绿鳍马面鲀ERα基因部分cDNA序列克隆及其表达研究
2014-06-26赵梅琳温海深张冬茜李吉方
赵梅琳,温海深,张冬茜,何 峰,李吉方
(中国海洋大学 水产学院,山东 青岛 266003)
绿鳍马面鲀(Navodon septentrionalis)属鲀形目(Tetraodontiformes)革鲀科(Aluteridae),马面鲀属(Navodon),英文名 Bluefin leatherjacket,俗称面包鱼、象皮鱼、马面鱼等[1]。主要分布于日本、朝鲜半岛,非洲东岸,南非以及中国的东海、黄渤海和台湾海峡,其中东海是中国 绿鳍马面鲀产量最大的海区[2]。20世纪80年代以前,绿鳍马面鲀是中国海洋捕捞居第二位的重要经济鱼类[3]。近些年,由于自然海域资源严重衰退,加之肉质鲜嫩,肝脏营养价值高,绿鳍马面鲀的市场价格明显提高,绿鳍马面鲀冻鱼片、烤鱼片等诸多产品畅销国内外,市场前景十分广阔[1]。
雌激素参与调节生殖、骨骼以及脑的发育,是一种重要的性类固醇激素。研究表明,雌激素在体内的重要生理功能是由雌激素受体介导的,包括核受体介导的基因组效应和膜受体介导的非基因组效应[4-6]。雌激素受体(Estrogen receptors,ER)是核受体超家族成员之一,其蛋白的分子结构大致划分为六大功能结构域:A/B区为转录激活区、C区为DNA结合区、D区为铰链区、E区为配体结合区和F区[7]。雌激素受体在哺乳动物中有两个基因亚型,分别为雌激素受体α(ERα)和雌激素受体β(ERβ)。由于鱼类中发生了一个额外的基因复制事件,一些研究报道了上述两种亚型的变体。在虹鳟(Oncorhynchus mykiss)[8]、金鱼(Carassius auratus)[9]、倒刺 (Spinibarbus denticulatus)[10]等硬骨鱼类中均发现了两种 ERα亚型,而在斑马鱼(Danio rerio)[11]、尼罗罗非鱼(Oreochromis niloticus)[12]、许氏平鲉(Sebastes schlegeli)[13]等硬骨鱼类中均只发现一种 ERα。从目前的研究来看,不同种类的雌鱼、雄鱼的 ERα显示出不同的组织表达模式。在性成熟倒刺 和尼罗罗非鱼中,性腺、肝脏、肾脏和脑中均检测到 ERα的分布[10,12]: 金鱼中,垂体的表达量最高,性腺、肝脏、肠、脑中均有较高的表达[9]: 许氏平 鲉中,ERα在雌鱼中有丰富的表达,在卵巢、肝脏、肠、脂肪、肾脏、脑、心、胃、脾、头肾中均有较高表达,而雄鱼中只在精巢和肝脏检测到较高表达[13]。目前,国内外对绿鳍马面鲀分子生物学的研究甚少,关于其雌激素受体的研究尚未见报道。
作者克隆了绿鳍马面鲀ERα基因核心序列,采用半定量方法分别检测了其在雌鱼、雄鱼多个组织中的表达情况,并采用实时定量RT-PCR方法分析其在卵巢和精巢组织中的周年表达情况。该项研究为进一步阐明绿鳍马面鲀ERα的生理功能提供了科学依据,同时也为其人工繁育提供一定的理论基础。
1 材料与方法
1.1 样品采集
2010年11月至2011年9月,从烟台百佳水产有限公司每2个月采集1次,每次采样用于本实验的有3尾雄鱼和3尾雌鱼,共36尾。实验鱼规格为体长(20.0~29.5)cm,体质量(195.5~648.8)g。实验鱼麻醉后解剖,收集各组织样品,于–80℃超低温冰箱保存用于基因克隆和表达,并测定各生物学指标,包括体质量、体长、去内脏质量、性腺质量等。计算性腺指数(GSI=[性腺质量/去内脏质量]×100)。
1.2 总RNA的提取和cDNA的合成
绿鳍马面鲀各组织总 RNA用 RNAiso reagent(Takara)按照说明书进行提取。分用UV分光光度法(Ultrospec2100Pro,Amersham)和琼脂糖凝胶电泳进行浓度和完整性检测。各样本的1 μL总RNA用随机引物和反转录酶M-MLV(Takara)合成第一链。
1.3 ERα的克隆及序列分析
根据已知鱼类的ERα序列,利用CodeHop在线[14]设计简并引物,如表1,并以卵巢组织的cDNA为模板进行扩增。所得序列用 DNAMAN软件推算氨基酸序列,并在 NCBI数据库(http: //www.ncbi.nlm.nih.gov/)中进行BLAST比对和同源性分析。

表1 绿鳍马面鲀ERα cDNA克隆及表达所用引物Tab.1 Primers used for Navodon septentrionalis ERα cDNA cloning and mRNA expression analysis
1.4 系统进化树构建
使用Clustal X将其与其他物种ERα氨基酸序列进行多重序列比对,使用 MEGA 4.0邻接法(Neighbour-Joining,NJ)构建系统进化树[15]。
1.5 ERα的组织表达和周年表达
利用 Primer5.0软件设计一对特异性引物tsERαF5和tsERαR5。通过半定量RT-PCR分析ERα在雌鱼和雄鱼的各组织分布,包括性腺、肌肉、肠、鳃、肾、胃、脾脏、肝脏、心脏、垂体和脑。PCR反应条件:94℃预变性5 min,94℃ 30 s,57℃ 30 s,72℃ 30 s共35个循环,72℃ 10 min,4℃ 1 h。18 S rRNA作为内参基因,反应条件:94℃预变性5 min,94℃ 30 s,57℃ 30 s,72℃ 30 s共25个循环,72℃10 min,4℃ 1 h。PCR产物进行1.5%琼脂糖凝胶电泳,并用GIS凝胶成像系统3.60软件(Tanon)分析扩增条带的光密度。
设计特异性引物tsERαF4和tsERαR4,运用实时定量qRT-PCR技术检测ERα在精巢、卵巢(n=3)中的周年表达情况,并以18S为内参基因对数据进行分析[16]。
1.6 数据处理
实验数据用平均数±标准误表示,并采用 SPSS 13.0软件进行 ANOVA分析,用 Duncan’s多重比较进行数据处理和检验分析,当P<0.05时差异显著。
2 结果
2.1 绿鳍马面鲀雌激素受体ERα cDNA核心序列的克隆
克隆得到绿鳍马面鲀雌激素受体ERα cDNA核心序列共1410 bp,按照DNAMAN V6的翻译纵览确定该序列的翻译起始位点并进行翻译,该序列编码470个氨基酸,如图1所示。该序列已提交GenBank,登录号为 JX508794。在绿鳍马面鲀中,只克隆得到了一种型式的ERα,尚没有发现其他亚型。
2.2 ERα基因序列分析与系统进化分析
推导得到的氨基酸序列与其他鱼的ERα蛋白序列比对,该序列与其他核受体相似,可以分为6个功能区,如图 2所示。其中 C区和 E区比较保守。C区有由 8个保守的半胱氨酸残基组成的两个锌指结构,并具有高度保守性的 D-box(PATNQ)、P-box(CEGCKA)以及PKA位点(RRKS)。E区有一个配体依赖的转录激活区(AF-2),它在硬骨鱼和哺乳动物中都是高度保守的。

图1 绿鳍马面鲀ERα cDNA部分序列Fig.1 The partial sequence of Navodon septentrionalis ERα cDNA
用Clustal X软件将推导得到的氨基酸序列与人(Homo sapiens)、褐家鼠(Rattus norvegicus)、非洲爪蟾(Xenopu stropicalis)、斑马鱼等 13个物种的 ERα序列进行比对。图 3显示,硬骨鱼类汇为一个分支,哺乳动物和两栖类汇为另外一 个分枝。绿鳍马面鲀ERα与黑棘鲷(Acanthopagrus schlegelii)ERα的进化地位最为接近,Blastp比对,两者的相似度为89%。此外,绿鳍马面鲀ERα与舌齿鲈(Dicentrarchus labrax)、许氏平 鲉、点带石斑鱼(Epinephelu scoioides)等的进化地位较为接近,其氨基酸相似度分别为88%、87%、86%。与倒刺、金鱼等亲缘关系较远。
2.3 ERα的组织分布
绿鳍马面鲀ERα在雌鱼、雄鱼的11种组织间的表达模式不同。从图4中可以看出,雌鱼中,ERα在垂体、心脏、卵巢中的表达量最高,其次为脑、肝脏、胃、肾、肌肉。在脾脏、肠中的表达最较低,而在鳃中没有检测到表达: 雄鱼中,ERα在垂体、肝脏、肾脏、精巢中的表达量相对较高,其次为脑和肠,在胃、脾脏、心脏、鳃、肌肉中没有检测到表达。
2.4 不同性腺发育期中ERα的周年变化
绿鳍马面鲀的 GSI随着性腺发育呈明显的周年变化规律(图 5A),雌鱼 GSI在 11月份出现最低值2.15,之后缓慢上升,在翌年5月显著升高并达到全年最大值6.30,之后逐渐下降,7月份出现一次显著性降低4.47(P<0.05): 雄鱼GSI与雌鱼呈相似的变化规律(图5B),11月出现最低点,为1.30,最高峰同样出现在5月,为4.73。
绿鳍马面鲀ERα在卵巢、精巢间呈现不同的周年表达模式(图6A),卵巢中ERα相对mRNA水平在5月份有最大值1.60,7月份显著降低,并出现最低值0.72(P<0.05),之后开始回升。而在精巢中,9月份出现最大值,为1.57,之后缓慢下降,在5月份显著降低,并达到最低值0.71(P<0.05,图6B)。
3 讨论
本试验 克隆得到绿鳍马面鲀ERα的基因核心cDNA片段共 1410bp,其所推导的氨基酸序列与黑棘鲷同源性最高(89%),且氨基酸序列含有核受体超家族典型的特征:分为6个功能区,C区和E区高度保守,且E区包含AF-2活性功能区。表明该序列为目的序列。

图2 绿鳍马面鲀ERα与其他物种ERα氨基酸序列比较Fig.2 Comparison of amino acid sequences of Navodon septentrionalis ERα with other species
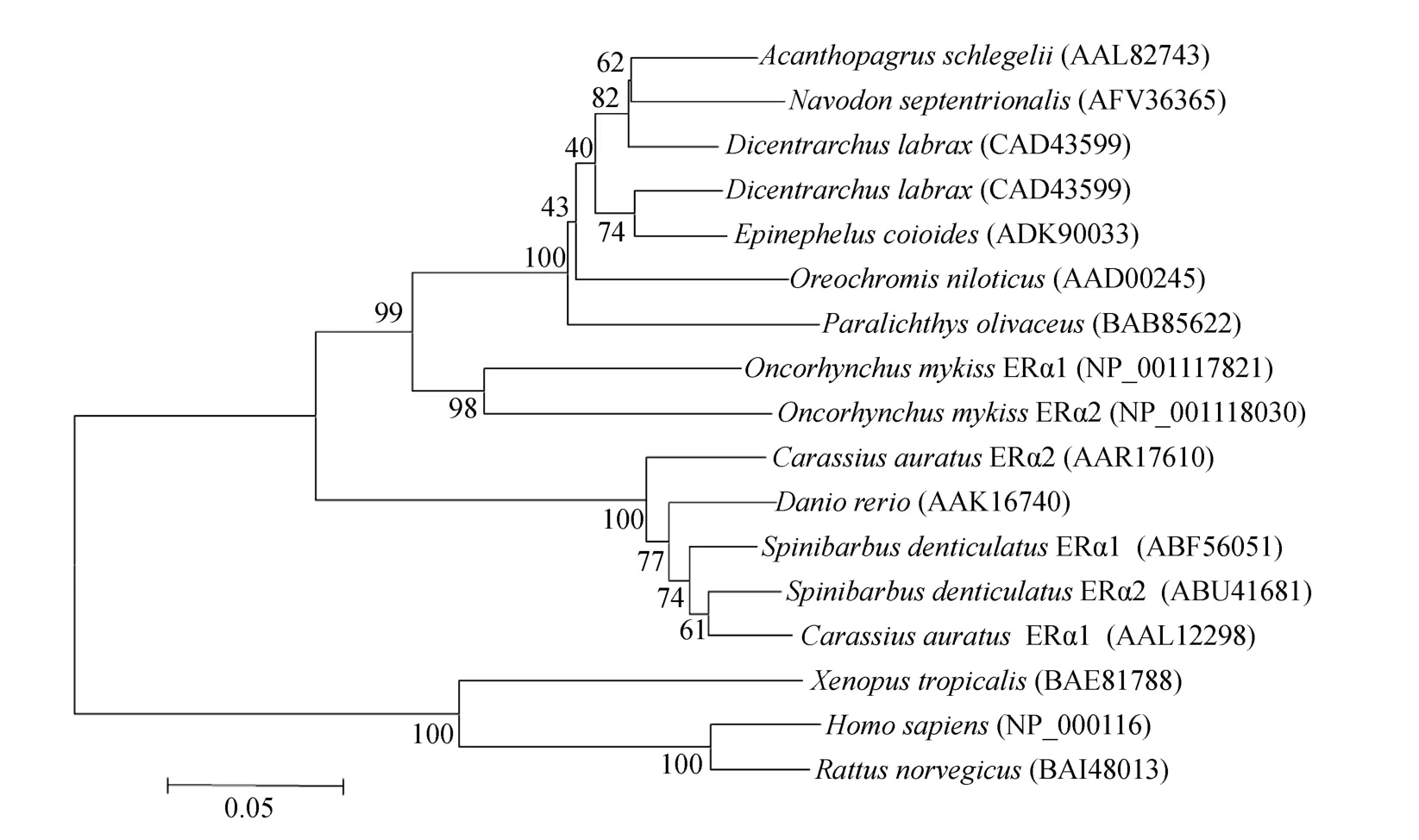
图3 不同物种的ERα氨基酸构建的系统进化树Fig.3 Phylogeneticanalysis of the amino acid sequences of ERα

图4 ERα 在绿鳍马面鲀雌鱼、雄鱼的组织表达Fig.4 Tissue distribution of ERα in Navodon septentrionalis by RT-PCR
雌激素受体在生物体内广泛表达,其研究具有重要的生理意义。随着20世纪80年代首次成功克隆得到人 ERα受体开始[17],其在哺乳动物中的研究逐渐兴起。近年来雌激素受体在硬骨鱼中的研究较多。据报道,金鱼ERα在垂体中表达信号最强,其他组织次之[9]: 斑马鱼ERα在垂体、肝脏、精巢中表达量较高[11]: 金头鲷(Sparus auratus)成鱼 ERα主要在肝脏和垂体中表达[18]。本研究与上述研究结果相似,绿鳍马面鲀 ERα在垂体、肝脏和性腺中也有较高的表达。已有研究显示,雌激素在中枢神经组织中与其受体结合,对生殖行为、神经保护、学习记忆等有重要作用[19]:雌激素在性腺和肝脏中与雌激素受体结合,分别调控卵黄蛋白原合成相关基因的表达,进而影响性腺发育,调节生殖行为[10,20,21]。
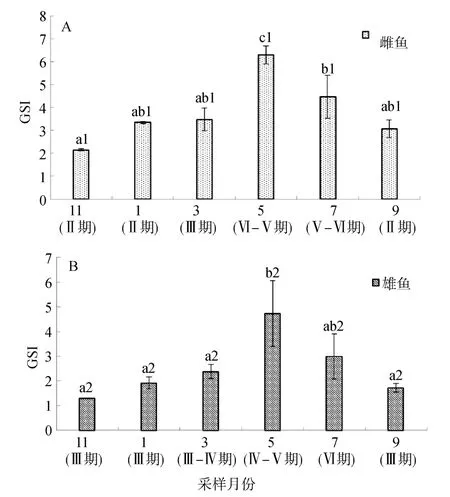
图5 绿鳍马面鲀雌鱼、雄鱼的GSI周年变化Fig.5 The annual changes of GSI in female and male Navodon septentrionalis

图6 ERα在精巢和卵巢中的周年表达模式Fig.6 The relative expression pattern of ERα in ovary and testis of Navodon septentrionalis during the annual reproductive cycle
绿鳍马面鲀为周年产卵鱼类,一般从 4月中下旬到 6月底为繁殖期[22],人工养殖的雌性绿鳍马面鲀,其生殖周期与野生绿鳍马面鲀生殖周期基本一致。绿鳍马面鲀雌鱼GSI在11月有最低值,可能是由于鱼体要大量储存能量用以越冬,只有少量用以性腺发育所致,与人工养殖半滑舌鳎(Cynoglossus semilaevis)进入12月份后GSI显著下降相一致[22]。从11月到翌年3月,随着卵巢发育,卵黄积累,雌鱼的GSI逐渐升高,5月开始进入繁殖期,此时GSI显著高于 11月(P<0.05)。与此同时,卵巢 ERα表达水平也逐渐上升并在 5月达到峰值,推测可能由于卵巢发育后期,卵黄大量积累,必须有大量雌激素参与卵母细胞的发育,此时雄激素大量转化为雌激素所致[23],已有研究证实,雌激素水平的升高与卵黄生成有关[24]。之后 GSI逐渐下降,由于卵巢完成排卵并开始退化吸收,在7月份GSI出现一次显著性降低(P<0.05),此时卵巢ERα达到全年最低。9月卵巢完成退化吸收进入重复发育Ⅱ期,GSI有所降低,卵巢ERα 表达量开始回升。绿鳍马面鲀雌鱼GSI的这一变化规律与黄鲷(Dentextu mifron)[25]、半滑舌鳎[22]等硬骨鱼类的研究结果一致。Shi等[13]的研究结果显示,许氏平 鲉ERα在卵巢发育成熟时有最高水平,其他时期均保持在较低水平,与本研究中绿鳍马面鲀卵巢ERα变化趋势相似。
绿鳍马面鲀雄鱼GSI变化趋势与雌鱼基本相同。其精巢ERα表达水平与GSI没有直接的相关性。雄鱼GSI在11月出现最低值。随着精原细胞的分裂分化,精母细胞的逐渐成熟,翌年5月进入繁殖期并达到峰值,此时精巢ERα呈下降的趋势,并在5月达到最低,可能是由于随着精巢发育成熟雌激素含量减少所致。之后随着GSI的降低,精巢ERα水平上升。据报道,鳗鲡(Anguilla japonica)、倒刺、许氏平鲉随着精子生成的全部过程,精巢 ERα表达水平逐渐升高,可以认为雌激素在雄鱼精子发生中是不可或缺的,但其作用机制尚不明确[10,13,26],而许氏平鲉ERα的表达水平是否与其卵胎生这一特殊的繁殖方式有关尚不十分清楚,同时也不排除对部分鱼类特定发育期,精巢雌激素受体表达研究结果有互相矛盾的现象[27]。雌激素受体在雄鱼中的机理尚不完全了解,有待于进一步研究。
作者首次克隆绿鳍马面鲀ERα核心cDNA序列,并对其进行了基因结构与进化分析,分别检测了雌鱼、雄鱼的组织表达情况,同时还研究了 ERα在绿鳍马面鲀生殖周期中的表达,为进一步揭示 ERα的生理功能提供科学依据,同时也为绿鳍马面鲀的人工繁育提供了一定的理论基础。
[1]孟宪菊,张利民.绿鳍马面鲀的生物学特性及人工养殖前景[J].齐鲁渔业,2009,26(6): 47-48.
[2]李平伦,李德军,徐金波,等.绿鳍马面鲀海上网箱养殖技术要点[J].齐鲁渔业,2003,20(10): 10-11.
[3]陈莲芳,郑元甲.东海绿鳍马面鲀的早期发育和产卵场、产卵期的探讨[J].生态学报,1984,4(2): 73-79.
[4]Edwards D P.Regulation of signal transduction pathways by estrogen and progesterone[J].Annual Review of Physiology,2005,67: 335-376.
[5]Levin E R.Plasma membrane estrogen receptors[J].Trends in Endocrinology &Metabolism,2009,20(10):477-482.
[6]Thomas P,Dressing G,Pang Y,et al.Progestin,estrogen and androgen G-protein coupled receptors in fish gonads[J].Steroid,2006,71(4): 310-316.
[7]Krust A,Green S,Argos P,et al.The chicken oestrogen receptor sequence: homology with v-erbA and the human oestrogen and glucocorticoid receptors[J].The EMBO Journal,1986,5: 891–897.
[8]Nagler J J,Krisfalusi M,Cyr D G.Quantification of rainbow trout(Oncorhynchus mykiss)estrogen receptora messenger RNA and its expression in the ovary during the reproductive cycle[J].Molecular Endocrinology,2000,25: 243-251.
[9]Choi C Y,Habibi H R.Molecular cloning of estrogen receptor a and expression pattern of estrogen receptor subtypes in male and female goldfish[J].Molecular and Cellular Endocrinology,2003,204: 169-177.
[10]Zhu P,Zhang Y,Zhou Q,et al.Discovery of four estrogen receptors and their expression profiles during testis recrudescence in male Spinibarbus denticulatus[J].General and Comparative Endocrinology,2008,156: 265-276.
[11]Menuet A,Pellegrini E,Anglade I,et al.Molecular characterization of three estrogen receptor forms in zebrafish: binding characteristics,transactivation properties,and tissue distributions[J].Biology of Reproduction,2002,66(6): 1881-1892.
[12]Wang D S,Senthilkumaran B,Sudhakumari C C,et al.Molecular cloning,gene expression and characterization of the third estrogen receptor of the Nile tilapia,Oreochromis niloticus[J].Fish Physiology and Biochemistry,2005,31(2-3): 255-266.
[13]Shi D,Wen H S,He F,et al.The physiology functions of estrogen receptor a (ER a)in reproduction cycle of ovoviviparous black rockfish,Sebastes schlegeli Hilgendorf[J].Steroids,2011,76: 1597-1608.
[14]陈彩芳,温海深,何峰,等.程序化设计的简并引物克隆半滑舌鳎 CYP17基因[J].中国海洋大学学报,2009,39(6): 1213-1218.
[15]Kumar S,Tamura K,Nei M.MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment[J].Briefings in bioinformatics,2004,5(2): 150-163.
[16]Mu W J,Wen H S,He F,et al.Cloning and expression analysis of the cytochrome P450c17senzymes during the reproductive cycle in ovoviviparous Korean rockfish(Sebastes schlegeli)[J].Gene,2013,512: 444-449.
[17]Walter P,Green S,Greene G,et al.Cloning of the human estrogen receptor cDNA[J].Proceeding of the National Academy of Sciences of the USA,1985,82(23): 7889-7893.
[18]Pinto P I,Passos A L,Martins R S,et a1.Characterization of estrogen receptor b in sea bream (Sparus auratus): phylogeny,ligand-binding,and comparative analysis of expression[J].General and Comparative Endocrinology,2006,145: 197-207.
[19]Mcewen B S,Alves S E.Estrogen action in the central nervous system[J].Endocrine Reviews,1999,20(3):279-307.
[20]Campbell C M,Idler D R.Characterization of an estradiol-induced protein from rainbow trout as vitellogenin by the composition and radioimmunological cross reactivity to ovarian yolk fractions[J].Biology of Reproduction,1980,22(3): 605-617.
[21]Sabo A T ,KrollK J,Denslow N D.Differential expression of largemouth bass(Micropterus salmoides)estrogen receptor isotypes alpha,beta,and gamma by estradion[J].Molecular and Cellular Endocrinology,2004,218(1-2): 107-118.
[22]陈彩芳,温海深,陈晓燕,等.人工养殖半滑舌鳎卵巢发育及其产卵类型研究[J].海洋科学,2010,34(8):29-34.
[23]方永强,翁幼竹,胡晓霞.性类固醇激素及其受体在文昌鱼性腺和神经系统中的分布[J].动物学报,2001,47(4): 398-403.
[24]Lubzens E,Young G,Bobe J,et al.Oogenesis in teleosts: how fish eggs are formed [J].General and Comparative Endocrinology,2010,165(3): 367-389.
[25]施兆鸿,王建钢,高露姣,等.南海黄鲷性腺发育的初步研究[J].台湾海峡,2006,25(3): 353-359.
[26]Miura T,Miura C,Ohta T,et al.Estradiol-17β stimulates the renewal of spermatogonial stem cells in males[J].Biochemical and Biophysical Research Communications,1999,264: 230-234.
[27]温海深,董双林.硬骨鱼类雌激素受体及其在生殖调节中的作用研究[J].中国海洋大学学报,2008,38(3):367-370.
