外源稀土Ce3+对凤眼莲钙相关光合作用的影响
2014-08-07宋雪梅赵仕林四川师范大学化学与材料科学学院四川成都60066农田生态服务能力建设四川省高校工程中心四川成都60066四川大学建筑与环境学院四川成都60065
宋雪梅,夏 超,廖 洋,2*,赵仕林,余 江(.四川师范大学化学与材料科学学院,四川 成都 60066;2.农田生态服务能力建设四川省高校工程中心,四川 成都 60066;.四川大学建筑与环境学院,四川 成都 60065)
外源稀土Ce3+对凤眼莲钙相关光合作用的影响
宋雪梅1,夏 超1,廖 洋1,2*,赵仕林1,余 江3(1.四川师范大学化学与材料科学学院,四川 成都 610066;2.农田生态服务能力建设四川省高校工程中心,四川 成都 610066;3.四川大学建筑与环境学院,四川 成都 610065)
以造成水体污染的主要水生生物之一的凤眼莲为研究对象,探究外源稀土 Ce3+对凤眼莲钙相关光合作用的影响.相比于对照组,营养液中缺钙但添加外源稀土Ce3+的条件下,凤眼莲叶绿素含量减少8.31%、叶绿体荧光产额分别减少6.14%(440nm)和3.82%(480nm)、类囊体膜上Mg2+-ATP酶和Ca2+-ATP酶分别减少1.93%和5.05%,蛋白质含量减少3.94%;但较缺钙的营养液处理组相应指标分别增加了9.89%、15.21%和3.05%、8.08%和10.26%、10.16%.添加Ce3+的营养液处理组的凤眼莲钙相关光合作用指标同样显著提高.结果表明,Ce3+有类似于 Ca2+的特性,适量的 Ce3+可以补偿替代钙,对植物光合作用有促进效果,增加植物生物量,促进凤眼莲型水生植物的生长,对水体富营养化进程的发展起正催化作用.
Ce3+;Ca2+;凤眼莲;光合作用
稀土元素及其化合物因具有独特的光、磁、电子性能,被誉为“工业味精”,不仅广泛应用于轻工、冶金、化工、农业等传统行业,而且在电子、信息、新能源等众多高新技术领域也在逐步开发应用[1].然而,随着稀土应用强度的增加,稀土元素及其化合物大量进入环境,我国淡水域中的稀土元素含量比世界平均水平(0.64μg/L)高出6倍之多[2].据报道,长江上游主要支流及部分干流的丰水期在2007~2009年稀土含量为9.61~63.37μg/L,枯水期稀土含量为 12.11~90.20μg/L,其中以轻稀土离子为主,且稀土元素含量比20世纪80年代丰水期已高出100多倍[3-6].
稀土元素能够在一定浓度范围内能增强植物的光合作用,有效促进植物生物量的增加,影响植物的外部形态和生长发育[7].宋凌云等[8]研究表明稀土元素对水生植物的光合作用有促进效应,可以促进水生植物对N、P元素的吸收,导致水生植物大量繁殖,引起水体富营养化;张贝克等[9]研究表明外源稀土铈对紫背浮萍生长的影响中也具有类似作用.同时,稀土 Ce3+与 Ca2+具有许多共同的性质,如 Ce3+半径与 Ca2+的半径相近[10],因而在生物体内常作为Ca的拮抗剂或发生取代作用,研究Ca对植物的生理功能[11-12]. Hong等[13]研究发现,部分稀土元素可以促进酶的活性.
凤眼莲(Eichhornia crassipes),俗称水葫芦,浮水植物,广泛生长于我国南方水域,具有较强的净化污水能力[14],但大量繁殖的水葫芦覆盖水域表面,造成水体富营养化,破坏水体使用功能.大量研究表明水体中氮磷等营养元素可以促进凤眼莲的生长,进而促进水体富营养化的发生发展
[15],然而稀土元素对凤眼莲生长的影响及其机理的研究甚少.因此,本文以凤眼莲为实验材料,研究稀土元素是否具有类似钙的生理作用,进而阐明稀土元素对长江上游水域水体富营养化的影响机理,为防治水体富营养化提供理论依据.
1 材料与方法
1.1 材料培养
凤眼莲取自成都东郊三环外一水塘.将凤眼莲用清水清洗干净,放入实验室外水池中培养,待用.将凤眼莲横生的匍匐茎割成几段或带根切离几个腋芽,选择带有3~4片嫩叶的大小形态基本相同的凤眼莲,放入1000mL烧杯中进行实验,并对杯身用黑色塑料袋进行遮光处理.
1.2 营养液配置
营养液配制是以改良后Hoagland营养液为基础,并参考其他文献[16-17]确定配方.
1.3 培养方法
Ce3+最适浓度确定的培养,将稀土 Ce3+的浓度梯度设置为以下五个浓度梯度:0、0.6、1.2、1.8、2.4mg/L,每个浓度梯度设置3个平行样.
将烧杯置于 25℃培养箱中,模拟自然光(光照强度3500~4500lx,光照比13:11),培养7d后测定各试样中凤眼莲的叶绿素含量.
培养液分为4个组:(1)改良 Hoagland 营养液对照组;(2)缺 Ca2+的 Hoagland 营养液处理组;(3)添加1.20mg/L Ce3+的Hoagland营养液处理组;(4)添加1.20mg/LCe3+的缺Ca2+的Hoagland营养液处理组.每组设置3个平行样.
凤眼莲培养7d后测定试样中叶绿素、蛋白质、叶绿体荧光光谱、Mg2+-ATP和 Ca2+-ATP酶活性,根据《植物生理学》中的方法测定[18-19].凤眼莲叶片清洗、切片后,采用电子显微镜观察其细胞形态结构.
1.4 数据处理
数据均由均值±标准差表示,统计处理采用软件SPSS 16.0,显著性水平设置为P<0.05.
2 结果分析与讨论
2.1 不同Ce3+浓度的凤眼莲叶绿素含量
如图1所示,叶绿素含量随稀土Ce3+浓度的增加先上升再下降.稀土浓度在 0.6,1.2,1.8mg/L时凤眼莲叶绿素含量均比对照组高,且 1.2mg/L时叶绿素含量达到最大值.稀土浓度为 1.8mg/L时叶绿素含量下降,与对照组相差无几,稀土浓度为2.4mg/L时凤眼莲叶绿素含量降到最低.

图1 不同稀土Ce3+浓度对凤眼莲叶绿素含量影响Fig.1 Amount of chlorophyll in Eichhornia crassipes with different concentration of Ce3+
叶绿素含量的高低可在一定程度上反映出植物光合作用的强弱以及植物的生长情况.实验结果表明适宜浓度(0~1.2mg/L)的稀土元素,可以促进凤眼莲叶绿素含量增加,进而促进凤眼莲生长发育,其中最适稀土浓度为 1.2mg/L;高浓度(2.4mg/L)稀土会抑制凤眼莲的生长,甚至会产生类似重金属的毒性作用.
2.2 叶片电子显微镜图
由图 2可知,叶片中细胞呈半月状且处于张开状态,两两相对而生的保卫细胞围成了气孔,保卫细胞中所含的叶绿体数量比对照组明显增多.
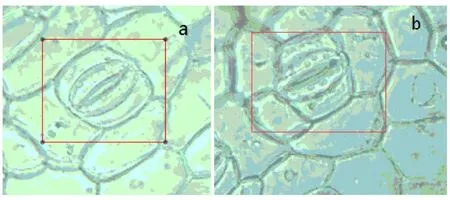
图2 凤眼莲叶片(a)对照组和(b)1.2mg/L Ce3+处理组细胞电子显微镜图(×400)Fig.2 The electron microscope images of Eichhornia crassipes leaf cells (magnification: ×400)
植物叶片内含叶绿体的保卫细胞的气孔作为植物与外界环境进行气体交换的重要通道,在调节植物光合作用中扮演着至关重要的角色
[20-21],其会随着光合作用强度的增大而增大.受控实验表明,添加 Ce3+营养液处理组凤眼莲叶片中叶绿体较对照组明显增多,表明 Ce3+可以促进凤眼莲的光合作用.因此适宜浓度的稀土可以促进凤眼莲的生长,其中 1.2mg/LCe3+是促进凤眼莲生长的最适浓度.
2.3 叶绿素含量
如图3所示,相比于对照组,缺Ca2+处理组凤眼莲叶绿素含量降低了0.19mg/g,添加Ce3+处理组叶绿素含量增加了0.17mg/g.缺Ca2+添加Ce3+处理组的叶绿素含量少于对照组,但比缺 Ca2+处理组增加了 0.1mg/g.统计表明,添加 Ce3+处理组凤眼莲叶绿素含量明显高于对照组(P<0.05),比对照组增加了 18.2%;缺 Ca2+处理组的凤眼莲叶绿素含量明显低于对照组,减少了 16.31%;缺 Ca2+添加 Ce3+处理组的凤眼莲叶绿素含量较对照组变化不明显.
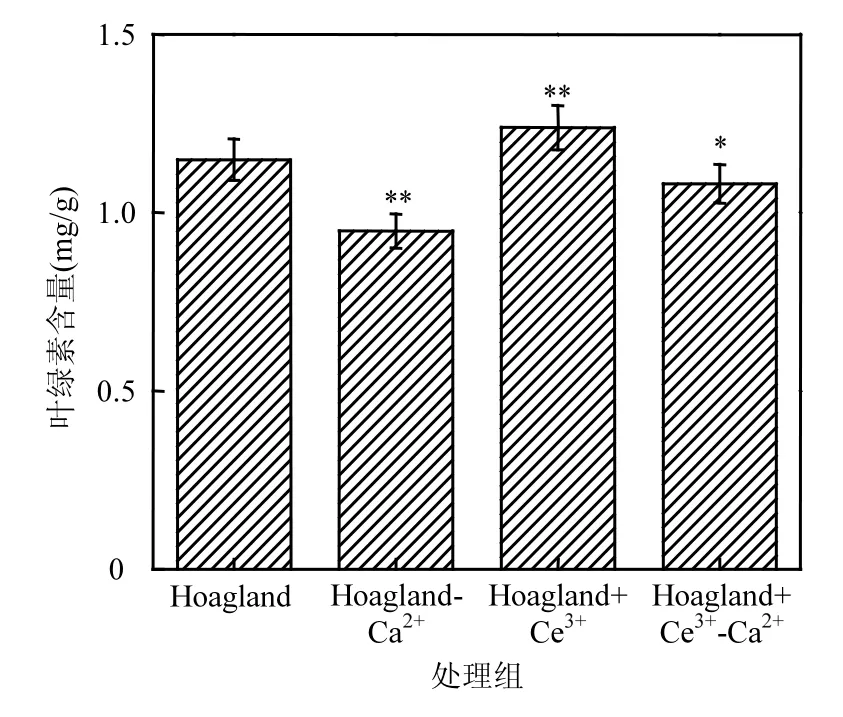
图3 不同培养液中凤眼莲叶绿素含量Fig.3 The amount of chlorophyll of Eichhornia crassipes in different culture solutions
钙元素是植物体生长发育所必需的矿质营养元素,作为叶绿体基本单元的重要组分,Ca2+在光合作用中扮演重要角色[22-25].若植物体内缺少Ca2+则会直接影响植物的光合作用进而影响生长速率[26-27].而植物生长过程中最重要的是光合作用,植物叶片吸收光和热,通过光合作用产生淀粉、脂肪、蛋白质等有机物,实现光能转化为化学能.绿色植物通过叶绿体中的光合作用将光能转换为化学能并将其储存在有机体中.
叶绿素是一类与光合作用有关的最重要色素.添加 Ce3+处理组凤眼莲叶绿素含量的增加表明稀土可以促进植物的光合作用,而在缺 Ca2+条件下的凤眼莲叶绿素含量下降,说明钙对植物叶绿素的合成起主要作用,钙的缺少抑制了叶绿素对光能的吸收.但在缺Ca2+情况下添加了Ce3+,叶绿素含量的部分增加表明 Ce3+可以减弱缺 Ca2+导致的损伤,补偿性代替Ca2+,保持植物进行光合作用的能力.
2.4 叶绿体荧光光谱
由图4可知,在440nm的光激发下,各处理组中凤眼莲叶绿体荧光光谱在680nm附近有一较强发射峰.加Ce3+处理组的荧光峰值增加,比对照组上升了 5.4%,缺 Ca2+处理组峰值最低,比对照组下降了21.35%,缺Ca2+添加Ce3+处理组荧光峰值比缺Ca2+处理组上升了15.21%.
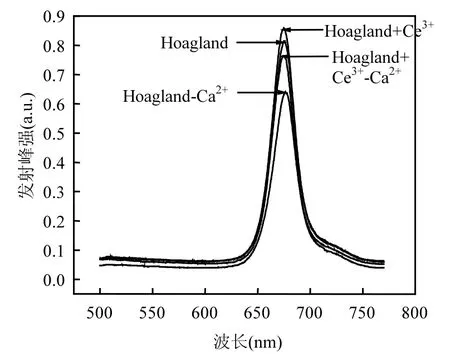
图4 不同培养液中凤眼莲叶绿体荧光(440nm)Fig.4 Chlorophyll fluorescence spectrum (λ=440nm) of Eichhornia crassipes in different culture solutions
由图5可知,在480nm的光激发下,各处理组中凤眼莲叶绿体荧光光谱在660nm附近有一较强发射峰,在680nm附近有一较弱发射峰.加Ce3+处理组的荧光峰值增加,比对照组上升了 9.16%,缺Ca2+处理组峰值最低,比对照组下降了6.87%,缺Ca2+添加Ce3+处理组荧光峰值比缺Ca2+处理组上升了3.05%.
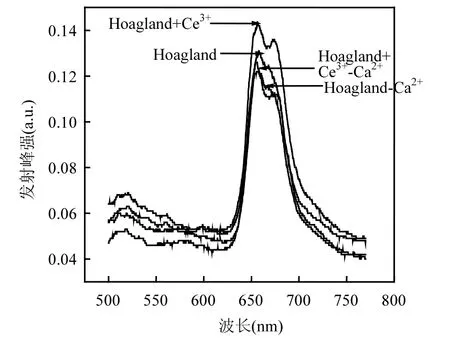
图5 不同培养液中凤眼莲叶绿体荧光(480nm)Fig.5 Chlorophyll fluorescence spectrum (λ=480nm) of Eichhornia crassipes in different culture solutions
叶绿素吸收光能后通过类胡萝卜素和叶绿素a、b传递给中心色素(PSⅠ和PSⅡ),再经过光合磷酸化将电能转化为活跃的化学能.在光系统II(PS II)中含有析氧中心复合体,在此发生原初反应,最初电子供体H2O被氧化并释放氧.该中心结合了特有的Ca2+,在水氧化过程中有重要作用.已知680nm左右的荧光发射峰来自PSII, 440nm和480nm的光激发下,缺Ca2+处理组中凤眼莲叶绿体的PSII荧光发射峰峰强最低,两个光系统的电子流被显著抑制,这对从 PSII 中获得电子有严重影响,因此降低了光能到电能的转化效率,降低了 PSII中心色素叶绿素 a(P680)的荧光产额;而缺Ca2+处理组中添加了Ce3+后,凤眼莲叶绿体的PSII荧光发射峰峰强相比于单独缺Ca2+组要高,表明经过 Ce3+处理后被胡萝卜素和叶绿素 b吸收的光能有效传递到PSII作用中心色素叶绿素a后使其荧光产额提高.这表明Ce3+在析氧中心具有和Ca2+相似的功能,补偿了Ca2+缺失导致的光能转化率,所以Ce3+可以减弱缺Ca2+导致的光能转化率下降的损伤.
2.5 叶绿体Mg2+-ATP酶和Ca2+-ATP酶活性
由图6可知,相比于对照组,缺Ca2+处理组凤眼莲叶绿体的Mg2+-ATP酶和Ca2+-ATP酶活性分别降低了 25.355,54.73μmol/(mg⋅min),加 Ce3+处理组Mg2+-ATP酶和Ca2+-ATP酶活性分别增加了37.89,35.50μmol/(mg⋅min),缺Ca2+添加Ce3+处理组的叶绿体的Mg2+-ATP酶和Ca2+-ATP酶活性比缺 Ca2+处理组分别增加了 20.45, 24.22μmol/(mg⋅min),但低于对照组.统计分析发现,添加Ce3+处理组凤眼莲叶绿体Mg2+-ATP酶和Ca2+-ATP酶活性明显高于对照组(P<0.05),缺Ca2+处理组的叶绿体 Mg2+-ATP酶和 Ca2+-ATP酶活性明显低于对照组;缺Ca2+添加Ce3+处理组的叶绿体Mg2+-ATP酶和Ca2+-ATP酶活性较对照组变化不明显.
植物以三磷酸腺苷(ATP)和NADPH的形式通过非循环和循环光合磷酸化过程,将电能转换为活跃的化学能,产生的化学能被暂时储存在ATP中,这耦合了类囊体膜上的电子传输和光合磷酸化,并最终将二氧化碳和水转化为碳水化合物和氧气.ATP合成酶包括 Mg2+-ATP酶和Ca2+-ATP酶,是整个光合磷酸化的终端.在缺Ca2+处理组中凤眼莲的该两种酶相比于对照组下降较多,表明叶绿体中两个光合磷酸化的效能被抑制,由缺 Ca2+导致的两种光合磷酸化的抑制和叶绿体的耦合水平有密切联系,缺 Ca2+还导致PSII反应中心复合体构造和功能的改变,这对PSII、PSI和Cytb6f化合物间电子传递产生影响并降低了整个光合磷酸化的效率;在加 Ce3+缺Ca2+处理组中凤眼莲两种酶活性得到了一定修复,减轻了因缺Ca2+而引起的损伤.ATP合成酶的减少可能归因于 Ca2+的缺失,特别是类囊体膜上Ca2+-ATP酶含量的减少,而这种情况通过添加Ce3+得到一定程度改善.总之,Ce3+减少了由缺Ca2+导致的光合磷酸化效率的降低,在一定程度上,源于其与Ca2+相似的结构.
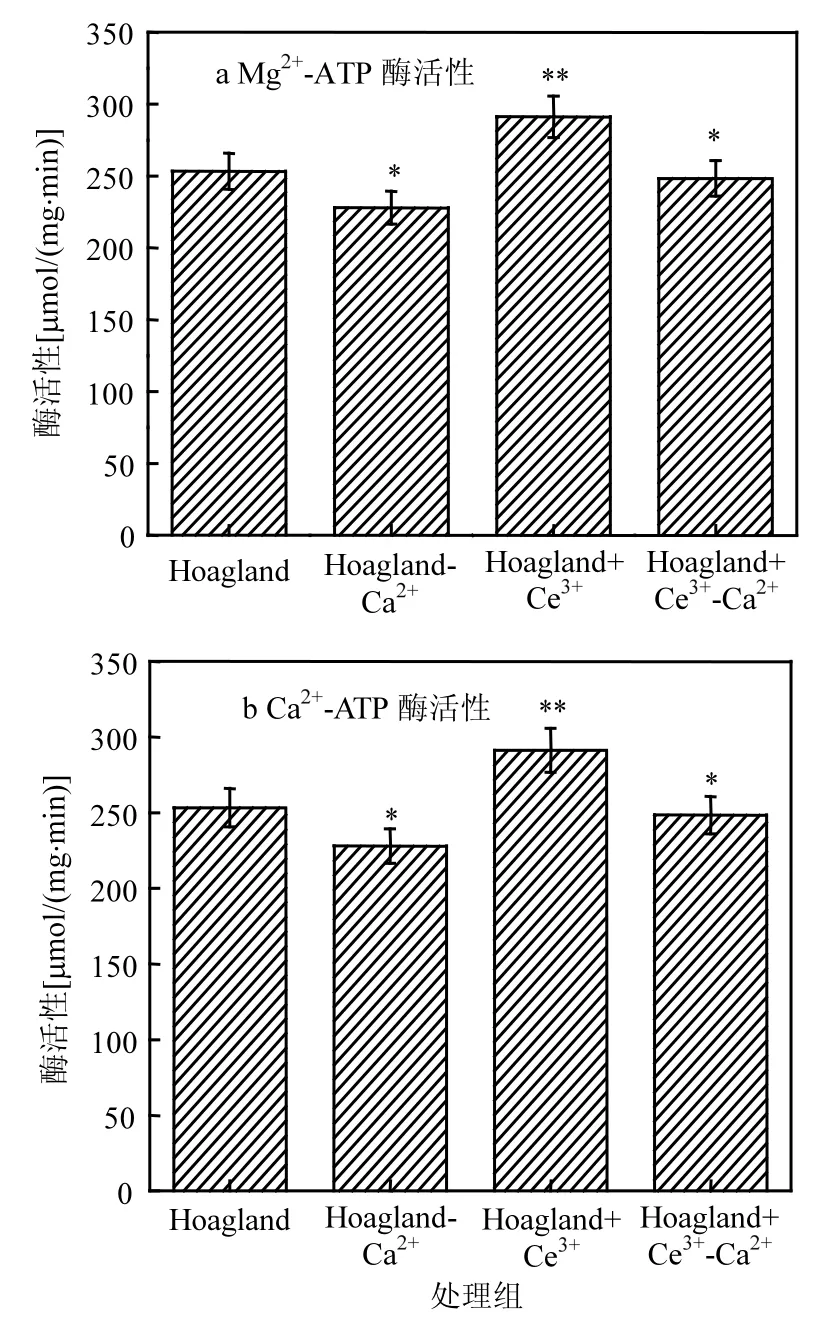
图6 不同培养液中凤眼莲叶绿体Mg2+-ATP酶活性和Ca2+-ATP酶活性Fig.6 the Mg2+-ATP and the Ca2+-ATP activity of Chloroplast of Eichhornia crassipes in different culture solutions
2.6 蛋白质含量
由图7可知,相比于对照组,缺Ca2+处理组凤眼莲的蛋白质含量降低了0.38mg/g,添加Ce3+处理组的蛋白质含量增加了0.51mg/g,缺Ca2+添加Ce3+处理组的蛋白质含量比缺 Ca2+处理组增加了 0.27mg/g,但低于对照组.统计发现,添加 Ce3+处理组凤眼莲蛋白质含量显著高于对照组(P<0.05);缺 Ca2+处理组的蛋白质含量显著低于对照组;缺Ca2+添加Ce3+处理组的蛋白质含量较对照组变化不明显.
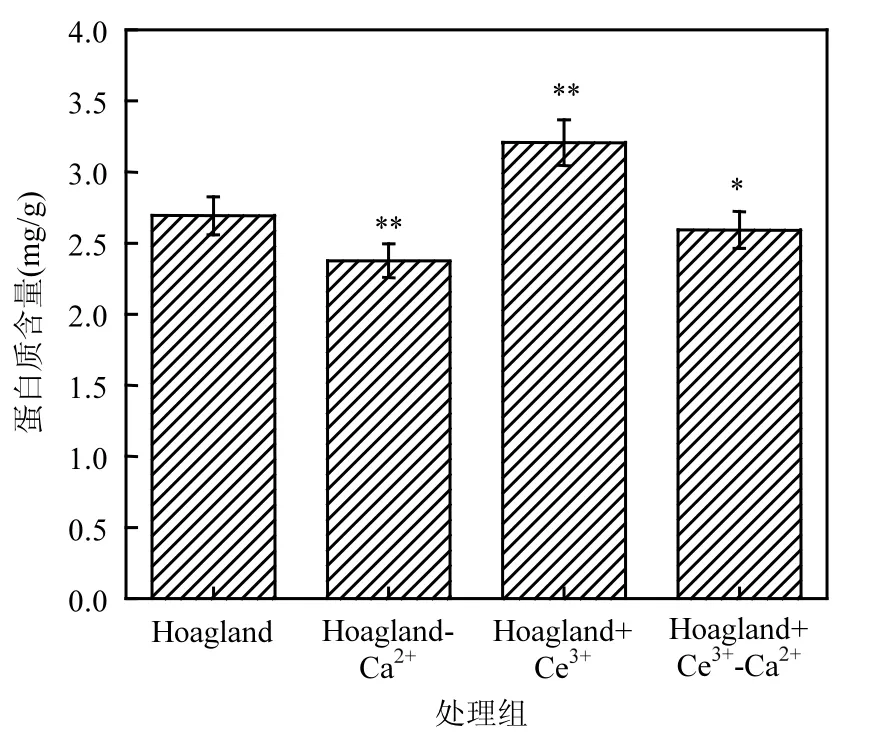
图7 不同培养液中凤眼莲蛋白质含量Fig.7 the protein content of Eichhornia crassipes indifferent culture solutions
植物细胞代谢的主要能源物质是淀粉,而主要的结构物质是蛋白质,光合作用中不管是光反应还是暗反应都需要多种酶的催化作用, 其大部分酶都是蛋白质.实验表明缺 Ca2+处理组的凤眼莲蛋白质含量相比于对照组降低较明显,但加Ce3+到缺 Ca2+的处理组中,明显看出 Ce3+减轻了缺Ca2+的损伤,蛋白质含量有多增加.在添加Ce3+的情况下,降低了因缺Ca2+而引起的损害程度,相比于单独缺 Ca2+处理组,Ce3+与 Ca2+有着相似的功能,Ce3+发挥着Ca2+的作用,促进酶的产生,保证光合作用的正常运行.
由统计分析可知,叶绿素含量和蛋白质含量之间的Pearson相关系数是0.935,表明两者具有相关性;叶绿素含量和 Ca2+-ATP酶活性之间的Pearson相关系数是0.968,并且在0.05水平上显著相关;蛋白质含量和 Mg2+-ATP酶活性之间的Pearson相关系数是0.998,并且在0.01 水平上显著相关.表明在不同处理条件下光合作用各指标间存在一定的相关性且相关性较显著,最终表现为稀土离子对植物钙相关光合作用的促进作用.
3 结论
3.1 不同 Ce3+浓度可以影响凤眼莲的生长,浓度为 0~1.2mg/L对凤眼莲的生长有促进作用,其中 1.2mg/L Ce3+促进作用最明显,但高浓度
(2.4mg/L)Ce3+可抑制凤眼莲的生长,甚至产生毒害作用.
3.2 适宜浓度的Ce3+可能替代Ca2+影响与钙相关的光合作用,在一定浓度范围内促进植物的光合作用,增加有机物质的合成,进而增加其生物量.在长江上游地区的四川支流中,由于稀土元素的协同作用,可能导致植物生长,最终表现为促进水生植物凤眼莲的生长,推进水体富营养化进程.
[1]徐光宪.稀土 [M]. 北京:冶金工业出版社, 2005.
[2]赵仕林,廖 洋,李瑞桢,等.四川地区稀土污染源调查及防控对策研究 [M]. 决策参考, 2009,184-203.
[3]Yu J, Chen W Q, Li J Q, et al. Study of the promoting mechanism of rare earth on water eutrophication in the upper reaches of the Yangtze River [J]. Disaster Advances, 2012,5(4):1595-1602.
[4]Matthew I L, Karen H J. Rare earth elements (REE) and yttrium in stream waters, stream sediments, and Fe-Mn oxyhydroxides: fractionation, speciation, and controls over REE+Y patterns in the surface environment [J]. Geochimicaet Comsmochimica Acta, 2008,72(24):5962-5983.
[5]胡勤海,金明亮.稀土元素在水体中的环境化学行为及其生物效应 [J]. 农业环境保护, 2006,25(5):274-277.
[6]张 辉,冯 嘉,朱为方.稀土高背景区生物链中稀土元素的分布特征 [J]. 中国稀土学报, 1999,17:365-368.
[7]钱 芸,戴树桂,刘广良,等.硝酸镧对铜绿微囊藻生长特性的影响 [J]. 中国环境科学, 2003,23(1):7-11.
[8]宋凌云,胡文月,赵继贞.稀土元素镧对满江红鱼腥藻的生理影响 [J]. 北京大学学报(自然科学版), 2000,36(6):123-128.
[9]张贝克,余 江,廖 洋,等.外源稀土 Ce3+对紫背浮萍光合作用和水体富营养化的影响 [J]. 中国稀土学报, 2012,30(6):236-242.
[10]周晓波,魏幼璋.稀土离子与Ca2+在生物体内的相互作用机制及应用 [J]. 生命科学研究, 1999,3(1):30-35.
[11]王伟列,涂楚桥,王光辉.稀土金属离子与酶分子的相互作用研究进展 [J]. 稀土, 1998,19(3):57-65.
[12]肖凤娟,常 虹,刘德龙,等.稀土离子与钙调素相互作用的研究进展 [J]. 稀土, 2003,2(6):64-68.
[13]Hong F S, Liu C, Zheng L. Formation of complexes of Rubisco-Rubisco activase from La3+, Ce3+treatment spinach [J]. Sci China Ser B, 2005(b),48(1):67-74.
[14]王 智,张志勇,张君倩,等.水葫芦修复富营养化湖泊水体区域内外底栖动物群落特征 [J]. 中国环境科学, 2012,32(1):142-149.
[15]凌 祯,杨具瑞,于国荣,等.不同植物与水力负荷对人工湿地脱氮除磷的影响 [J]. 中国环境科学, 2011,31(11):1815-1820.
[16]种云霄,胡洪营,钱 易,等.无机氮化合物及pH值对紫背浮萍生长的影响 [J]. 中国环境科学, 2003,23(4):417-421.
[17]丁士明,梁 涛,闫军才,等.有机配体对稀土元素在小麦体内积累和分异的影响 [J]. 生态学报, 2005,25(11):2888-2894.
[18]陈建勋.植物生理学实验指导 [M]. 广州:华南理工大学出版社, 2006.
[19]赵世杰,史国安,董新纯.植物生理学实验指导 [M]. 北京:中国农业科学技术出版社, 2002.
[20]孙 丽,宋长春,黄 耀.沼泽湿地N2O通量特征及N2O与CO2排放间的关系 [J]. 中国环境科学, 2006,26(5):532-536.
[21]Chaerle L, Saibo N, Van Der Straeten D. Tuning the pores: towards engineering plants for improved water use efficiency [J]. Trends Biotechnol., 2005,23(6):308-315.
[22]李青云,葛会波,胡淑明,等.钠盐和钙盐胁迫对草莓光合作用的影响 [J]. 西北植物学报, 2006,26(8):1713-1717.
[23]肖鹏程,朱端卫,罗 媛,等.环境因素对菹草根和叶细胞质膜 Ca2+-ATPase活性的影响 [J]. 中国环境科学, 2012,32(6):1123-1128.
[24]简令成,王 红.Ca2+在植物细rt5443胞对逆境反应和适应中的调节作用 [J]. 植物学通报, 2008,25(3):255-267.
[25]束良佐,孙五三.Ca2+浸种对酸雨伤害玉米幼苗的影响 [J]. 中国环境科学, 2001,21(2):185-188.
[26]Liang W J, Wang M L, Ai X Z. The role of calcium in regulating photosynthesis and related physiologicalindexes of cucumber seedlings under low light intensity and suboptimal temperature stress [J]. Scientia Horticulturae, 2009,123:34-38.
[27]李 强,曹建华,余龙江,等.干旱胁迫过程中外源钙对忍冬光合生理的影响 [J]. 生态环境学报, 2010,19(10):2291-2296.
Influences of rare earths (Ce3+) on calcium-regulated photosynthesis in Eichhornia crassipes
SONG Xue-mei1, XIA Chao1, LIAO Yang1,2*, ZHAO Shi-lin1, YU Jiang3(1.College of Chemistry and Material, Sichuan Normal University, Chengdu 610068, China;2.Engineering Research Center for the Development of Farmland Ecosystem Service Functions, Sichuan Province Institutions of Higher Education, Chengdu 610068, China;3.College of Architecture and Environment, Sichuan University, Chengdu 610065, China). China Environmental Science, 2014,34(9):2362~2367
Influences of rare earth Ce3+on calcium-regulated photosynthesis in Eichhornia crassipes were investigated. When cultured in Ce3+-containing nutrient solution without Ca2+, the chlorophyll produced in Eichhornia crassipes was found to decrease by 8.31% compared to that of counterparts cultured in Ca2+-containing nutrient solution (control experiment). The corresponding fluorescence quantum yields of chloroplast at 440and 480nm were also decreased by 6.14% and 3.82%, respectively. Additionally, the activities of Mg2+-ATP enzyme and Ca2+-ATP enzyme, and the amount of proteins were also decreased by 1.93%, 5.05% and 3.94%, respectively. However, the above parameters related to the calcium-regulated photosynthesis increased by 9.89%, 15.21%, 3.05%, 8.08%, 10.26% and 10.16%, respectively, when compared with those of Eichhornia crassipes cultured in nutrient solution without Ca2+. The photosynthesis activity of Eichhornia crassipes cultured in the absence of Ca2+had been significantly improved by the use of rare earth Ce3+. The experimental results have shown that compared with Ca2+, Ce3+exhibited similar regulation functions on the growth of Eichhornia crassipes. In the absence of Ca2+, a suitable amount of Ce3+will promote the photosynthesis of plant by increasing the plant biomass, thus resulting in positive catalytic effects on the process of eutrophication.
Ce3+;Ca2+;Eichhornia crassipes;photosynthesis
X171
A
1000-6923(2014)09-2362-06
宋雪梅(1987-),女,四川泸州人,四川师范大学硕士研究生,主要从事水体污染预防与控制研究.
2014-01-08
国家自然科学基金资助项目(31100374,51173122);四川师范大学校团队基金(025091)
* 责任作者, 副教授, Liaoyang_sicnu@163.com
