青海湖流域沙柳河湿地草地和具鳞水柏枝灌丛小气候特征研究
2014-07-02李岳坦李小雁
李岳坦,李小雁
(1.北京师范大学 地表过程与资源生态国家重点实验室,北京 100875;2.中国科学院大学,北京 100049)
青海湖流域沙柳河湿地草地和具鳞水柏枝灌丛小气候特征研究
李岳坦1,2,李小雁1
(1.北京师范大学 地表过程与资源生态国家重点实验室,北京 100875;2.中国科学院大学,北京 100049)
本文利用波文比系统和ECH2O土壤水分监测系统在青海湖流域沙柳河下游同一河道断面监测的两种典型湿地植被群落类型——草地和具鳞水柏枝灌丛的空气温度、相对湿度、净辐射、土壤热通量和土壤温度等数据,对流域内两种湿地植被群落的小气候特征进行了研究,研究发现:受大气候的变化影响,草地和具鳞水柏枝灌丛的空气温度日变化呈现出相同的变化趋势,在整个生长季具鳞水柏枝灌丛月平均值显著高于草地,具鳞水柏枝灌丛群落上层的空气温度高于群落下层,而草地上下层没有显著差异;具鳞水柏枝灌丛和草地空气相对湿度日变化规律相同,白天较低,晚上较高,在整个植物的生长季,草地和具鳞水柏枝灌丛空气相对湿度差异显著;草地和具鳞水柏枝灌丛群落冠层净辐射的日变化均呈单峰曲线,在7—9月份,具鳞水柏枝灌丛冠层与草地冠层的净辐射差异显著;草地和具鳞水柏枝灌丛地下5 cm土壤热通量的日变化呈单峰曲线,月变化趋势较为类似,具鳞水柏枝灌丛变化幅度显著大于草地;在整个植被生长季草地和具鳞水柏枝灌丛地下10 cm土壤温度日变化曲线呈相似的形状,月变化趋势基本一致,但是草地的月均值大于具鳞水柏枝灌丛;草地在白天和夜晚均由大气向地表输送热量,具鳞水柏枝灌丛在白天由大气向地表输送热量,而在晚上基本是由地表向大气输送热量;在两种湿地植被群落中,白天是水汽蒸发的过程,而夜晚是水汽凝结的过程,在地表与大气的能量交换中,潜热占主导。
青海湖流域;沙柳河湿地;具鳞水柏枝灌丛;草地;小气候
青海湖流域位于青藏高原东北部,地处高寒半干旱地区,是国际重要湿地。流域内自然条件比较优越,雨热同季,牧草茂盛,是青海省的优良牧场、重要的农牧业基地和经济发展区。但是由于自然环境的变化和人类活动的综合影响(冯宗炜和冯兆忠,2004),流域内的生态环境不断恶化,同时由于过度放牧等原因,流域内草地退化现象普遍,载畜量大大降低。此外,灌丛植被也由于人为掠夺樵采,几近消亡(杨修等,2003;董得红和王世红,2006;张学元,2006;刘进琪等,2007),流域内的植被退化对整个流域的农牧业发展具有制约作用。
小气候是生物生长发育最重要的环境因子,不同的植物群落形成不同的小气候环境,而各异的小气候生境又构成了不同的生物群落。群落小气候的形成是植被与环境综合作用的结果,是群落质量的综合反应,也是退化植被恢复与重建效果评价的一个重要指标。国内外部分学者已经对不同植物群落的小气候特征进行了研究(McCafferty et al,2003;李宗峰等,2005;司建华等,2005;Heithecker and Halpern,2006;Lott et al,2009;Rambo and North,2009),但是对湿地生境下植被群落的小气候研究较少(付为国等,2006),特别是对高寒半干旱地区的植被群落的小气候研究尚未见报道。本文利用波文比系统和ECH2O土壤水分监测系统在青海湖流域沙柳河下游同一河道断面监测的两种典型湿地植被群落类型——草地和具鳞水柏枝(Myricaria squamosa Desv.)灌丛的空气温度、相对湿度、净辐射、土壤热通量和土壤温度等数据,对流域内两种湿地植被群落的群落小气候特征进行了研究,以揭示湿地植物群落演替的生态学过程,为受损群落的生态修复提供理论和技术依据。
1 材料与方法
1.1 试验区概况
本试验在位于青海省刚察县境内的沙柳河下游河段进行,试验区域地理坐标:东经100°21′,北纬37°23′,海拔3206 m。草地土壤质地为沙壤土,草地以二裂委陵菜(Potentilla bifurca Linn.)、毛穗赖草(Leymus paboanus (Claus.) Pilger.)和短穗兔耳草(Lagotis brachystachya Maxim.)为优势物种,伴生种有鹅绒委陵菜(Potentilla ansrina Rosaceae)、荸荠(Heleocharis dulcis (Burm. F.) Trin. ex Henschel)、戟叶火绒草(Leontopodium dedekensii (Bur. et Franch.) Beauv.)、矮嵩草(Kobresia humilis (C. A. Mey ex Trauvt.) Sergievskaya.)和海乳草(Glaux maritima L.)。具鳞水柏枝灌丛土壤质地为沙土,多含砾石。
1.2 具鳞水柏枝灌丛和草地的微气象测定
选择沙柳河下游的某一河段,沿垂直于河道的方向在草地上布设了波文比系统(图1、2),于2009年5月17日至10月16日对草地和具鳞水柏枝灌丛的小气候进行测定。测定的指标主要有草地1 m、1.5 m的相对湿度,1 m、1.5 m的空气温度,1.3 m净辐射及地下0.05 m的土壤热通量;具鳞水柏枝灌丛2.2 m、2.7 m的相对湿度,2.2 m、2.7 m的空气温度,2.3 m净辐射和地下0.05 m的土壤热通量。

图1 草地微气象观测Fig.1 Micrometeorology data measurement of grassland

图2 具鳞水柏枝灌丛微气象观测Fig.2 Micrometeorology data measurement of Myricaria squamosa Desv. shrubs
1.3 显热通量和潜热通量的计算方法
热量交换系数和水汽交换系数相等是波文比—能量平衡法的一个合理假设条件,同时也是波文比—能量平衡法成立的一个至关重要的条件。梯度扩散理论认为,近地层大气只有因垂直的水汽热量输送过程而形成相应的垂直温度和湿度梯度,没有水平输送过程的影响。下垫面能量平衡的方程如下所示(Mastrorilli et al,1998;Savage et al,2009;Zhang et al,2010):



式中,Rn是净辐射;LE是潜热通量;H是显热通量;e是空气相对湿度;T是空气温度;ρ是空气密度;Cp是空气定压比热;ε是空气水汽分子对空气分子重量比;P是大气压;Kw是潜热交换系数;Kh是感热交换系数。
根据相似理论,假设Kw= Kh,同时引入波文比β(潜热通量与潜热通量之比),并将微分方程转化为差分方程:
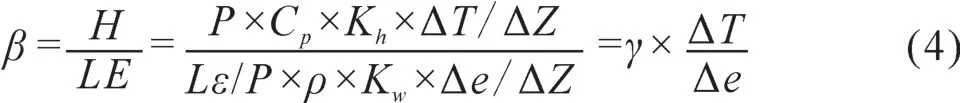
式中,γ为湿度常数系数;则公式可以简化为:

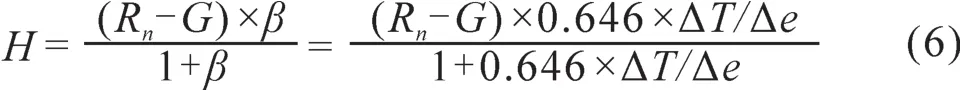
2 结果与分析
2.1 空气温度
植被群落内的空气温度既受大气候的影响,又受植被群落生长状况的影响,不同植被群落类型的差异会导致植被群落内部空气温度变化的差异。图3和图4分别为整个植被生长季草地和具鳞水柏枝灌丛平均气温的日变化和月平均值变化趋势。从图3可以看出,二者的空气温度日变化表现出相同的变化趋势,其中最低值出现在早晨6时左右,最高值出现在15—16时,这是由大气候的变化决定的。进而通过显著性t检验发现,在5月份(检验结果为|t|=1.886<t0.01/2=2.750)草地和具鳞水柏枝灌丛的月平均气温值较为接近,而在6—9月份(检验结果分别为|t|=3.699>t0.01/2=2.660、|t|=25.726>t0.01/2=2.660、|t|=2.939>t0.01/2=2.750、|t|=27.793>t0.01/2=2.750)具鳞水柏枝灌丛的月平均气温值显著大于草地的月平均气温值(图4);这是因为在青海湖流域5月份是植被开始萌发的季节,植被群落对小气候的影响小于大气候对群落内小气候的影响,而在6—9月份是植被生长相对茂盛的季节,受植被生长状况不同的影响,具鳞水柏枝灌丛和草地的小气候差异显著从而导致二者空气温度差异显著。此外,在整个生长季具鳞水柏枝灌丛的空气温度月平均值显著大于草地的空气温度月平均值(显著性t检验的结果为|t| = 4.473 > t0.01/2= 2.617)。
从垂直方向上来看,在整个生长季,具鳞水柏枝灌丛2.7 m处的空气温度月平均值为10.5℃,2.2 m处的空气温度月平均值为10.2℃,经显著性t检验,|t|=3.388>t0.01/2=2.576,说明在具鳞水柏枝灌丛林冠上方或群落上层的空气温度高于群落下层,这与黑河流域的胡杨和柽柳群落(司建华等,2005)以及北热带岩溶植被的乔、灌木群落(邓艳等,2004)的状况相一致,而草地1.0 m和1.5 m的空气温度月平均值均为9.3℃,经显著性t检验,|t|=2.196<t0.01/2=2.576,上下层没有显著差异。这是因为对灌木林地来讲,林冠是主要热源,林内乱流交换弱,林冠吸收的热量向下传递慢,而草地没有林冠作用,吸收的热量很快到达下部(邓艳等,2004)。
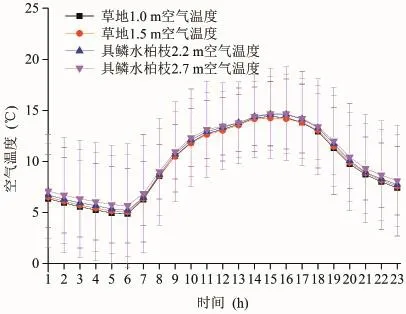
图3 草地和具鳞水柏枝灌丛不同高度空气温度的日变化Fig.3 Air temperature daily change of different altitude between grassland and Myricaria squamosa Desv. shrubs
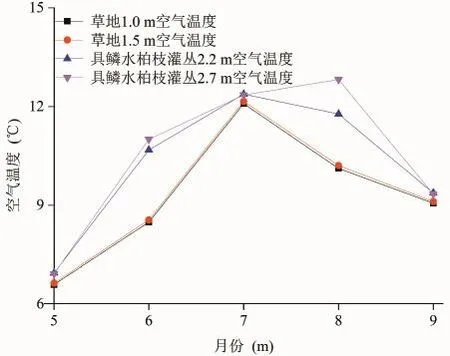
图4 草地和具鳞水柏枝灌丛不同高度空气温度的月平均值变化Fig.4 Air temperaturemonthly change of different altitude between grassland and Myricaria squamosa Desv. shrubs
2.2 相对湿度
图5和图6分别为整个植被生长季草地和具鳞水柏枝灌丛空气相对湿度的日变化和月平均值变化趋势,从图5可知,在整个植被生长季,两种植被类型空气相对湿度的日变化呈U型曲线,总体来说,白天的空气相对湿度较低,晚上的空气相对湿度较高,最高值出现在早晨6时左右,约为87%,最低值出现在15—16时,为54%左右,这是因为群落空气相对湿度受群落空气温度的影响较大,早晨6时空气温度最低,饱和水汽压最小,从而导致相对湿度最大,而在15—16时空气温度最高,饱和水汽压最大,因而相对湿度最小;从图6可以看出,草地和具鳞水柏枝灌丛的月平均相对湿度表现出不同的变化规律;此外,经过显著性t检验发现,在5月份和8月份,草地和具鳞水柏枝灌丛的相对湿度没有显著差异(显著性t检验结果分别为|t|=2.619<t0.01/2=2.756、|t|=0.554<t0.01/2=2.660),在6、7和9月份,两种植被类型的相对湿度的差异显著(显著性t检验结果分别为|t|=2.766>t0.01/2=2.660、|t|=16.800>t0.01/2=2.660、|t|=13.000>t0.01/2=2.660),并且具鳞水柏枝灌丛的相对湿度略大于草地的相对湿度,这与具鳞水柏枝灌丛距河道较近,受河道内水面蒸发量的影响相对较大有关,而造成5月份和8月份二者没有显著差异的原因可能是在5月份,河道内河水发生断流,没有河道内水面蒸发对二者相对湿度的影响,在8月份,由于在中上旬有几次较大的降水,造成草地和具鳞水柏枝灌丛均被洪水淹没,这样二者由于均受水面蒸发量的影响,从而导致相对湿度差异不显著;此外,在整个植物的生长季,草地和具鳞水柏枝灌丛平均相对湿度亦表现出显著性差异(显著性t检验的结果为|t|=2.071>t0.01/2=2.576),分别为69.6%和70.7%。

图5 草地和具鳞水柏枝灌丛不同高度空气相对湿度的日变化Fig.5 Relative humidity daily change of different altitude between grassland and Myricaria squamosa Desv. shrubs

图6 草地和具鳞水柏枝灌丛不同高度空气相对湿度的月平均值变化Fig.6 Relative humiditymonthly change of different altitude between grassland and Myricaria squamosa Desv. shrubs
2.3 净辐射
净辐射是作用面辐射能收入与支出的差额,即一切辐射能的净通量,是作用面热量、动量和水分交换的主要动力(刘淑明等, 2005)。图7和图8分别为整个植被生长季草地和具鳞水柏枝灌丛植被冠层净辐射的日变化和月平均值变化趋势,从图7可知,在整个植被生长季,草地和具鳞水柏枝灌丛冠层净辐射的日变化均呈单峰曲线,从早上7时至晚上19时为正值,其余时段为负值,最大值出现在13时;此外,经显著性t检验发现,从上午10时至下午17时具鳞水柏枝灌丛冠层的净辐射要大于草地冠层的净辐射(检验结果为|t|=7.719>t0.01/2=3.499),说明在这个时段内具鳞水柏枝灌丛冠层与大气之间的能量交换要大于草地冠层与大气之间的能量交换。
图8为草地和具鳞水柏枝灌丛的冠层净辐射月平均值变化,从图中可以看出,草地和具鳞水柏枝灌丛的冠层净辐射的月变化表现出较为相似的趋势;此外,经过显著性t检验发现,在5月份和6月份,草地和具鳞水柏枝灌丛冠层的净辐射没有显著差异(t检验结果分别为|t|=0.958<t0.01/2=2.977、|t|=1.571<t0.01/2=2.756),而在7—9月份,草地和具鳞水柏枝灌丛冠层的净辐射差异显著(t检验结果分别为|t|=2.779>t0.01/2=2.750、|t|=3.886>t0.01/2=2.750、|t|=3.129>t0.01/2=2.756),这是造成7—9月份草地和具鳞水柏枝灌丛群落内空气温度差异的原因之一。
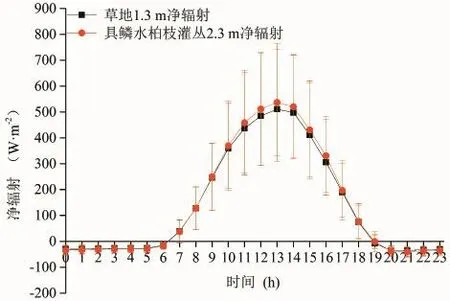
图7 草地和具鳞水柏枝灌丛冠层净辐射的日变化Fig.7 Daily change of canopy net radiation between grassland and Myricaria squamosa Desv. shrubs
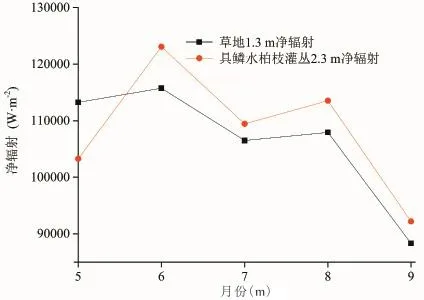
图8 草地和具鳞水柏枝灌丛冠层净辐射的月平均值变化Fig.8 Monthly change of canopy net radiation between grassland and Myricaria squamosa Desv. shrubs
2.4 土壤热通量
土壤热通量是土壤表层与下层的热交换,它是影响陆地表面能量平衡的重要因素之一(闵文彬,2009)。在研究地表能量平衡问题时,需要精确计算土壤中的热量传输(吴红颜等,2001;贺芳芳,2008;颜江良等,2008)。图9和图10分别为整个植被生长季草地和具鳞水柏枝灌丛地下5 cm土壤热通量的日变化和月平均值变化趋势,从图9可知,在整个植被生长季,草地和具鳞水柏枝灌丛地下5 cm土壤热通量的日变化均呈单峰曲线,草地地下5 cm土壤热通量从上午10时至下午20时的时段内为正值,其余时段为负值;具鳞水柏枝灌丛地下5 cm土壤热通量从上午10时至下午18时的时段内为正值,其余时段为负值;草地地下5 cm土壤热通量的最大值出现在14时,为29.1 W·m−2,而具鳞水柏枝灌丛地下5 cm土壤热通量的最大值出现在13时,为33.2 W·m−2。土壤热通量的变化通常是以净辐射的变化为基础的,由于植被冠层及群落小环境的影响,土壤热通量对净辐射的反馈存在延时现象(申双和和崔兆韵,1999;李明财等,2008),通过上述对本地区冠层净辐射的研究发现,该地区草地和具鳞水柏枝灌丛的冠层净辐射最大值均出现在13时左右,而草地和具鳞水柏枝灌丛的土壤热通量最大值分别发生在14时和13时,说明草地的地下5 cm土壤热通量与冠层净辐射存在1小时的延时,而具鳞水柏枝灌丛的地下5 cm土壤热通量与冠层净辐射不存在延时现象,这可能与两种植被类型生境的土壤结构、性质、土壤含水量的水平差异以及植被冠层的差异性有关(王旭等,2005)。

图9 草地和具鳞水柏枝灌丛地下5 cm土壤热通量的日变化Fig.9 Daily change of soilheat f ux between grassland and Myricaria squamosa Desv. shrubs
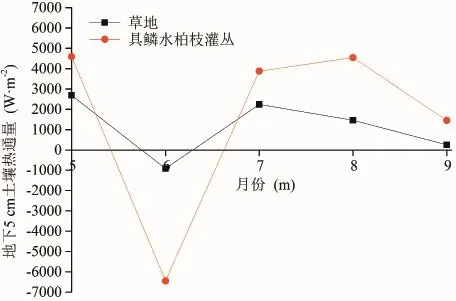
图10 草地和具鳞水柏枝灌丛地下5 cm土壤热通量的月平均值变化Fig.10 Monthly change of soilheat f ux between grassland and Myricaria squamosa Desv. shrubs
从图10可以看出,草地和具鳞水柏枝灌丛的地下5 cm土壤热通量月变化趋势较为类似,在6月份具鳞水柏枝灌丛和草地地下5 cm土壤热通量的月平均值为负,而在7、8、9月份为正;此外具鳞水柏枝灌丛的5 cm土壤热通量的变化幅度显著大于草地地下5 cm土壤热通量的变化幅度,这可能与二者之间的土壤物理结构不同有关,草地的土壤类型为沙壤土,具鳞水柏枝灌丛的土壤类型为砂土(图11)且含砾石较多,相对于草地的土壤类型而言,具鳞水柏枝灌丛的土壤导热性强,持水性差,因而对净辐射的反映较为敏感,土壤热通量的变化幅度大。
2.5 土壤温度
土壤温度作为一种重要的环境因子会直接或间接的对植被的生长产生影响(Warrington and Kanemasu,1983;Bonan,1989;Dang and Cheng,2004;Nabi and Mullins,2008),植物的许多生理过程诸如气孔导度、蒸腾、营养的迁移转化以及二氧化碳的排放都受其影响(DeLucia,1987;Al-Hamdani et al,1990;DeLucia et al,1991;Harper and Camm,1993;Landhäusser et al,1996;McMichael and Burke,1996),较高的土壤温度会对根的生长、呼吸以及营养吸收产生影响(Kaspar and Bland,1992;Boone et al,1998;Nabi and Mullins,2008),来自地表的热辐射会对植被的冠幅产生影响(Timlin et al,2002)。
图12和图13分别为整个植被生长季草地和具鳞水柏枝灌丛地下10 cm土壤温度的日变化和月平均值变化趋势,从图12可知,在整个植被生长季草地和具鳞水柏枝灌丛地下10 cm土壤温度日变化曲线呈相似的形状,草地地下10 cm土壤温度在18—19时达到最高值,之后逐渐下降,在次日9时出现最低值,具鳞水柏枝灌丛的最高值出现在15—16时,之后逐渐下降,在次日8时到达最低值。与空气温度相比,空气温度的最高值出现在15—16时,最低值出现在次日早上6时,说明具鳞水柏枝灌丛的10 cm土壤温度对空气温度的响应更为敏感,这跟具鳞水柏枝灌丛的土壤条件有关,具鳞水柏枝灌丛的土壤为砂土且含砾石较多,相对于沙壤土温度更容易发生变化(图11)。

图11 草地和具鳞水柏枝灌丛的土壤剖面Fig.11 Soil prof le of grassland and Myricaria squamosa Desv. shrubs
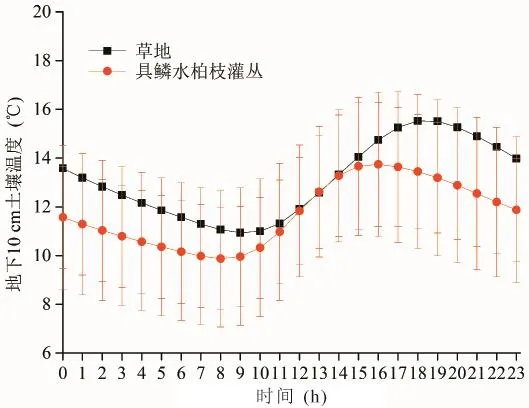
图12 草地和具鳞水柏枝灌丛地下10 cm土壤温度的日变化Fig.12 Daily change of soiltemperature between grassland and Myricaria squamosa Desv. shrubs

图13 草地和具鳞水柏枝灌丛地下10 cm土壤温度的月平均值变化Fig.13 Monthly change of soiltemperature between grassland and Myricaria squamosa Desv. shrubs
此外,从图13可以看出,草地和具鳞水柏枝灌丛的地下10 cm土壤温度月变化趋势基本一致,但是草地的地下10 cm土壤温度月均值大于具鳞水柏枝灌丛的10 cm土壤温度月均值,显著性t检验结果为|t|=16.169>t0.01/2=2.617,进一步验证了二者之间的差异性,这是因为具鳞水柏枝灌丛距离河道较近,地下水埋深不大,在整个植被生长季,实测的具鳞水柏枝灌丛的地下水埋深为0 ~56 cm,而草地的地下水埋深比具鳞水柏枝灌丛要高100 cm左右,并且由于具鳞水柏枝灌丛距离河道近,地下水与河水之间的水分交换密切,通过水分的转移带走了土壤内部的部分热量,使土壤温度不容易升高。
2.6 草地和具鳞水柏枝灌丛显热通量和潜热通量的变化
显热通量是由于对流作用产生向上的热流,代表着地表能量的损失(胡顺军等,2006);潜热通量主要是由于地面蒸发和大气凝结潜热所致,它表征了地表植被水分蒸散量的多少,其能量交换与水的相变相联系(李英年等,2003)。根据公式5和公式6,计算了在整个生长季内草地和具鳞水柏枝灌丛的显热通量和潜热通量,并绘制了二者的日变化趋势图(图14和图15),从图中可以看出,草地的显热通量在白天9—19时均为负值,在−3.8 ~ −59.4 W·m−2波动,13时达到最低值,说明在白天的这个时段内是由大气向地表输送热量,而在20时至次日8时之间有正负波动,但就平均而言为负值,约为−3.8 W·m−2,说明晚上也基本是由大气向地表输送热量;具鳞水柏枝灌丛的显热通量在白天7—18时均为负值,在−8.1 ~ −92.8 W·m−2波动,15时达到最低值,说明在白天在这个时段内是由大气向地表输送热量,而在19时至次日8时之间有正负波动,但就平均而言为正值,约为0.8 W·m−2,说明晚上基本是由地表向大气输送热量,但是很微弱;草地潜热通量的变化规律是7—18时为正值,13时到达最高值,为546.6 W·m−2,其余时段为负值,说明白天是水汽蒸发的过程,而夜晚是水汽凝结的过程;具鳞水柏枝灌丛潜热通量的变化规律与草地潜热通量的变化规律类似,在7—18时为正值,在14时到达最高值,为579.6 W·m−2,其余时段为负值,说明具鳞水柏枝灌丛在白天是水汽蒸发的过程,而夜晚是水汽凝结的过程;总的来说,草地和具鳞水柏枝灌丛的显热通量在白天均为负值,而潜热通量在白天均为正值,这与湿润地区的农田以及宁南半干旱区草地和农田的特征基本一致(Jerald and Kenneth,2000;张晓煜等,2005)。
此外,从图14和图15还可以看出,草地和具鳞水柏枝灌丛的显热通量和潜热通量的标准偏差较大,在白天尤为显著,说明每日的显热通量和潜热通量差异明显,这是因为在青海湖流域每日的气候变化多端所致。
图16和17分别为6—9月份草地和具鳞水柏枝灌丛的显热通量与潜热通量的月值柱状图,从图中可以看出,草地和具鳞水柏枝灌丛在6—9月份的潜热通量皆为正值,且6月份的潜热通量最大,之后逐渐降低,而显热通量只有草地在8月份为正,其余月份全为负值,并且显热通量的绝对值显著小于潜热通量的绝对值,说明在地表与大气的能量交换中,潜热占主导,即地表主要通过潜热向大气输送热量,而显热的作用主要是将大气的热量输送到地表。

图14 生长季草地显热通量(H)和潜热通量(LE)的日变化Fig.14 Daily change ofsensible and latent heat f ux of grassland during the growing season
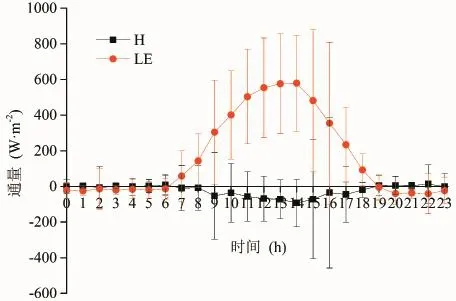
图15 生长季具鳞水柏枝灌丛显热通量(H)和潜热通量(LE)的日变化Fig.15 Daily change ofsensible and latent heat f ux of Myricaria squamosa Desv. shrubs during the growing season
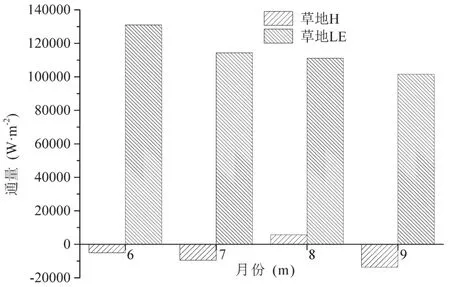
图16 生长季草地显热通量(H)和潜热通量(LE)的月值变化Fig.16 Monthy change ofsensible and latent heat f ux of grassland during the growing season
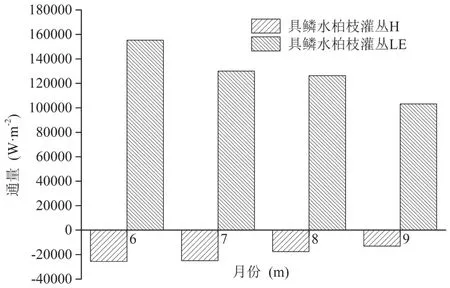
图17 生长季具鳞水柏枝灌丛显热通量(H)和潜热通量(LE)的月值变化Fig.17 Monthy change ofsensible and latent heat f ux of Myricaria squamosa Desv. shrubs during the growing season
3 结论
本文利用波文比系统和ECH2O土壤水分监测系统在青海湖流域沙柳河下游同一河道断面监测的两种典型湿地地植被群落类型——草地和具鳞水柏枝灌丛的空气温度、相对湿度、净辐射、土壤热通量和土壤温度等数据,对流域内两种湿地植被群落的群落小气候进行了研究,主要结论如下:
(1)受大气候的变化影响,草地和具鳞水柏枝灌丛的空气温度的日变化呈现出相同的变化趋势,其中最低值出现在6时左右,最高值出现在15—16时;在5月份草地和具鳞水柏枝灌丛的月平均气温值较为接近,而在6—9月份具鳞水柏枝灌丛的月平均气温值显著高于草地的月平均气温值,在整个生长季具鳞水柏枝灌丛的空气温度月平均值显著大于草地的空气温度月平均值;具鳞水柏枝灌丛林冠上方或群落上层的空气温度高于群落下层,而草地上下层没有显著差异。
(2)具鳞水柏枝灌丛和草地空气相对湿度日变化规律相同,白天较低,晚上较高,最高值出现在6时左右,最低值出现在15—16时;在整个植物的生长季,具鳞水柏枝灌丛和草地空气相对湿度差异显著,特别在6、7和9月份,具鳞水柏枝灌丛的空气相对湿度显著大于草地的空气相对湿度。
(3)具鳞水柏枝灌丛和草地群落冠层净辐射的日变化均呈单峰曲线,从7时至19时为正值,其余时段为负值,最大值出现在13时,从10时至17时具鳞水柏枝灌丛冠层的净辐射要大于草地冠层的净辐射;在5月份和6月份,具鳞水柏枝灌丛冠层与草地冠层的净辐射没有显著差异,而在7—9月份,具鳞水柏枝灌丛冠层与草地冠层的净辐射差异显著。
(4)具鳞水柏枝灌丛和草地地下5 cm土壤热通量的日变化呈单峰曲线,草地地下5 cm土壤热通量从10时至20时的时段内为正值,其余时段为负值;具鳞水柏枝灌丛地下5 cm土壤热通量从10时至18时的时段内为正值,其余时段为负值,草地地下5 cm土壤热通量的最大值出现在14时,为29.1 W·m−2,而具鳞水柏枝灌丛地下5 cm土壤热通量的最大值出现在13时,为33.2 W·m−2;具鳞水柏枝灌丛和草地的地下5 cm土壤热通量月变化趋势较为类似,在6月份为负,而在7、8、9月份为正;此外具鳞水柏枝灌丛的5 cm土壤热通量的变化幅度显著大于草地地下5 cm土壤热通量的变化幅度。
(5)在整个植被生长季,具鳞水柏枝灌丛和草地地下10 cm土壤温度日变化曲线呈相似的形状,草地地下10 cm土壤温度在18—19时达到最高值,之后逐渐下降,在次日9时出现最低值,具鳞水柏枝灌丛的最高值出现在15—16时,之后逐渐下降,在次日8时到达最低值,具鳞水柏枝灌丛的10 cm土壤温度对空气温度的响应更为敏感。具鳞水柏枝灌丛和草地的10 cm土壤温度月变化趋势基本一致,但是草地的10 cm土壤温度月均值大于具鳞水柏枝灌丛的10 cm土壤温度月均值。
(6)草地在白天和夜晚均由大气向地表输送热量,具鳞水柏枝灌丛在白天由大气向地表输送热量,而在晚上基本是由地表向大气输送热量;草地和具鳞水柏枝灌丛潜热通量的变化规律是7—18时为正值,其余时段为负值,说明白天是水汽蒸发的过程,而夜晚是水汽凝结的过程;草地和具鳞水柏枝灌丛的显热通量和潜热通量的标准偏差较大;草地和具鳞水柏枝灌丛在6—9月份的潜热通量皆为正值,且6月份的潜热通量最大,之后逐渐降低,而显热通量只有草地在8月份为正,其余月份全为负值,并且显热通量的绝对值显著小于潜热通量的绝对值,说明在地表与大气的能量交换中,潜热占主导,即地表主要通过潜热向大气输送热量,而显热的作用主要是将大气的热量输送到地表。
邓 艳, 蒋忠诚, 李先琨, 等. 2004. 广西弄岗不同演替阶段植被群落的小气候特征研究[J].热带地理, 24(4): 316 – 325. [Deng Y, Jiang Z C, Li X K, et al. 2004. AGE on microclimate in different successive vegetation community in Nonggang, Guangxi [J]. Tropical Geography, 24(4):316 – 325.]
董得红,王世红. 2006. 青海湖流域生态环境保护与治理对策[J].林业建设, (5): 16 – 19. [Dong D H, Wang S H. 2006. Environmental protection and rehabilitation strategies of Qinghai lake basin [J]. Forestry construction, (5): 16 – 19.]
冯宗炜, 冯兆忠. 2004. 青海湖流域主要生态环境问题及防治对策[J]. 生态环境, 13(4): 467 – 469. [Feng Z W, Feng Z Z. 2004.Major ecological and environmental problemsand countermeasures in the Qinghai Lake watershed, Qinghai [J]. Ecology and Environment, 13(4): 467 – 469.]
付为国, 李萍萍, 吴沿友,等. 2006. 镇江内江湿地不同演替阶段植物群落小气候日动态[J]. 应用生态学报, 17(9): 1699 – 1704. [Fu W G, Li P P, Wu Y Y, et al. 2006. Diurnal dynamics of microclimate at different successions stages of vegetation communities in inner-river of Zhenjiang City [J]. Chinese Journal of Applied Ecology, 17(9):1699 – 1704.]
贺芳芳. 2008. 上海市郊林带附近晴天地表热量平衡特征的分析[J].气象科学, 28(1): 37 – 44. [He F F. 2008. Characteristic analysis of surface heat balance of the near forest in f ne winter and summer days in Shanghai shrub [J]. Scientia Meteorologica Sinica, 28(1): 37 – 44.]
胡顺军,宋郁东,田长彦,等. 2006. 渭干河平原绿洲适宜规模[J].中国科学, 36(S1): 51 – 57. [Hu S J, Song Y D, Tian C Y, et al. 2006. Suitable scale of Weigan River plain oasis [J]. Science in China, 36 (S1): 51 – 57.]
李明财,罗天祥,郭 军,等. 2008. 藏东南高山林线冷杉原始林土壤热通量[J].山地学报, 26(4): 490 – 495. [Li M C, Luo T X, Guo J, et al. 2008. Soil heat f ux in Abies George Virgin Forest at the Alpine Timberline, Southeastern Tibetan Plateau [J]. Journal of mountain sciences, 26(4): 490 – 495.]
李英年,赵 亮,古 松,等. 2003. 海北高寒草甸地区能量平衡特征[J].草地学报, 11(4): 289 – 295. [Li Y N, Zhao L, Gu S, et al. 2003. Energy balance features of the Alpine Meadows on the Haibei Area (Northern Shores of the Qinghai Lake) [J]. Acta Agrestia Sinica, 11(4): 289 – 295.]
李宗峰, 陶建平, 王 微,等. 2005. 岷江上游退化植被不同恢复阶段群落小气候特征研[J]. 生态学杂志, 24(4): 364 – 367. [Li Z F, Tao J P, Wang W, et al. 2005. Community microclimate characteristics at different vegetation restoration stages of Upper Minjiang River [J]. Chinese Journal of Ecology, 24(4): 364 – 367.]
刘进琪, 王一博, 程慧艳. 2007. 青海湖区生态环境变化及其成因分析[J].干旱区资源与环境, 21(1): 32 – 37. [Liu J Q, Wang Y B, Cheng H Y. 2007. The eco-environmental change and causes of Qinghai lake watershed [J]. 21(1): 32 – 37.]
刘淑明, 孙丙寅, 王得祥. 2005. 不同地面覆盖对花椒冠层光能分布的影响[J]. 干旱地区农业研究, 23(6): 124 – 127. [Liu S M, Sun B Y, Wang D X. 2005. The influence of different covering on the radiation energy distribution of Zanthoxylum bungeanum [J]. Agricultural research in arid Areas, 23(6): 124 – 127.]
闵文彬. 2009. 丘陵区土壤热通量遥感估算模型适应性分析[J].气象科学, 29(3): 386 – 389. [Min W B. Analysis of adaptabilities of remote sensing models for heat flux over hilly land [J]. Scientia Meteorologica Sinica, 29(3): 386 – 389.]
申双和, 崔兆韵. 1999. 棉田土壤热通量的计算[J].气象科学, 19(3): 277 – 281. [Shen S H, Cui Z Y. 1999. Calculation of soil heat f ux in cotton f eld [J]. Meteorological science, 19(3): 277 – 281.]
司建华, 冯 起, 张小由, 等. 2005. 荒漠河岸林胡杨和柽柳群落小气候特征研究[J].中国沙漠, 25(5): 668 – 674. [Si J H, Feng Q, Zhang X Y, et al. 2005. Features of Microclimatein Populus euphratica and Tamarix ramosissima Communities in Desert Riparian Forest [J]. Journal of desert research, 25(5): 668 – 674.]
王 旭, 周国逸, 张德强,等. 2005. 南亚热带针阔混交林土壤热通量研究[J].生态环境, 14(2): 260 – 265. [Wang X, Zhou G Y, Zhang D Q, et al. 2005. Soil heat fluxes of mixed coniferous and broad-leaf forest in the south subtropics in China [J]. Ecology and Environment, 14(2): 260 – 265.]
吴红颜, 申 双, 徐为根. 2001. 棉田SPAS水热传输的多层模式[J].南京气象学院学报, 21(4): 137 – 142. [Wu H Y, Shen S, Xu W G. 2001. A multi-layer mode for water and heat transfer in soil-plant-atmosphere of cotton [J]. Journal of Nanjing Institute of Meteorology, 21(4): 137 – 142.]
颜江良, 吴 息, 江志红,等. 2008. 南京冬季城、郊下垫面能量平衡特征分析[J].气象科学, 28(1): 21 – 29. [Yan J L, Wu X, Jiang Z H, et al. 2008. Characteristic analysis of energy budget over urban and suburban underlying surface in Nanjing [J]. Scientia Meteorologica Sinica, 28(1): 21 – 29.]
杨 修, 孙 芳, 任 娜. 2003.环青海湖地区生态环境问题及其治理对策[J]. 地域研究与开发, 22(2): 39 – 42. [Yang X, Sun F, Ren N. 2003. The eco-environmental problems and rehabilitation strategies in the areas around Qinghai lake [J]. Areal research and development, 22(2): 39 – 42.]
张晓煜,王连喜,袁海燕. 2005.宁南半干旱地区农田和草地生态系统能量通量的季节变化[J].生态学报, 25(9):2333 – 2340. [Zhang X Y, Wang L X, Yuan H Y. 2005. Seasonal differences of energy fluxes among crop and grass ecosystems in semi-arid of southern Ningxia [J]. Acta Ecologica Sinica, 25(9): 2333 – 2340.]
张学元. 2006. 青海湖流域生态环境保护与治理对策[J].四川林勘设计, (3): 2 – 4, 23. [Zhang X Y. 2006. Ecological environment’s protecting and controlling countermeasures in Qinghai lake valley [J]. Sichuan Forestry Exploration and Design, (3): 2 – 4, 23.]
Al-Hamdani S H., Todd G W, Francko D A. 1990. Response of wheat growth and CO2assimilation to altering rootzone temperature [J]. Canadian Journal of Botany, 68(12): 2698 – 2702.
Bonan G B. 1989. Environmental factors and ecological processes controlling vegetation patterns in boreal forests [J]. Landscape Ecology, 3(2): 111 – 130.
Boone R D, Nadelhoffer K J, Canary J D, et al. 1998. Roots exert a strong inf uence on the temperature sensitivity of soil respiration [J]. Nature, 396: 570 – 572.
Dang Q L, Cheng S. 2004. Effect of soil temperature on ecophysiological traits in seedlings of four boreal tree species [J]. Forest Ecology and Management, 194(1 – 3): 379 – 387.
DeLucia E H, Day T A, Oquist G. 1991. The potential for photoinhibition of Pinussylvestris L. seedlings exposed to high light and low soil temperature [J]. Journal of Experimental Botany, 42: 611 – 617.
DeLucia E H. 1987. The effect of freezing nights on photosynthesis, stomatal conductance, and internal CO2concentration in seedlings of Engelmann spruce [J]. Plant Cell Environ, 10: 333 – 338.
Harper G J, Camm E L. 1993. Effects of frozen storage duration and soil temperature on the stomatal conductance and net photosynthesis of Piceaglauca seedlings [J]. Canadian journal of forest research, 23: 2459 – 2466.
Heithecker T D, Halpern C B. 2006. Variation in microclimate associated with dispersed-retention harvests in coniferous forests of western Washington [J]. Forest Ecology and Management, 226: 60 – 71.
Jerald A B, Kenneth C C. 2000. Estimating sensible heat flux from Oklahoma Mesonet [J]. Journal of Applied Meteorology and Climatology, 39:102 – 111.
Kaspar T C, Bland W L. 1992. Soil temperature and root growth [J]. Soil Science, 154: 290 – 299.
Landhäusser S. M, Wein, R W, Lange P. 1996. Gas exchange and growth of three arctic tree-line tree species under different soil temperature and drought pre conditioning regime [J]. Canadian Journal of Botany, 74: 1686 – 1693.
Lott J E, Ong C K, Black C R. 2009. Understorey microclimate and crop performance in a Grevillearobusta-based agroforestry system in semi-arid Kenya [J]. Agricultural and Forest Meteorology, 149: 1140 – 1151.
Mastrorilli M, Katerji N, Rana G, et al. 1998. ,Daily actual evapotranspiration measured with TDR technique in Mediterranean conditions [J]. Agricultural and Forest Meteorology, 90: 81 – 89.
McCafferty D J , Moncrieff J B , Taylor I R . 2003. Winter microclimate of field voles (Microtusagrestis) in SW Scotland [J]. Journal of Thermal Biology, 28: 397 – 401.
McMichael B L, Burke J B. 1996. Temperature effects on root growth [M]// Waisel et al. Plant Roots: The Hidden Half. New York: Marcel Dekker, Inc.
Nabi G, Mullins C E. 2008. Soil temperature dependent growth of cotton seedlings before emergence [J]. Pedosphere, 18(1): 54 – 59.
Rambo T R, North M P. 2009. Canopy microclimate response to pattern and density of thinning in a Sierra Nevada forest [J]. Forest Ecology and Management, 257: 435 – 442.
Savage M J, Everson C S, Metelerkamp B R. 2009. Bowen ratio evaporation measurement in a remote montane grassland: Data integrity and f uxes [J]. Journal of Hydrology, 376: 249 – 260.
Timlin D J, Pachepsky Y, Acock B A, et al. 2002. Error analysis of soil temperature simulations using measured and estimated hourly weather data with 2DSOIL [J]. Agricultural Systems, (72): 215 – 239.
Warrington I J, Kanemasu E T. 1983. Corn growth response to temperature and photoperiod. II. Leaf initiation and leaf appearance rates [J]. Agronomy Journal, 75: 755 – 761.
Zhang B Z, Kang S Z, Li F S,et al. 2010. Variation in vineyard evapotranspiration in an arid region of northwest China [J]. Agricultural Water Management, 97: 1898 – 1904.
Microclimate features of grassland communities and Myricaria squamosa Desv. shrubs in Shaliu River wetland, Qinghai Lake basin
LI Yue-tan1,2, LI Xiao-yan1
(1. State Key Laboratory of Earth Surface Processes, Beijing Normal University, Beijing 100875, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China)
Based on the microclimate data measured by Bowen-ratio system and soil water monitoring system (ECH2O), the microclimate features of the grassland and Myricaria squamosa Desv. shrubs on the same cross-section of Shaliu River were studied. Affected by macroclimate, the air temperature of grassland and Myricaria squamosa Desv. shrubs showed the same daily change trend, the mean monthly air temperature of Myricaria squamosa Desv. shrubs was significantly larger than that of grassland during the growing season, the air temperature on the top of Myricaria squamosa Desv. shrubs was higher than the bottom, while grassland showed no difference. The air relative humidity of grasslandand Myricaria squamosa Desv shrubs. which was higher during the daytime and lower during the night, also showed the same daily change trend, while it was signif cantly different between the two vegetation types during the growing season. The canopy net radiation daily change trend of the two vegetation types both showed the unimodal curve, but it was signif cantly different between the two vegetation types from July to September. The soil heat f ux change trend of the two vegetation types both showed the unimodal curve, and their monthly change trades were similar, but the changing amplitude of grassland was larger than Myricaria squamosa Desv. shrubs. The soil temperature daily change trend of grassland was similar to Myricaria squamosa Desv. shrubs, so did the monthly change trend, but the mean monthly value of grassland is larger than Myricaria squamosa Desv. shrubs. Energy transferred from ground surface to air on grassland both at daytime and night, but in Myricaria squamosa Desv. shrubs, energy transferred from air to ground surface at daytime and from ground surface to air at night. In the two wetland vegetation communities, evapotranspiration occured at daytime and water condensation occured at night, latent heat was the dominant way in the energy exchange between ground surface and air.
Qinghai Lake basin; Shaliu River wetland; Myricaria squamosa Desv. shrubs; grassland; microclimate
P463.2
:A
:1674-9901(2014)03-0173-13
10.7515/JEE201403001
2014-05-22
“十一五”国家科技支撑计划项目(2007BAC30B02);国家重点基础研究发展计划项目(2004CB720207);国家自然科学基金项目(NSFC40599423,Y3110391A1)
李岳坦,E-mail: liyuetan@sohu.com
