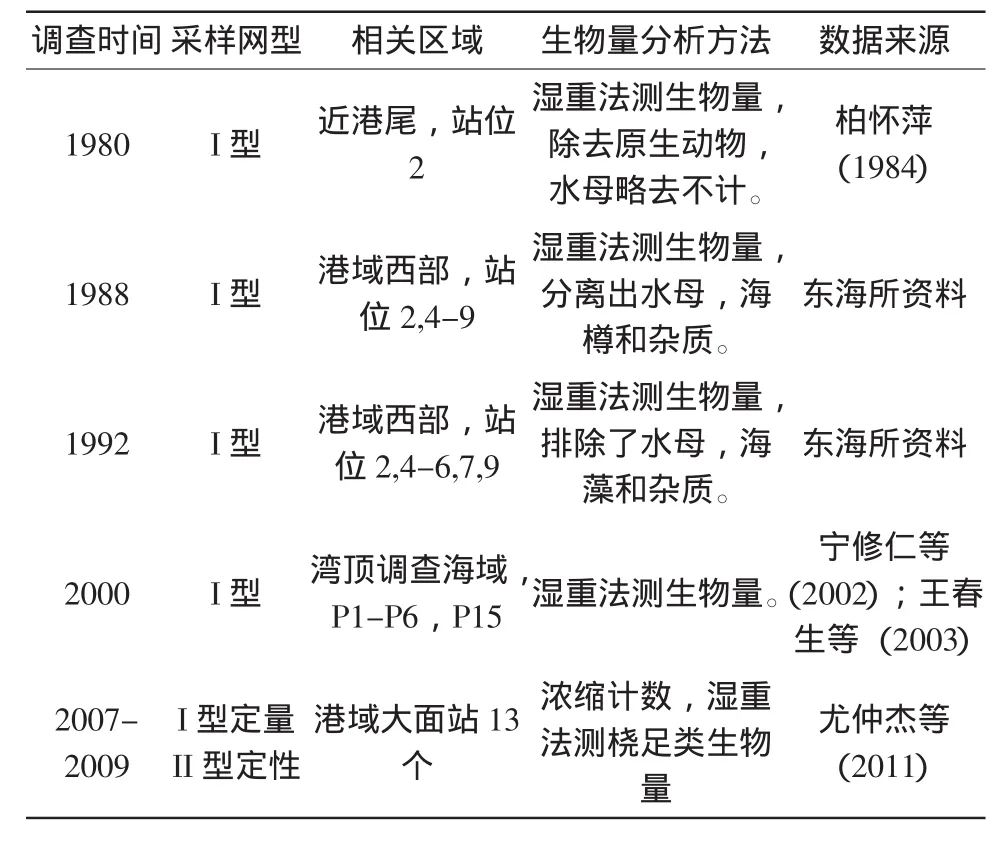
表1 象山港浮游动物相关资料
有关丰度、生物量、优势种及多样性的含义参考王春生等(1999)。
象山港的海洋牧场生态环境监测区(121°32′51.53″-121°37′18.66″E,29°28′50.76″-29°31′14.32″N)如图1,1988年在此范围的相关站位7个,1992年6个。在与其它调查对比时,选择了与监测区相关的数据进行分析。
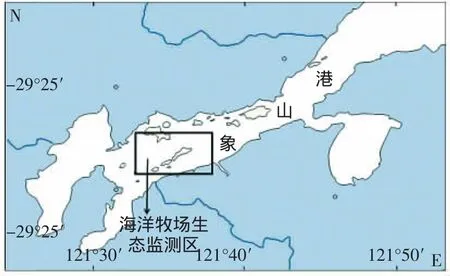
图1 象山港海洋牧场生态环境监测区域图
2 结果
2.1 浮游动物生物量及丰度年际、季节变化
从1980年至2000年,浮游动物年均生物量和丰度呈大致下降趋势(图2),1992年生物量处于低谷,2000年有适度反弹,但与1988年的高值仍差距较大。丰度在20世纪80 s处于稳定的较高水平,但在90 s初急剧下降,在2000年同样有增长迹象。此近20年的分析数据虽然有限,但数据综合得到的变化趋势值得关注,相关结论可在今后调查中进一步取证。
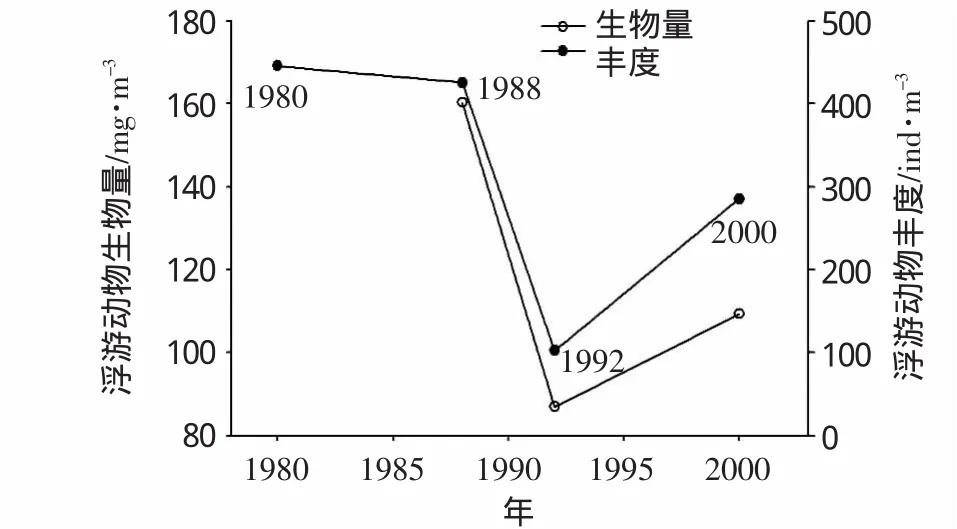
图2 浮游动物生物量及丰度年际变化
浮游动物生物量的季节变化(图3):1988年由高到低的顺序为冬、夏、春、秋,而1992年为春、夏、秋、冬,2000年为夏、春、秋、冬。可见在季节变化模式上,生物量发生了由冬季最低到夏春季跃为最高的转变。
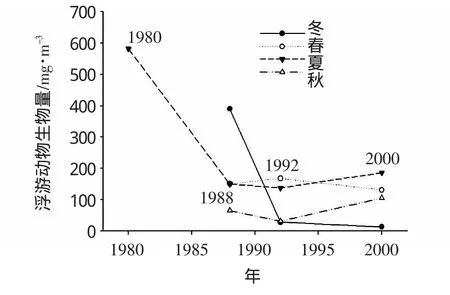
图3 浮游动物生物量季节变化(1980-2000年)
2.2 桡足类生物量及丰度季节变化
比较1988年与2007-2009年间的桡足类生物量及丰度(图4,此为两个不同时间段的对比),2008年各季节的值均较高,尤其是夏季的峰值,生物量达到3 000mg·m-3以上;2007和2009年各季节的生物量值相对较低,但仅冬季值显著低于1988年,其它季节则与1988年相当;1988年冬季的生物量与2008年冬季相当(分别是495.6和571.3mg·m-3)。
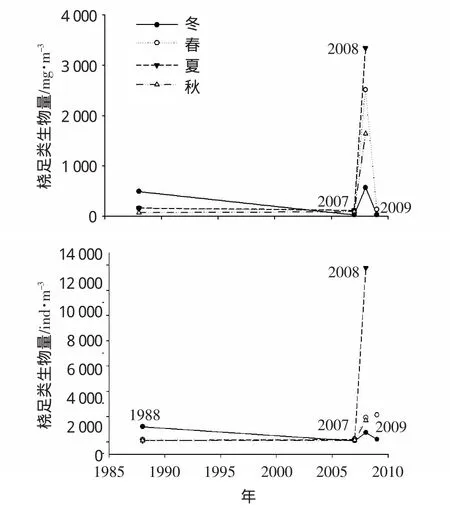
图4 桡足类生物量(上图,mg·m-3)和丰度(下图,ind·m-3)季节变化
不同年的生物量值由高到低的季节变化不同,1988年为冬、夏、春、秋,2007和2008年均为夏、春、秋、冬,此夏季生物量转为最高而冬季降为最低的变化表明桡足类对该海域生态环境变化的显著响应。
在象山港,桡足类对浮游动物总生物量的贡献最大,依据2.1小节对浮游动物总生物量和丰度的变化趋势分析,2000年以后桡足类的生物量和丰度可能也呈现增长变化,正如在2008年观测到了桡足类生物量的显著高值。2009年仅有冬、春季的调查数据,但该年春季桡足类的丰度已高于2008年(分别是2 132.8和1 919.4 ind·m-3)。
2.3 生态类群
自1980年在象山港开展生态调查以来,发现浮游动物群落均以近岸低盐类群为主,其它半咸水性河口、外海高温高盐及广温广盐类群处于非优势地位。近岸低盐类群的物种组成在周年内的季节更替明显,部分物种仅在特定季节形成优势,如墨氏胸刺水蚤 [现多以腹针胸刺水蚤 (Centropages abdominalis)出现在文献资料中。](Centropages mcmurrichi)在冬季,太平洋纺锤水蚤(Acartia pacifica)在夏季,针刺拟哲水蚤 (Paracalanus aculeatu)在秋季;还有物种能跨多个季节形成一定数量,如真刺唇角水蚤(Labidocera euchaeta)和汤氏长足水蚤(Calanopia thompsoni)。
2.4 物种多样性
浮游动物物种数在1992年的各季节均最低(表2),2007-2009年的平均物种数多于1988年。各年夏秋季的物种数高于冬春季,但1988年及2007-2009年的物种数在各季节间的差异显著,1992年的则相对均匀。物种的Pielou均匀度指数在20世纪80 s和90 s间的差异不显著(表3),Shannon-Weiner多样性指数则80 s明显高于90 s,均匀度和多样性的季节特征均为夏秋季高于冬春季。20世纪80 s的多样性 (2.15)和均匀度(0.55)指数的年均值与2007-2009年的对应值接近(分别是1.99和0.54)。
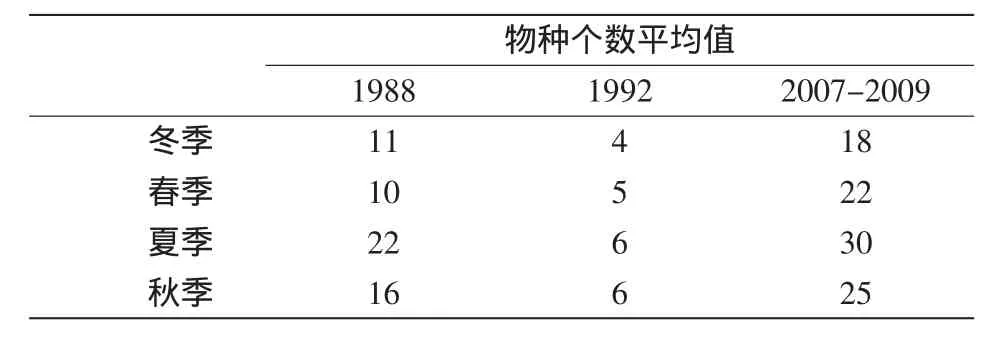
表2 物种数各季节值的年际比较
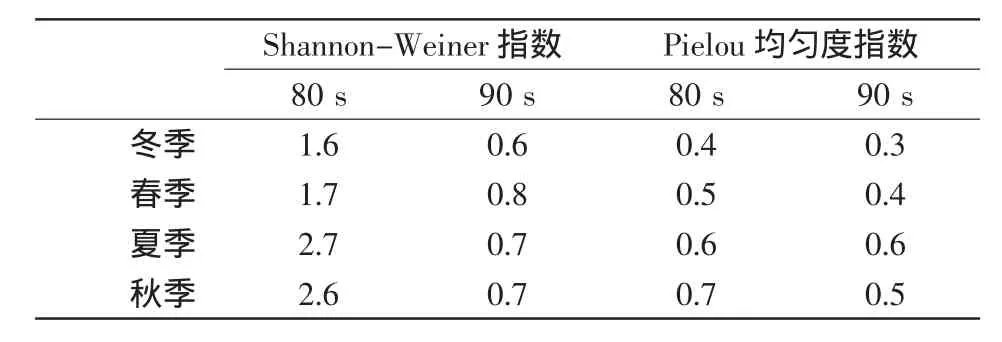
表3 物种多样性各季节值的年际比较
2.5 优势种变化
2000年及此前,主要优势种的年际变化不明显(表4),属于特定生态类型的物种在适宜的季节期间形成数量优势,年际的小波动与横跨多个季节形成优势的物种发生更替有关,如真刺唇角水蚤曾分别是1988年秋季、1992年冬春季的优势种之一。20世纪80 s、90 s及2000年,稳定出现在冬季的优势种有墨氏胸刺水蚤和捷氏歪水蚤(Tortanus derjugini),两者的优势地位可持续到春季。中华哲水蚤(Calanus sinicus)和短尾类蚤状幼体(Brachyura larva)在春季逐渐增多。太平洋纺锤水蚤是夏季的典型优势种,背针胸刺水蚤(Centropages dorsispinatus)仅在2007年夏季为优势种。秋季优势种组成的变异相对较大,除了太平洋纺锤水蚤能延续其在夏季的高值外(1992和2000年),汤氏长足水蚤、针刺拟哲水蚤、真刺唇角水蚤及拿卡箭虫(Sagitta nagae)均曾在不同调查年期间出现。
2000年以后,墨氏胸刺水蚤在2007年冬季和2008年春季的优势地位仍比较突出,但并非群落中的唯一优势种(图5),其丰度比重自1988年以后波动变大。瘦尾胸刺水蚤(Centropages tenuiremis)是出现在2008和2009年冬春季的新优势种,该种曾在1988年(6、12月)和2000年(1、7月)有少量出现。夏季太平洋纺锤水蚤的比重呈下降趋势(图6),与其竞争的物种有:2007年为细巧华哲水蚤(Sinocalanus Tenellus),2008年是长腹剑水蚤属(Oithona sp.)和小拟哲水蚤(Paracalanu parvus)。2008年秋季长腹剑水蚤属秉承了其在夏季的优势,而2007年秋季的优势种为针刺拟哲水蚤。可以看到在此近30年的时间跨度上,优势种组成的多样性及变异性均有增大的变动。
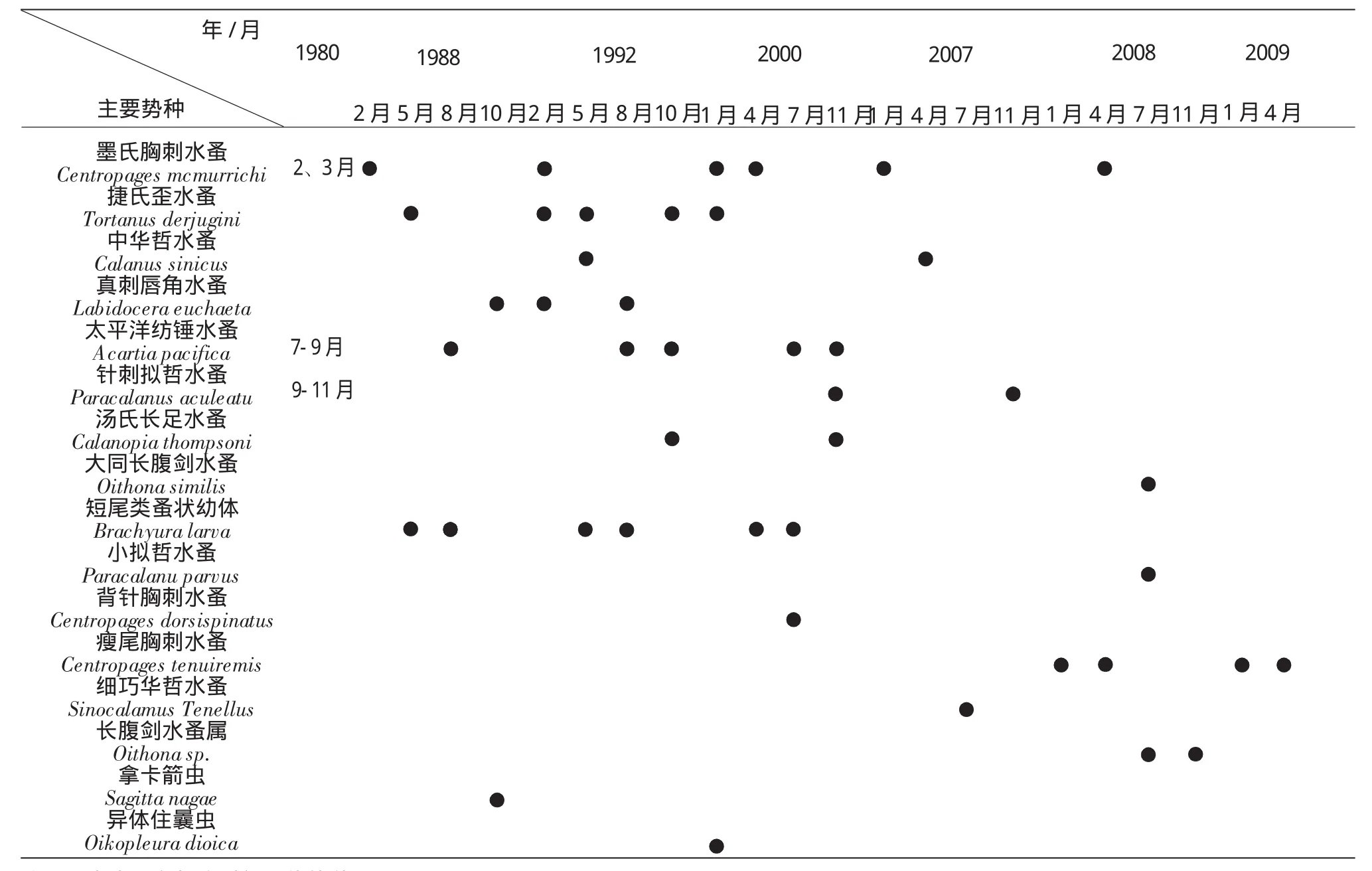
表4 1980-2009年期间浮游动物优势种在各季节组成的年际变化
3 讨论和结论
3.1 浮游植物与浮游动物生物量峰值变化关系
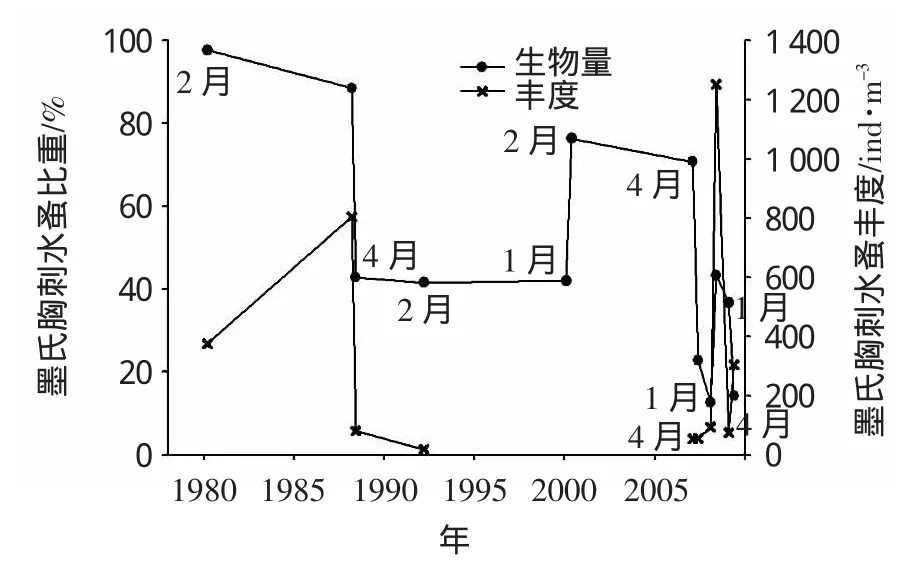
图5 墨氏胸刺水蚤1980-2009年期间冬春季(1、2及4月)丰度及其比重的变化
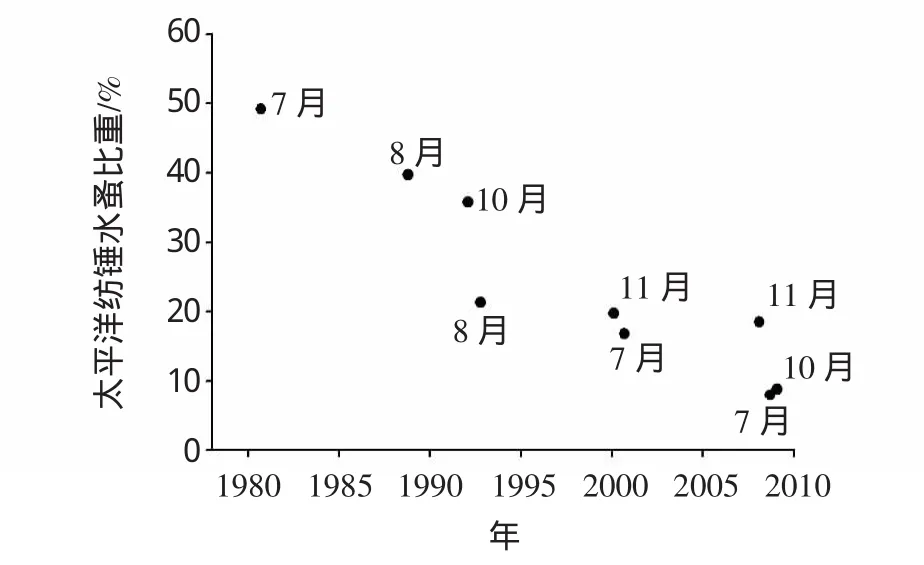
图6 太平洋纺锤水蚤在夏秋季(7、8、10及11月)比重变化
据柏怀萍(1984)1980年的调查,象山港浮游动物生物量的最大值在夏季(5-8月),主要由于近岸低盐类群自5月份以后随水温升高及其他环境因子适宜而大量繁殖的结果,此结论与1988年近岸低盐类群的生物量高峰形成在冬季不同。顾新根等(1993)指出1988年象山港浮游植物的季节变化不同于温带春秋季的双高峰模式,而是特有的秋末冬初最高峰(1月及11、12月)、夏季(7月)次高峰类型,这与同年的浮游动物的季节特征一致,1988年冬季1月的中肋骨条藻(Skeletonema costatum)和 12月 短 角 弯 角 藻 (Eucampia zodiacus)赤潮都对应着桡足类墨氏胸刺水蚤的数量暴发,显然赤潮藻提供了充足的饵料资源(徐兆礼等,1992),两者的匹配变化还如1992年春季(刘子琳 等,1997),王春生等 (2003)也发现2000年的浮游植物和浮游动物的摄食关系是影响彼此现存量变化的重要因子。近几年来象山港浮游植物的季节模式随水文化学等条件的变化而适应性地改变,张海波等(2005)报道了2002年浮游植物的赤潮发生已从1988年的秋末冬初转为春末夏初,2007和2008年浮游植物的丰度是夏秋高、冬春低,这与桡足类生物量的季节顺序一致(尤仲杰等,2011),但2008年浮游植物丰度均低于2007和2009年,这又与同步浮游动物生物量的年际比较结果相反,分析认为象山港的浮游生态系统已显著受海域富营养化等人类社会活动的影响(张健等,2003;张丽旭等,2008;刘俊峰等,2012),随着无机氮磷及有机污染物的严重超标,港湾的浮游植物动态规律变得多样化,生态系统的各营养级平衡关系进而受到干扰,增加了从浮游植物到浮游动物及上层鱼类各营养级间的作用关系及发展趋势的解释和预测难度。
3.2 桡足类优势物种变化探讨
象山港桡足类的小型化趋势明显,大中型桡足类的物种数和个体密度明显低于历史水平(王晓波,2010),表明物种演替随生态环境发生的变化。王扬才等(2011)发现了克氏纺锤水蚤(Acartia clausi)在象山港电厂附近海域成为新优势种的现象。长期以来冬春季的典型优势种墨氏胸刺水蚤的优势地位开始变得不稳定,如2002年12月未有该种记录(刘镇盛等,2004),2008和2009年瘦尾胸刺水蚤削弱了其在群落中的独占优势地位(尤仲杰等,2011),但2010年1月(杜萍等,2011)该种再次在象山港养殖区和放流区占绝对优势(丰度高达570.5~2 780.9 ind·m-3)。研究表明温度是影响墨氏胸刺水蚤种群动态的重要环境因子,在日本近海的Fukuyama港湾的冷水季节期间(11月至次年6月),当水温介于8.9℃~21.1℃时该种大量繁殖,2月有丰度最高值(23 000 ind·m-3)(Liang et al,1996);在邻近日本海的Amursky湾当出现峰值(40 000 ind·m-3)时对应的水温介于15℃~18℃(Kas’yan etal,2004);Irigoien (2004)则报道了其最佳适温范围10℃~15℃;该种也作为北加利福尼亚流系统夏季冷水性的上升流(低于15℃)的桡足类指示种之一(Peterson etal,2003)。Slater(2005)通过实验证明东部亚北极太平洋水域的墨氏胸刺水蚤的低温生存值为4.9℃,温度过高很可能导致该种“异常消失”或低数量。而用温度因素解释墨氏胸刺水蚤在象山港的种群变化也是合理的,2002年12月水温记录为16.2℃,1988冬季有高生物量时的水温介于8.0℃~9.2℃,4月当温度升至16.1℃时,数量急剧下降,5月19.7℃时,该种消失。1992年冬季水温9.3℃,2000年冬季10.3℃~11.7℃,2007-2009冬季平均水温10.2℃,春季15.8℃,2010年1月为9.5-11.6℃,这些冬春季的低温值与该种的同步优势在时间上吻合(表3)。渤、黄海的陆架区生态调查也证实墨氏胸刺水蚤繁盛在低温的冬春季(武文魁,1991;毕洪生等,2001;朱延忠,2008)。
饵料结构的改变也会影响到浮游动物群落的物种更替及种群补充。象山港小粒径的微型(<20μm)和微微型 (<200μm)浮游植物的生物量和生产力在总量中的比重最大,在2000年分别为85%和90%,且有增加的趋势(宁修仁等,2002)赤潮种存在由硅藻向甲藻及原生生物的演变趋势(张海波等,2005)。但有关墨氏胸刺水蚤在自然海域摄食行为的研究尚未见报道,是否由于饵料原因及以怎样的程度影响其种群变动还未详知,可成为今后研究饵料如何调控种群动态机制的切入点。
2008年瘦尾胸刺水蚤数量剧增并超过墨氏胸刺水蚤,尤仲杰等(2011)解释为当年冬季气候变暖,水温较高,导致该暖水种相比适低温性的墨氏胸刺水蚤更有竞争优势。Wu等(2007)从产卵机制上解释厦门港的瘦尾胸刺水蚤极易在冬春季通过迅速提高产卵率以实现其种群扩大,同时充分利用了其它桡足类竞争种处于能导致其数量降低或消失的不利环境时机。近些年象山港热电厂的温排水提高了周边水域温度,也改变了墨氏胸刺水蚤适应的低温生存环境,不利于冬春季的种群扩充,该水域温度的升高也不排除受到全球气候变暖大趋势的影响,而更确凿的解释,需要在综合考虑气候变化和人类活动共同影响的前提下分析物种及生物量的变化机制。
Irigoien X,Huisman J,Harris R P,2004.Global biodiversity patterns of marine phytoplankton and zooplankton.Nature,429:863-867.
Kas'yan V V,2004.Distribution and seasonaldynamics ofthe abundance of Centropages abdominalis Sato and C.tenuiremis Thompson et Scott(Copepoda)in Amursky Bay,Sea of Japan.Russian Journal of Marine Biology,30(2):87-95.
Liang D,Uye S,Onbe T,1996.Population dynamics and production of planktonic copepods in a eutrophic inletofthe Inland of Japan Sea.I.Centropages abdominalis.Marine Biology,124:527-536.
Peterson W T,Keister J E,2003.Interannualvariability in copepod community composition at a coastal station in the northern California Current:a multivariate approach.Deep-Sea Research II,50:2499-2517.
Slater L M,HopcroftR R,2005.Development,growth and egg production of Centropages abdominalis in the eastern subarctic Pacific.Journal ofPlankton Research,27(1):71-78.
Wu L,Wang G,Jiang X,et al,2007.Seasonal reproductive biology of Centropages tenuiremis(Copepoda)in Xiamen waters,People's Republic of China.Journalof Plankton Research,29(5):437-446.
柏怀萍,1984.象山港浮游动物调查报告.海洋渔业,6(6):249-253.
毕洪生,孙松,高尚武,等,2001.渤海浮游动物群落生态特点:II桡足类数量分布及变动.生态学报,21(2):177-185.
杜萍,刘晶晶,徐晓群,等,2011.象山港不同生境冬季浮游动物生态特征比较研究.水产科技情报,38(2):92-99.
顾新根,徐兆礼,1993.象山港中、西部浮游植物生态及赤潮生态研究.海洋渔业,15(2):199-209.
刘俊峰,潘建明,薛斌,等,2012.象山港水域营养状况分析与富营养化水平评价.福建水产,34(1):37-42.
刘镇盛,王春生,杨俊毅,等,2004.象山港冬季浮游动物分布.东海海洋,22(1):34-42.
刘子琳,宁修仁,史君贤,等,1997.象山港对虾增殖放流区浮游植物现存量和初级生产力.海洋学报,19(6):110-115.
宁修仁,胡锡刚,等,2002.象山港养殖生态和网箱养鱼的养殖容量研究与评价.北京:海洋出版社.
王春生,刘镇盛,何德华,2003.象山港浮游动物生物量和丰度的季节变动.水产学报,27(6):595-599.
王春生,杨关铭,何德华,等,1999.秦山核电站邻近水域浮游动物的群落结构和年际变化.东海海洋,17(1):37-47.
王晓波,2010.象山港浮游动物群落结构与时空分布研究.硕士学位论文.杭州:浙江工业大学.
王扬才,吴雄飞,施慧雄,等,2011.象山港电厂附近海域浮游动物群落特征研究.宁波大学学报(理工版),24(3):5-10.
武文魁,1991.墨氏胸刺水蚤在黄海的生态特点.动物学杂志,26(3):1-5.
徐兆礼,顾新根,王云龙,等,1992.象山港赤潮期浮游生物生态特征的分析.海洋通报,11(5):46-53.
尤仲杰,焦海峰,等,2011.象山港生态环境保护与修复技术研究.北京:海洋出版社.
张海波,蔡燕红,项有堂,2005.象山港水域浮游植物与赤潮生物种群动态研究.海洋通报,24(1):93-96.
张健,邬翱宇,施青松,2003.象山港海水养殖及其对环境的影响.东海海洋,24(1):55-59.
张丽旭,蒋晓山,蔡燕红,2008.象山港海水中营养盐分布与富营养化特征分析.海洋环境科学,27(5):489-491.
朱延忠,2008.夏、冬季北黄海大中型浮游动物群落生态学研究.硕士学位论文.青岛:中国海洋大学.
