基于AFLP的垂乳银杏种质遗传多样性分析
2014-08-01吴岐奎邢世岩范圣明孙立民刘晓静
吴岐奎 邢世岩 范圣明 王 萱 孙立民 刘晓静
(1.山东农业大学林学院,山东 泰安 271018;2.中国林业科学研究院林业科技信息研究所,北京 100091)
银杏(GinkgobilobaL.)是银杏类植物经历第四纪冰川浩劫后孑遗下来的为数不多的物种之一,素有“活化石”的美称,是世界上现存最古老的高等植物。银杏原产中国,据文献记载三国时盛植江南,唐代已产于中原,宋朝更为普遍[1]。有关垂乳银杏的研究在中国、日本均有所报道,美国、欧洲等尚未有明确的报道[2]。北京大学李正理教授称银杏垂乳为“类钟乳枝或钟乳枝(stalactite-like branches)”,“钟乳”(“zhong-ru”),意思是钟乳石;江苏农学院何凤仁教授称之为“树奶”,银杏产区的群众称为“树瘤”、“撩子或撩”。银杏根部垂乳称“根钟乳”( 又名根奶、椅子根、根台),我国四川、重庆一带称“乳包”。 在日本称“垂乳”为“chichi”(nipple or breast) ,意思是乳头状凸起、奶子或树奶[3]。垂乳银杏主要分布在我国部分省市地区以及日本境内,以我国西南地区为主要分布地区。AFLP标记是目前较为常用的分子标记方法,具有方便快捷,只需极少的DNA材料,不需要预先知道基因组序列信息,多态性丰富、重复性好、稳定性高等优势,是检测种质遗传多样性和进行种质亲缘关系分析的有效工具[4-6]。
我国对垂乳银杏种质的研究主要集中在其个体发育及生物学意义上,在分子水平上对垂乳银杏的研究尚未有报道[7]。本研究采集我国云南、四川以及日本的优良垂乳银杏种质,使用AFLP分子标记技术对其进行遗传多样性和亲缘关系分析,为垂乳银杏种质资源的评价、保存、利用提供一定的理论基础和参考依据。
1 材料与方法
1.1 材料来源
2012年5月,在山东泰安市药乡林场银杏种质资源苗圃中,采集14个垂乳银杏种质嫁接苗(表1)的幼嫩叶片,放入自封袋中,再加入10倍质量的变色硅胶用于干燥,干燥过程中翻动2~3次,硅胶变色时及时更换,以保证叶片在12 h之内完全干燥,用于DNA的提取。

表1 垂乳银杏试验材料编号及来源
1.2 试验方法
1.2.1DNA的提取 称取垂乳银杏嫩叶约50 mg,垂乳银杏总DNA的提取参照Doyle等的方法[8],采用CTAB法提取幼嫩叶片总DNA。
1.2.2AFLP体系的建立 酶切与连接反应同时进行,内切酶为PstI和MseI,加入4 μL模板DNA(50 ng/μL)、1 μL Adapter(10 mmol/L)、2 μLPstI/MseI(10 U/μL)、2.5 μL 10×Reaction buffer、2.5 μL ATP(10 mmol/L)、1 μL T4 Ligase(5 U/μL)等构成20 μL酶切连接反应体系。将反应体系置于0.5 mL离心管中,混匀离心数秒,37 ℃保温5 h,8 ℃保温4 h,4 ℃过夜。
选择PstI(5’- GAC TGC GTA CAT GCA G- 3’)和MseI(5’- GAT GAG TCC TGA GTA A C- 3’)为预扩增引物。取2 μL酶切连接后的产物为模板,加入1 μL预扩增引物(10 μmol/L)、0.5 μL dNTPs(10 mmol/L)、2.5 μL 10×PCR buffer、0.5 μLTaqDNA聚合酶(2 U/μL)等构成25 μL预扩增体系。PCR扩增程序为:94 ℃预变性2 min; 94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸80 s,30个循环;72 ℃延伸5 min。
以预扩产物稀释20倍后作为选扩增模板,加入1 μLPstI引物(10 μmol/l)、1 μLMseI引物(10 μmol/L)、2.5 μL 10×PCR buffer、0.5 μL dNTPs(10 mmol/L)、0.5 μL的Taq酶(2 U/μL)等构成25 μL的选扩增体系。PCR程序为:94 ℃ 30 s,65 ℃ 30 s,72 ℃ 80 s进行第1轮扩增,以后每轮循环温度递减0.7 ℃,共扩增12轮,接着94 ℃ 30 s,55 ℃ 30 s,72 ℃ 80 s扩增23轮,然后72 ℃延伸5 min。选扩增产物用质量分数4%的聚丙烯酰胺凝胶电泳,获得的电泳图谱用ABI377 PRISM 377 sequencer测序仪(美国ABI公司)检测片段大小。
1.3 数据分析
电泳图谱经ABI377 PRISM377 sequencer测序仪检测片段大小,对迁移率相同的条带进行统计,经GENESCAN软件分析,得到原始数据,将原始数据中有带的换成“1”,无带的换成“0”,构建“01矩阵”,以供下一步分析。利用POPGENE version 1.31软件计算遗传多样性指标:多态性比例(PPB)、观测等位基因数(Na)、有效等位基因数(Ne)、Nei’s基因多样性(H)、Shannon’s信息指数(I)。采用NTSYSpc version 2.10e软件计算各单株间的遗传相似性系数(SC),并进行UPGMA聚类分析和主坐标分析以了解14个垂乳银杏种质间的遗传关系[9-10]。
2 结果与分析
2.1 垂乳银杏AFLP选择性扩增结果及多态性分析
引物组合P-GAA/M-CTG对垂乳银杏AFLP的选择性扩增结果见图1。
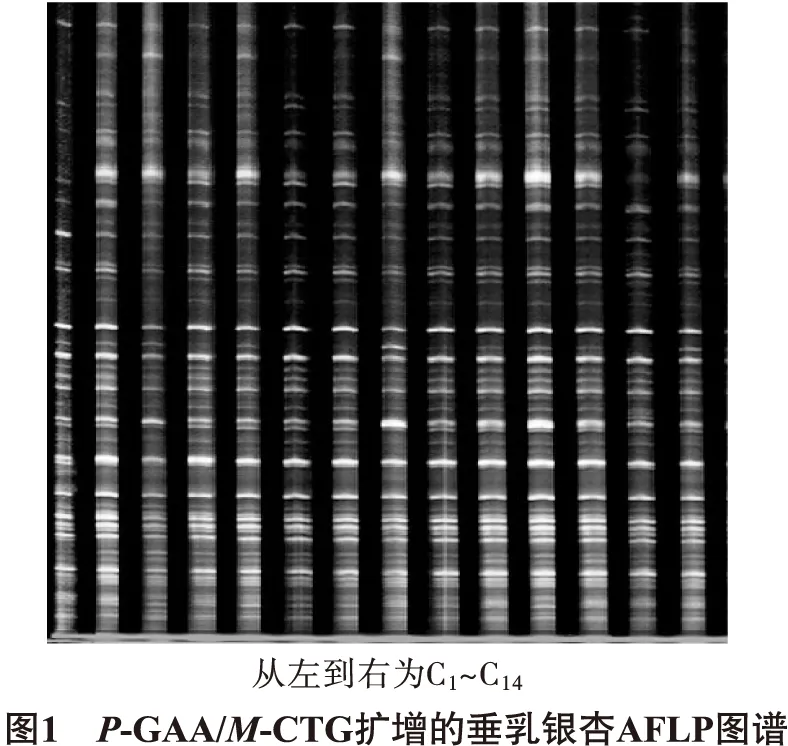
从64对引物组合中选出8对多态性较好的引物组合用于垂乳银杏AFLP的选择性扩增,由图1可知,图谱显示扩增多态性好,信号强度高,易于分辨,适合进行垂乳银杏遗传多样性统计分析。
用筛选出的8对引物进行选择性扩增,共产生935条谱带(表2),平均每对引物扩增产生117.88条谱带,其中多态带为920条,平均多态带比例为98.40%,不同引物的多态带比例为97.00%~100.00%。不同引物产生的谱带数量存在一定的差异,其中P-GAA/M-CTG引物产生的谱带最多(153条),而P-GAC/M-CTA引物产生的谱带最少(91条)。每对引物的鉴别效率为100%,由此可见:所选择的引物在垂乳银杏间表现出较高的多态性水平,说明AFLP是一种十分有效地分析垂乳银杏遗传多样性的方法。

表2 AFLP选择性扩增引物产生的条带多态性
运用POPGENE version 1.31软件对各位点观测的Na、Ne、H、I进行统计分析(表3)。14份垂乳银杏种质观测等位基因数(Na)平均值为 1.981 6,有效等位基因数(Ne)平均值为 1.457 9,Nei’s基因多样性(H)平均值为 0.276 5,Shannon信息指数(I)平均值为 0.429 3。由此可见,8对引物在所分析的垂乳银杏资源中多态性好,垂乳银杏的遗传多样性处于一个较高的水平。
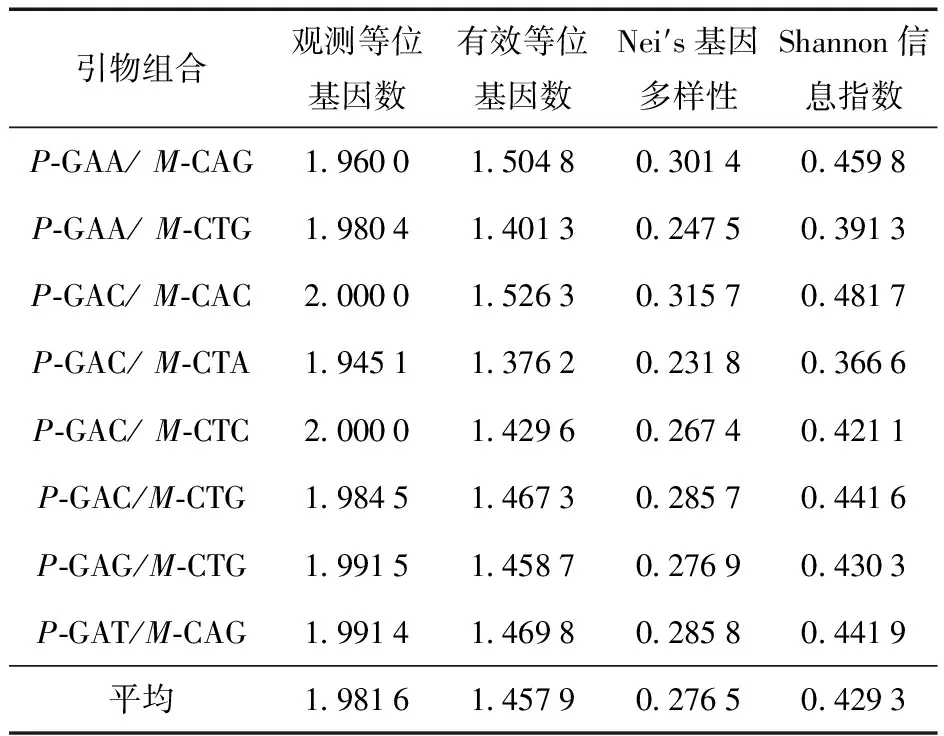
表3 基于不同引物组合的垂乳银杏遗传多样性水平
2.2 垂乳银杏种质资源的特异性位点分析
不同引物组合下垂乳银杏无性系特异性位点分析结果见表4。

表4 基于不同引物组合的垂乳银杏种质资源的特异位点
分析8对引物对14份垂乳银杏种质资源的电泳条带,产生210条特异性条带(包括单态带和缺失带),其中缺失带34条,占特异性总条带的16.19%。不同的垂乳银杏产生的特异性条带不同,C3产生的特异性条带最多,达到了47条,其中包括7条缺失带;其次是C7,达到了22条,其中包括4条缺失带。不同的引物组合产生的特异性条带也不相同,产生特异性条带最多的是P-GAA/M-CTG,共产生44条特异性条带,其中有3条缺失带;产生特异性条带最少的是P-GAC/M-CAC,共产生12条特异性条带,其中有3条缺失带。
2.3 垂乳银杏种质资源间的遗传关系分析
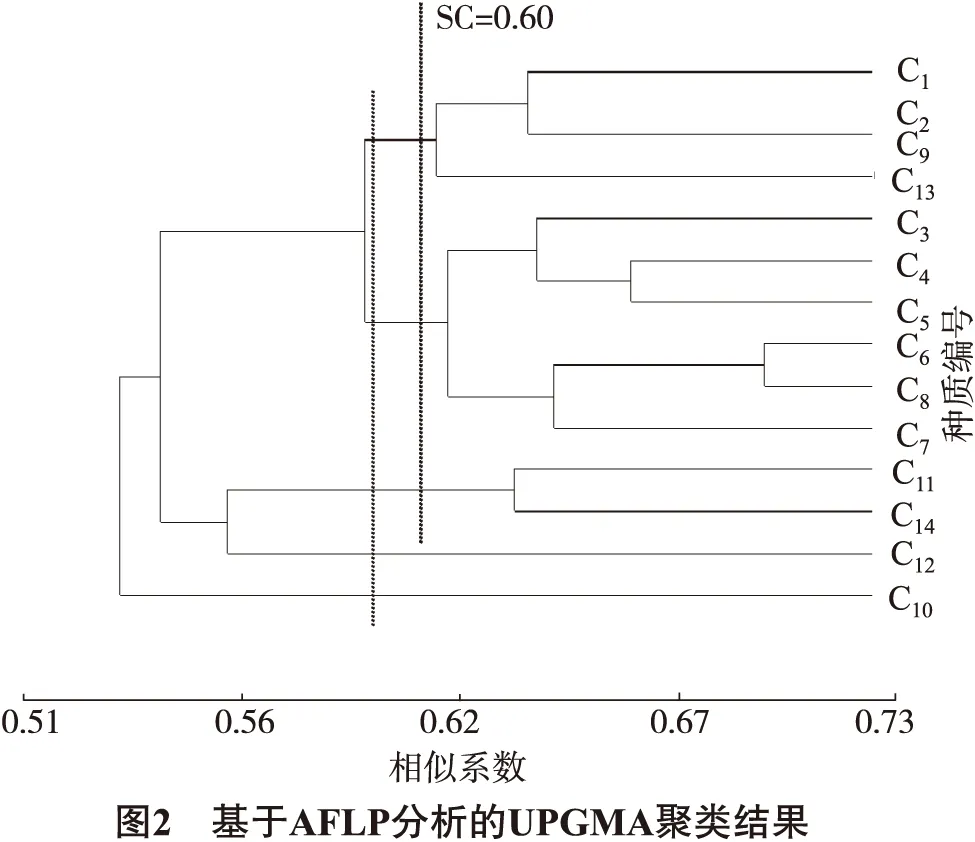
不同种质间的遗传相似系数为 0.440 4~0.729 9,平均值为 0.475 2。C1与C2的遗传相似系数最大(0.729 9),说明二者的亲缘关系最相近,遗传差异性最小。C7与C12的遗传相似系数最小(0.440 4),说明二者的亲缘关系最远,差异性最大。C2与其他垂乳银杏种质的相似性系数平均值最大(0.638 6);C10与其他垂乳银杏种质的相似性系数平均值最小(0.506 0),说明C10与其他种质相似性低,亲缘关系远。
将14个垂乳银杏种质进行UPGMA聚类(图2),在相似系数0.51处,供试材料分为2大类:I类包括13个种质;II类包括1个种质,即C10。在相似系数0.60处,可将垂乳银杏分为5类:第1类包括4个种质,即C1、C2、C9、C13;第2类包括6个种质,即C3、C4、C5、C6、C7、C8;第3类包括2个种质,即C11、C14;第4类包括1个种质,即C12;第5类包括1个种质,即C10。
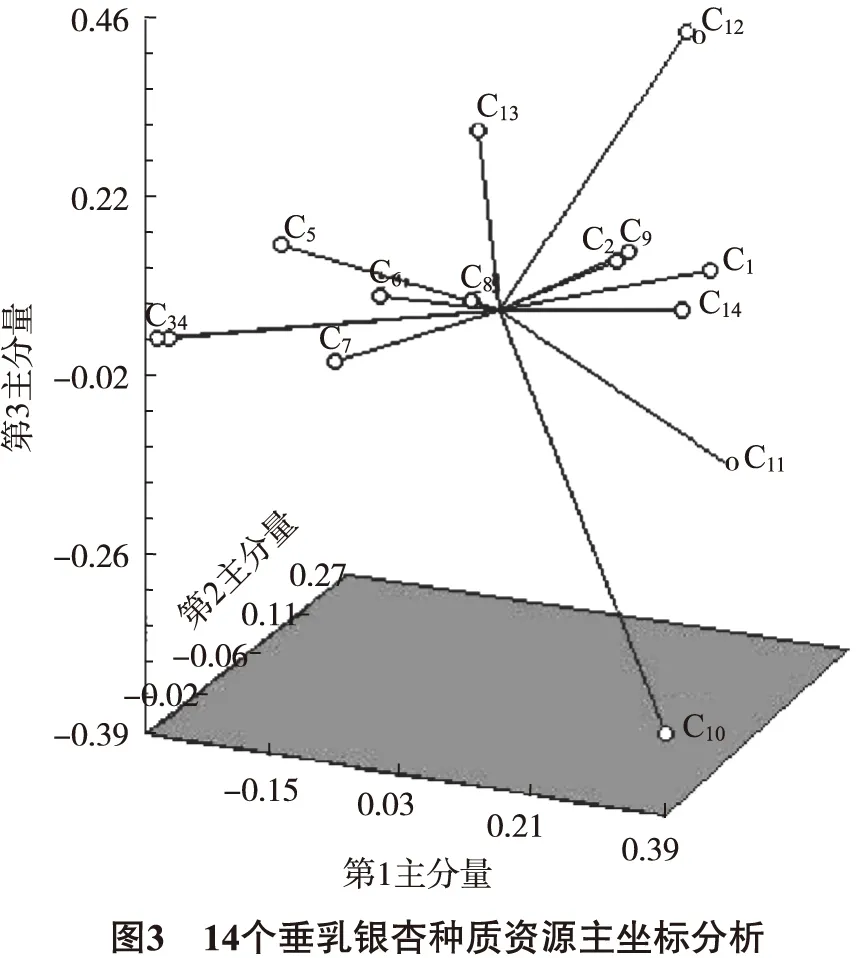
基于DICE遗传相似矩阵(GS),通过主坐标分析构建垂乳银杏种质资源的三维分布图,以进一步了解种质间的遗传关系(图3)。结果显示:第1主分量特征值为 0.941 4,贡献率16.40%;第2主分量特征值为 0.567 3,贡献率为9.88%;第3主分量特征值为 0.532 3,贡献率为9.27%,前3个主分量累计贡献率达到35.55%。从图3可以看出,来自云南腾冲的垂乳银杏种质较为聚拢,来自四川万源的垂乳银杏种质较为分散;其中C10种质与其他种质关系较远。主坐标分析结果与UPGMA聚类分析结果基本一致。
3 结论与讨论
8对引物共检测到935条谱带,其中有920条多态带,平均多态带比例为98.40%,平均每对引物产生115条多态带。8对引物共产生210条特异性条带,包括34条缺失带。14份垂乳银杏种质Nei’s基因多样性(H)平均值为 1.981 6,Shannon信息指数(I)平均值为 0.429 3。在相似系数0.60处,可将垂乳银杏分为5类。来自同一地区的垂乳银杏并没有完全聚在一起,说明垂乳银杏间的亲缘关系没有严格的相关性。
AFLP采用的限制性内切酶及选择性碱基的种类、数目有很多,因此理论上能产生的标记数目是无限的,能比较全面地揭示不同物种或种质资源的遗传特征,非常适用于亲缘关系较近材料的研究。AFLP能够稳定、高效地扩增垂乳银杏种质基因组DNA,是研究垂乳银杏种质遗传多样性的有效方法。
王利等研究了29个银杏雄株种质和21个观赏品种之间的遗传关系[11-13],遗传多样性反映了一个物种的环境适应力、生存能力、进化潜力及被改造和利用的空间。21个银杏观赏品种的多态带比例为88%;各品种间的相似系数为0.45~0.85,表明它们存在不同程度的遗传差异。本研究中14种垂乳银杏多态带比例为98.40%,高于21个观赏银杏品种的多态性,表明它们遗传差异大,遗传多样性丰富。
通过UPGMA聚类分析和主坐标分析,来自云南的种质遗传关系较为聚拢,来自四川的种质遗传关系较为分散,来自日本的种质并无明显的特异性,这与Y.Tsumura等的研究相一致,中国与日本银杏亲缘关系近,这与公元6世纪银杏从中国传至日本有关[14]。
S. K. Singh等采用AFLP技术对印度西南地区的20株银杏进行分析,表明地理来源与遗传距离之间没有确定的关系[15]。本研究中,来源相同的垂乳银杏种质并没有严格的聚集到一起,说明其变异与地理来源无严格的相关性,这与王利等的研究结果基本一致。由此可见,在垂乳银杏育种和研究利用过程中,不仅要考虑材料地理来源的差异,更要考虑材料间遗传差异的大小。
要全面了解垂乳银杏的遗传多样性,还需进一步的研究,结合其他的DNA分子标记技术在更大种质范围内,从多角度、多层面研究垂乳银杏的遗传多样性,为垂乳银杏种质的保护、利用、引种提供理论支持。另外,对垂乳银杏种质的取材不够全面、数量偏低是本研究的一大缺憾,在以后的研究中要有必要适当扩大取材范围。
[1] 邢世岩.中国银杏种质资源[M].北京: 中国林业出版社, 2013.
[2] 邢世岩,张倩,付兆军,等.银杏垂乳个体发生及系统学意义[J]. 林业科学, 2013, 49(8): 108-116.
[3] 邢世岩.银杏树瘤[J]. 植物杂志, 1996(3): 29-30.
[4] 林斌彬,张子平,王艺磊.AFLP 技术发展及其在水产生物学研究中的应用[J].集美大学学报: 自然科学版,2008,13( 1) : 45-51.
[5] Ahuja M R. Recent advances in molecular genetics of forest trees[J]. Euphytica, 2001, 121(2): 173-195.
[6] Ferdinandez Y S N, Coulman B E. Genetic relationships among smooth bromegrass cultivars of different ecotypes detected by AFLP markers[J]. Crop science, 2004, 44(1):241-247.
[7] 付兆军,邢世岩,刘莉娟,等. 银杏苗基生垂乳的生长特性[J]. 西南林业大学学报, 2013,33(1): 34-38.
[8] Doyle J J, Doyle J L. Isolation of plant DNA from fresh tissue[J]. Focus, 1990,12: 13-15
[9] 陈云坪.基于数字图像处理的DNA 指纹图谱分析系统的设计与算法研究[D]. 重庆:西南农业大学, 2004.
[10] 曹永生,孔繁胜,王宇生. 基于图象处理的种质资源指纹图谱分析[EB/OL][2013-12-26]. 中国作物种质信息网. http: //icgr. caas. net. cn/training/forum/基于图象处理的种质资源指纹图谱分析. htm.
[11] 王利,邢世岩,杨克强,等.银杏观赏品种亲缘关系的 AFLP 分析[J].遗传学报,2006,33(11):1020-1026.
[12] 王利,邢世岩,韩克杰,等.银杏雄株亲缘关系的 AFLP 分析[J].中国农业科学,2006,39(9):1940-1945.
[13] 王利,邢世岩,王芳,等.银杏雌株种质遗传多样性的AFLP分析[J].林业科学,2008,44(4):47-53.
[14] Tsumura Y, Ohba K. The genetic diversity of isozymes and the possible dissemination ofGinkgobilobain ancient times in Japan[M]//GinkgobilobaA Global Treasure. Springer Japan,1997: 159-172.
[15] Singh S K, Rajkumar S. Estimation of genetic diversity inGinkgobilobatrees from northwestern India using AFLP and microsatellite markers[J]. Journal of Plant Genetics and Transgenics, 2010, 1(1): 16-20.
