低氧诱导因子-3α的研究进展
2014-05-19李文斌贾正平张娟红
鹿 辉,尹 强,王 荣,谢 华,李文斌,贾正平,张娟红
(兰州军区兰州总医院全军高原损伤防治重点实验室,甘肃兰州 730050)
低氧诱导因子(hypoxia-inducible transcription factor,HIF)最早是由Semenza等[1]于1992年在研究促红细胞生成素基因时发现的。HIF是细胞应对氧气水平降低时的主要调节因子,调控依赖氧气水平相关基因的表达[2]。这些基因主要参与造血、血管生成、离子转运、葡萄糖的利用、细胞外基质的合成、细胞的增殖、生存及凋亡和肿瘤的发生等过程[3-4]。HIF是由一个不稳定的 α亚基(HIF-α)和一个稳定的β亚基(HIF-β)组成的二聚体,具有结合靶基因的低氧反应元件(hypoxia response elements,HREs)的能力,从而发挥调控作用。HIF-α和HIF-β亚基均属于 bHLH-PAS(bHLH,basic helix-loop-helix;PAS,Per-ARNT-Sim;Per,period circadian protein;ARNT,aryl hydrocarbon receptor nuclear translocator protein;Sim,single-minded protein)蛋白家族的成员[5]。序列同源性HIF-α亚基在人体中目前认为有3种:HIF-1α(由基因 HIF1A编码)、HIF-2α(由基因 EPAS1编码)和HIF-3α(由基因HIF3A编码HIF-3α的多种剪接变异体)[6],三者均受氧浓度调节,是调节HIF活性的功能亚单位[8]。。相对于HIF-1α和HIF-2α,对HIF-3α的认识和研究还比较匮乏。HIF-3α存在多种剪切变异体,其功能也不尽相同,调控包括低氧应激在内的多种生理活动。本综述将系统地介绍HIF-3α的结构、表达调控及其在多种生理活动中的调控作用。
1 HIF-3α的结构
HIF-1α和HIF-2α亚基具有相似的结构域,均具有1个N端bHLH结构域,2个PAS结构域,氧气依赖降解结构域(oxygen-dependent degradation domain,ODD)以及 N端和 C端的转录激活结构域(transactivation domain,TAD)组成,N端的TAD结构域的部分序列与ODD重叠[9]。1998年,在老鼠体内首次发现了HIF-α的第3种亚型HIF-3α[10],在人体内于2001年首次发现HIF-3α[11]。这种新的蛋白与HIF-1α和HIF-2α的bHLH-PAS区域有55%的相似性[10]。与HIF-1α和HIF-2α相似,HIF-3α中也含有1个ODD结构域和1个N-TAD结构域,但没有C-TAD结构域(Fig 1)。
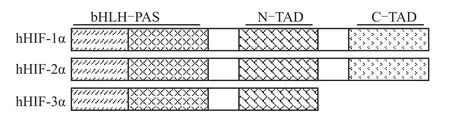
Fig 1 Structural alignment of human HIF-1α,HIF-2α and HIF-3αsubunits
2 人类 HIF-3α(hHIF-3α)的剪接变异体
人类HIF-3α(hHIF-3α)有8种剪接变异体,其结构域见Fig 2,与鼠 HIF-3α(mHIF-3α)相似,人类 HIF-3α基因包含19个内含子,长度大约是43 kb的碱基长度,位于染色体的19q13.2的位置。3个特别的外显子1a、1b和1c很可能含有8种剪接变异体的转录起始位点。外显子2编码bHLH结构域,外显子3-9编码PASa、PASb和PAC(PAS-associated C-terminal domain)结构域,外显子11的3′末端部分、整个的外显子12和外显子13的5′末端部分编码ODD结构域,外显子14a和16编码亮氨酸拉链结构域(leucinezipper,LZIP)[6]。
hHIF-3α1的序列从外显子1c开始,到外显子16终止。hHIF-3α1编码668个氨基酸长度的蛋白质,含有N-TAD结构域,但并不含有C-TAD结构域。有趣的是,与HIF-1α、HIF-2α及其他hHIF-3α的变异体不同的是,hHIF-3α1含有独特的亮氨酸拉链(leucine zipper,LZIP)结构域。LZIP结构具有介导与DNA结合和蛋白质-蛋白质相互作用的功能。hHIF-3α1的另外一个特殊的结构是,在ODD和LZIP结构域的上游具有LXXLL(Leu-Xaa-Xaa-Leu-Leu)基序。LXXLL基序主要在核受体的共因子中发现,是蛋白质相互作用的位点。LZIP和LXXLL结构在HIF-1α和HIF-2α中都不存在,表明hHIF-3α1可能具有结合启动子序列的能力或者结合不被HIF-1α和HIF-2α识别的新蛋白质的能力。hHIF-3α2从外显子1a开始,终止于外显子13a,不包含外显子1b和1c,编码632个氨基酸长度的蛋白质。hHIF-3α2的结构与hHIF-3α1非常相似,但缺少LZIP结构域及其上游的LXXLL基序。hHIF-3α4从外显子1a开始,终止于不完整的内含子8,内含子7没有被剪切掉。编码363个氨基酸长度的蛋白质,只含有 bHLH、PASa和 PASb结构域,缺少 N-TAD、CTAD、ODD和LZIP结构域及LXXLL基序,表明hHIF-3α4可能作为hHIF调控系统的负调控因子。hHIF-3α6从外显子1b开始,缺少外显子3和hHIF3α4相似,含有内含子7并终止于内含子8,hHIF-3α6蛋白只包含 C末端的结构域PASb。hHIF-3α7和 hHIF-3α8结构相似,但 hHIF-3α7缺少LZIP结构域。hHIF-3α9和 hHIF-3α1结构相似,差别仅在于外显子1的不同。值得注意的是hHIF-3α10,其编码序列被内含子1所阻断,因此仅编码7个氨基酸[6-7]。
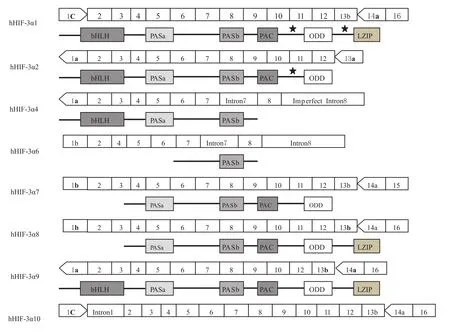
Fig 2 Multiple spliced variants of the human HIF-3α,Asterisk represents the conserved LXXLL motif
3 HIF-3α的基因表达
急性缺氧(12%O2,2 h)对 HIF-1α、HIF-2α和 HIF-1β的mRNA水平在心、肺、肾和骨骼肌组织中的表达没有明显的影响,但在上述组织中却强烈地诱导HIF-3α的mRNA表达。虽然5个星期的间歇性缺氧训练(intermittent hypoxic training,IHT)可以诱导HIF-3α的mRNA在上述组织的上调表达,但是IHT后的急性缺氧除了在骨骼肌组织可以诱导HIF-3α的mRNA表达上调外,在其他组织中HIF-3α的mRNA都没有被诱导。这说明IHT抑制了急性缺氧在心、肺、肾组织中对 HIF-3α的mRNA的诱导作用[12]。hHIF-3α4被认为是hHIF调控系统的负调控因子。在低氧的刺激下,HIF调控的基因将会被激活从而应对由缺氧造成的细胞损伤,作为HIF系统的负调控因子hHIF-3α4将会是被抑制的。在缺氧4 h的条件下,利用实时定量PCR测定HepG2肝癌细胞和HEK293A人胚肾细胞中内源性的hHIF-3α4的表达水平,与对照组相比分别降低了75%和50%[13]。这说明hHIF-3α4作为HIF系统的负调控因子,在低氧状态下是被抑制的,从而使细胞能够表达氧应激相关基因来应对低氧应激。大鼠mHIF-3α(mHIF-3α)的mRNA水平可在急性缺氧的情况被诱导表达。
令人感到兴奋的是,mHIF-3α除了被低氧诱导外,还可以被其他因素诱导。文献报道[14],mHIF-3α可以由胰岛素和2-脱氧-D-葡萄糖诱导表达。这提示,mHIF-3α除具有参与低氧应激的功能外,还可能具有其他方面的功能,例如可能在糖代谢的调节方面发挥重要的作用。
4 HIF3α4的负调控作用
HIF-3α4缺少N-TAD、C-TAD和 ODDD结构域,因此被认为是HIF系统的负调控因子。文献报道,HIF-3α4可以与HIF-1α结合,从而降低了 HIF1复合物的水平,对hHIF-1α起负调控的作用。HIF-3α4可以抑制HIF-1α/HIF-β复合物与靶基因HREs序列区的结合,从而抑制了HIF-1α调控的相关基因的表达,起到负调控的作用[13]。利用HEK293A研究发现,HIF-3α4可以与HIF-2α相结合,从而降低HIF2复合物的量,降低HIF-2α的调节作用。研究表明,HIF-3α4与HIF-2α相结合后将明显降低HIF-2α与靶基因HREs序列区的结合,从而抑制了相关基因的表达,起到负调控的作用[15]。HIF-3α4也可以与 HIF-β相结合,可以同时减少体内HIF1和HIF2复合物的量,对HIF-1α和HIF-2α同时起到负调控作用[13,15]。
5 HIF-3α在肺发育过程中的作用
肺的发育是在相对缺氧的条件下进行的,HIF信号通路在肺的发育过程中起了重要的作用。基因HIF3α/NEPAS/IPAS的缺失会导致刚出生小鼠肺重塑功能的缺损[16]。在小鼠肺上皮细胞中表达HIF-3α会导致后期支气管形态的缺陷、肺泡的数量降低和改变上皮细胞类型的分化。实验表明,HIF-3α基因的表达,会使Clara细胞、肺泡Ⅰ型和Ⅱ型细胞的数量有所减少,也使基底细胞处在非经典的空间位置。HIF-3α在小鼠的肺的发育过程中发挥作用,主要的机制是选择性地抑制了HIF-2α基因的表达,并上调Sox2、Rarβ和Foxp2基因的表达[17]。Sox2基因可以直接影响基底细胞的外形[18],Rarβ基因敲除小鼠表现为肺泡提前分隔,肺泡数量也是正常小鼠的两倍。所以,HIF-3α上调Rarβ基因,从而抑制了小鼠的肺在发育过程中肺泡数量的增加。Foxp2基因具有抑制Ccsp和远端上皮细胞标记(Spc,T1α)的表达作用[19]。这解释了HIF-3α表达后,Clara细胞、肺泡Ⅰ型和Ⅱ型细胞数量减少的原因。
6 HIF-3α在脂肪细胞分化中的作用
HIF-3α除了在肺的发育过程中发挥重要的作用之外,在其他组织的发育过程中也具有重要的作用。HIF-3α在3T3-L1细胞的脂肪细胞分化过程中被诱导表达,说明可能是脂肪细胞分化的促进因子,在脂肪细胞的分化过程中发挥重要的作用。基因芯片分析实验表明,当HIF-3α过量表达时,两个已知的脂肪细胞分化相关基因,aP2和AMPKγ1表达是上调的。脂肪细胞分化相关基因的表达上调,可能是由于HIF-3α的LZIP结构域能够激活脂肪细胞相关基因的表达造成的[20]。
7 HIF-3α在增强耐力中的负调控作用
抑制HIF-3α的mRNA水平将明显提高大鼠的身体耐力。除了IHT可以抑制HIF-3α的mRNA水平外,耐力训练(endurance training,ET)也可以抑制 HIF-3α的 mRNA水平。实验表明大鼠的HIF-3α的mRNA水平在ET联合IHT时只有空白对照组的15%,只进行ET时为对照组的30%。IHT后的ET训练,大鼠表现出最高的身体耐力水平。利用RNA干扰技术,在大鼠的比目鱼肌和腓肠肌中抑制HIF-3α的表达,可以明显的提高大鼠的身体耐力。这说明HIF-3α在大鼠身体耐力增强中具有重要的负调控作用[12]。
8 HIF-3α4在脑膜瘤中的作用
脑膜瘤是血管最丰富的脑瘤之一,有效抑制脑膜瘤的血管生成是一种有效的新疗法。HIF-1α和HIF-2α在肿瘤的血管生成中具有重要的作用,而HIF-3α4被认为是上述两个α亚基的负调控因子。最近的研究表明,在脑膜瘤中HIF-3α4的表达是受到抑制的,这是由于 HIF-3α4的启动子区DNA的甲基化导致的。在富血管脑膜瘤中诱导HIF-3α4高表达,可以有效地抑制脑膜瘤的血管生成、细胞分化和物质代谢,显著地阻滞脑膜瘤的生长。因此,HIF-3α4在脑膜瘤中发挥重要的作用,是潜在的治疗脑膜瘤的潜在分子靶点[21]。
9 总结
HIF是细胞应对氧气水平降低时的主要调节因子,HIF-1α、HIF-2α和HIF-3α是HIF的3种活性亚基。近年来,对前两种活性亚基,特别是HIF-1α进行了大量的研究工作,对这两个亚基的基因分布及表达调控机制、激活和降解机制以及参与生理活动的机制都有深入的了解,相比前两个亚基而言,对HIF-3α的研究和认识还比较有限。通过最近几年的研究表明,人体的HIF-3α具有6种剪切变异体,每一种的结构都不一样,其发挥的功能也可能不一样。值得注意的是,hHIF-3α4是负调控因子,对HIF信号通路系统起负调控作用,使其与其他亚基相比具有独特性。研究还提示,HIF-3α呈现出功能的多样性,除了参与低氧应激的反应外,还参加了体内的其他生理过程,这也是与HIF-1α和HIF-2α不同的。例如HIF-3α参与组织和器官的分化和发育,以及糖代谢的过程等。由目前的实验结果,我们认识到HIF-3α的功能是复杂的,具有独特性。因此,深入研究HIF-3α的功能及其作用机制是及其必要的,将会为肿瘤和高原疾病的治疗提供新的思路和策略。
参考文献:
[1]Semenza G L,Wang G L.A nuclear factor induced by hypoxia via de novo protein synthesis binds to the human erythropoietin gene enhancer at a site required for transcriptional activation[J].Mol Cell Biol,1992,12(12):5447-54.
[2]Kaelin W G Jr,Ratcliffe P J.Oxygen sensing by metazoans:the central role of the HIF hydroxylase pathway[J].Mol Cell,2008,30(4):393-402.
[3]Weidemann A,Johnson R S.Biology of HIF-1alpha[J].Cell Death Differ,2008,15(4):621-7.
[4]Semenza GL.HIF-1:upstream and downstream of cancer metabolism[J].Curr Opin Genet,2010,20(1):51-6.
[5]Wang G L,Jiang B H,Rue E A,Semenza G L.Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PASheterodimer regulated by cellular O2tension[J].Proc Natl Acad Sci USA,1995,92(12):5510-4.
[6]Pasanen A,Heikkil M,Rautavuoma K,et al.Hypoxia-inducible factor(HIF)-3αis subject to extensive alternative splicing in human tissues and cancer cells and is regulated by HIF-1 but not HIF-2[J].Int J Biochem Cell Biol,2010,42:1189-200.
[7]Heikkila M,Pasanen A,Kivirikko K,Myllyharju J.Roles of the human hypoxia-inducible factor(HIF)-3a variants in the hypoxia response[J].Cell Mol Life Sci,2011,68:3885-901.
[8]Makino Y,Kanopka A,Wilson WJ,et al.Inhibitory PASdomain protein(IPAS)is a hypoxia-inducible splicing variant of the hypoxia-inducible factor-3alpha locus[J].J Biol Chem,2002,277(36):32405-8.
[9]Patel SA,Simon M C.Biology of hypoxia-inducible factor-2alpha in development and disease[J].Cell Death Differ,2008,15(4):628-34.
[10]Gu Y Z,Moran SM,Hogenesch JB,et al.Molecular characterization and chromosomal localization of a third alpha-class hypoxia inducible factor subunit,HIF3alpha[J].Gene Expr,1998,7(3):205-13.
[11]Hara S,Hamada J,Kobayashi C,et al.Expression and characterization of hypoxia-inducible factor(HIF)-3alpha in human kidney:suppression of HIF-mediated gene expression by HIF-3alpha[J].Biochem Biophys Res Commun,2001,287(4):808-13.
[12]Drevytska T,Gavenauskas B,Drozdovska S,et al.HIF-3alpha mRNA expression changes in different tissues and their role in adaptation to intermittent hypoxia and physical exercise[J].Pathophysiology,2012,19(3):205-14.
[13]Maynard M A,Evans A J,Hosomi T,et al.Human HIF-3alpha4 is a dominant-negative regulator of HIF-1 and is down-regulated in renal cell carcinoma[J].FASEB J,2005,19(11):1396-406.
[14]Heidbreder M,Qadri F,Johren O,et al.Non-hypoxic induction of HIF-3alpha by 2-deoxy-D-glucose and insulin[J].Biochem Biophys Res Commun,2007,352(2):437-43.
[15]Maynard M A,Evans A J,Shi W,et al.Dominant-negative HIF-3α4 suppresses VHL-Null renal cell carcinoma progression[J].Cell Cycle,2007,6(22):2810-6
[16]Yamashita T,Ohneda O,Nagano M,et al.Abnormal heart development and lung remodeling in mice lacking the hypoxiainducible factor-related basic helix-loop-helix PASprotein NEPAS[J].Mol Cell Biol,2008,28:1285-97.
[17]Huang Y,Ochieng J K,Kempen M B,et al.Hypoxia inducible factor 3alpha plays a critical role in alveolarization and distal epithelial cell differentiation during mouse lung development[J].PLoSOne,2013,8(2):e57695.
[18]Gontan C,Munck A,Vermeij M,et al.Sox2 is important for two crucial processes in lung development:branching morphogenesis and epithelial cell differentiation[J].Dev Biol,2008,317(1):296-309.
[19]Shu W,Lu M M,Zhang Y,et al.Foxp2 and Foxp1 cooperatively regulate lung and esophagus development[J].Development,2007,134(10):1991-2000.
[20]Hatanaka M,Shimba S,Sakaue M,et al.Hypoxia-inducible factor-3alpha functions as an accelerator of 3T3-L1 adipose differentiation[J].Biol Pharm Bull,2009,32(7):1166-72.
[21]Ando H,Natsume A,Iwami K,et al.A hypoxia-inducible factor(HIF)-3αsplicing variant,HIF-3α4 impairs angiogenesis in hypervascular malignant meningiomas with epigenetically silenced HIF-3α4[J].Biochem Biophys Res Commun,2013,433(1):139-44.
