IgD在自身免疫病中作用的研究进展
2014-05-19陈文生吴育晶
陈文生,黄 琼,吴育晶,魏 伟
(安徽医科大学临床药理学研究所,抗炎免疫药物教育部重点实验室,安徽合肥 230032)
近年来,随着对炎症免疫相关疾病病理机制认识的不断深入,IgD在炎症免疫相关疾病中的作用越来越受到重视。研究表明,多种自身免疫病的发生伴随着IgD水平的异常及抗IgD自身抗体的产生[1],故IgD可能与自身免疫病的发生密切相关,并可能成为治疗靶点。本文将结合IgD结构和功能的特点讨论其与自身免疫病的关联。
1 IgD的结构特点
1.1 人IgD和其他Ig亚型间的结构差异 人IgD由两条重链和两条轻链组成,同其他免疫球蛋白(immunoglobulin,Ig)一样分为可变区(V区)和恒定区(C区),IgD的 Cδ1段和Cδ2段与其他Ig的结构相似,但其Cδ3区由于部分脯氨酸的缺失以及 N-聚糖的增多而异于其他 Ig。有研究表明[2],正是这种结构上的改变导致了IgD功能上的特异。人IgD绞链区由64个氨基酸组成,比其他Ig的绞链区都要长,人IgD以“T”形存在,其他免疫球蛋白亚型以“Y”形存在,如Fig 1所示,IgD的两个Fab段可以围绕Fc段旋转,这样的特殊结构使IgD有能力结合较低浓度的抗原[3]。
1.2 不同种属间IgD的结构差异 B淋巴细胞通过RNA选择性剪切和免疫球蛋白类别转换(class switch recombination,CSR)来表达IgD。人类和其他灵长类动物的IgD重链有3个Cδ结构域,啮齿类动物有两个,而偶蹄类动物的IgD由1个Cμ和两个Cδ结构域嵌合而成,这种嵌合结构在鱼类IgD中常见,可能是为便于重链与轻链发生共价结合而产生[4]。
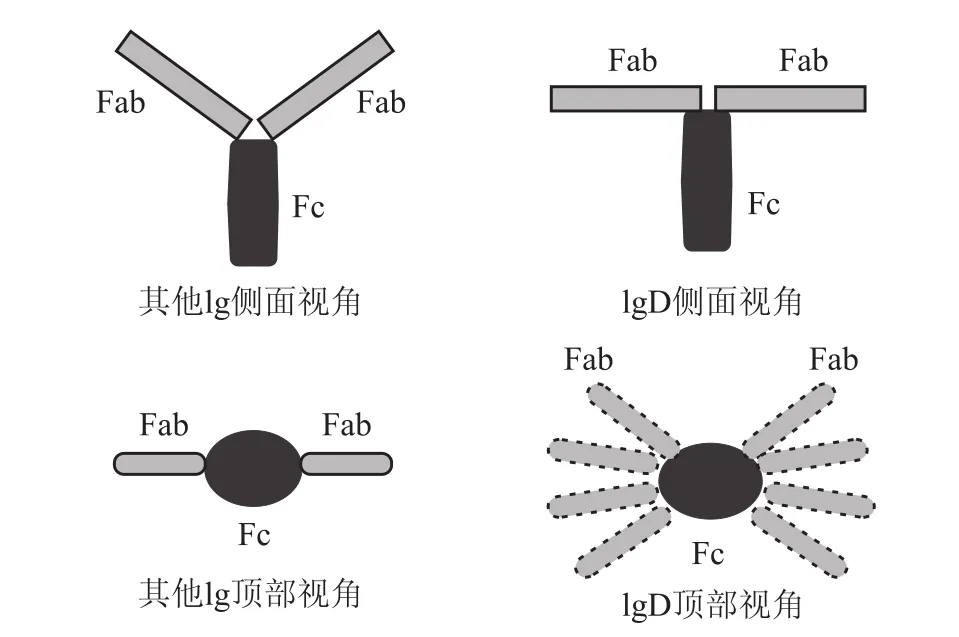
Fig 1 Sturctural differences between IgD and other Ig subtypes
2 IgD的功能特点
2.1 膜型 IgD(membrane IgD,mIgD)的功能特点 IgD是一种B细胞接受抗原刺激后,通过CSR产生的抗原受体,它分布于人和鼠的B细胞表面[5]。在B细胞成熟前,细胞仅表达IgM,当B细胞离开骨髓进入二级淋巴器官时,IgD才开始表达。IgD和IgM在B细胞上共同表达的作用和机制尚不明确[6]。有实验表明,抗原在体内与非成熟B细胞上的IgM结合会导致细胞凋亡,而抗原与IgD结合则不会导致凋亡的发生[7];另有实验得出了相反的结果,最小剂量的抗IgD抗体能导致成熟静息B细胞的凋亡[8]。产生这些矛盾实验结果的原因可能是:B细胞抗原受体(B cell receptor,BCR)信号的转导取决于实验所采用的B细胞系种类、B细胞的成熟程度以及B细胞表面的受体密度[9]。
一些基因敲除小鼠的实验显示,IgD可以替代IgM在B细胞生长早期的功能,且无论IgD或IgM重链缺陷都对小鼠影响轻微,可见IgD和IgM可以很大程度上的补偿彼此的功能[10]。
IgD、IgM的调节作用与不同的信号分子相关,有研究表明[11]:IgD可能通过提高IgM引起细胞活化、无能、死亡所需的信号强度来调节IgM的功能,因此,IgD作为受体传递的可能是一类耐受信号。然而,另有研究认为[12],IgD和IgM在B细胞上的共同表达可以使得不同浓度的抗原得到最优的结合,这一模型通过IgD晶体结构实验得到了验证。
2.2 SIgD(secreted IgD,sIgD)的功能特点 sIgD的免疫学功能尚不明确,目前的研究主要集中在:sIgD与IgD受体(IgD receptor,IgD-R)相关免疫功能及机制的探索。
上世纪80年代,有研究通过E-玫瑰花结实验证明人淋巴细胞上存在IgD-R[13],进一步研究显示IgD-R存在于小鼠CD4+和人CD4+、CD8+T细胞上,IL-2、IL-4、PHA以及低聚、偶联的IgD可以上调该类受体的表达[14]。然而,表达IgD-R的T细胞并不直接活化,T细胞特异性激动剂和低聚IgD共同作用于T细胞并不能增强IgD-R在T细胞上的表达。上述实验现象提示:IgD-R虽然在T细胞上表达,但却并不直接对T细胞产生效应。Tamma等[15]关于小鼠IgD-R+脾脏T细胞和IgD-R+T细胞杂交瘤的研究部分验证了这一推论,该研究发现这些IgD-R+细胞向其所处的生长介质中释放sIgD结合因子(secreted IgD binding factor,sIgD-BF),而sIgD-BF可能是IgD-R的效应形态。故IgD-R可能分为两种形态:一种是分布于相关淋巴细胞表面的IgD-R,另一种是分布于相关淋巴细胞临近介质中的sIgD-BF。目前,相关研究仅确定了IgD-R分布以及表达量,但IgD-R的编码基因和分子结构尚不明确,因此IgD与IgD-R特异性结合后产生的相关作用及机制需进一步探究。
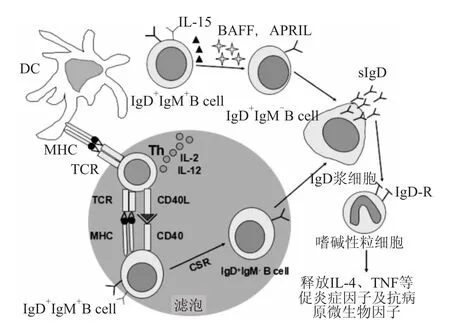
Fig 2 Model of human IgD regulation and function
许多固有免疫细胞可以结合sIgD,体内试验证明嗜碱性粒细胞表面结合了大量的sIgD,肥大细胞表面也可以结合sIgD,这一实验结果在体外试验中也得到了验证[16]。目前,关于上呼吸道黏膜sIgD的研究较为深入,其免疫调节过程如Fig 2所示:树突状细胞(dendritic cells,DCs)呈递抗原,激活CD4+Th细胞,之后Th细胞在CD40L、IL-2和IL-21的作用下诱导CSR过程,使得滤泡内IgM+IgD+B细胞转换为IgM-IgD+B细胞,这是一条T细胞依赖的通路。另一方面,B细胞活化因子(B-cell activating factor,BAFF)、增殖配体(a proliferation-inducing ligand,APRIL)、IL-2和 IL-15介导滤泡外IgM+IgD+B细胞通过CSR转换为IgM-IgD+B细胞,这是一条非T细胞依赖通路。通过这两条通路形成的IgMIgD+B细胞分化分泌sIgD的浆细胞。sIgD可以与嗜碱性粒细胞上的IgD-R发生结合,通过释放IL-4、IL-1β、TNF等促炎症因子及抗病原微生物因子来增强免疫功能[17]。
3 IgD与自身免疫病
3.1 IgD与T细胞相互作用参与自身免疫病 类风湿关节炎(rheumatoid arthritis,RA)是一种常见的自身免疫病,众多研究显示CD4+T细胞在RA发病中起着重要作用。CD4+T细胞会过量渗透进入炎性滑膜区域,可以提呈自身抗原,产生自身抗体;CD4+T细胞功能与RA易感基因相关,如HLA-DRB1等位基因[18]。CD4+T细胞可分化形成 Th1、Th2和Th17。最初,生成IFN-γ的Th1被认为是RA发病中起主要作用的T细胞,然而,越来越多的相关研究显示Th17才是起关键作用的细胞亚群[19]。
CD4+T细胞是IgD-R表达较多的细胞亚群,有研究显示,T细胞上IgD-R的偶联可以起到抑制凋亡的保护作用,IgD-R可以促进同源T细胞和幼稚B细胞之间形成免疫突触,从而增强抗原呈递及抗体生成等功能[20]。然而,病理状态下,IgD与IgD-R过度结合,则可能导致自身抗体生成增多、自身免疫反应增强,从而介导自身免疫病的发生。
3.2 IgD与B细胞相互作用参与自身免疫病 自身反应性B细胞的失调会导致自身免疫病,相关研究表明[21],自身反应性B细胞消除能力缺陷是RA和系统性红斑狼疮(systemic lupus erythematosus,SLE)的成因之一。转基因Ig模型也显示抗核染质B细胞和类风湿因子特异性B细胞(rheumatoid factor specific B cell,RFB)在发生自身免疫的组织中处于活化状态,这种B细胞的异常活化可能由Th细胞的功能改变或自身抗原的结合能力改变引起[22]。B细胞在RA和SLE等自身免疫病发生和发展过程中的作用可能有:分泌病原性抗体及细胞因子,活化相关T细胞。B细胞还可能通过非抗体依赖的途径来激活和加剧自身免疫反应:在SLE模型中,抗原特异性B细胞作为一种重要的抗原提呈细胞(antigen presenting cell,APC)为相应的 T细胞呈递抗原[23];胶原性关节炎(collagen induced arthritis,CIA)模型中,在其他种类APC正常存在的情况下,B细胞的缺失导致T细胞反应的明显下降;B细胞产生的一些细胞因子是自身免疫反应中重要的调节分子。
3.3 抗IgD抗体在自身免疫病治疗中的应用 有研究报道,在病程初期采用抗IgD抗体对小鼠CIA进行治疗,结果显示:抗IgD抗体可选择性地消耗成熟B细胞,而未对调节性B细胞(regulatory B cell,B-reg)产生影响,导致小鼠体内自身抗体含量明显降低,而对T细胞依赖抗原相关的抗体未产生影响。另外,抗IgD抗体在调节B-reg和T-reg水平的同时还可增强Th1/Th2细胞介导的适应性免疫反应。综上,抗IgD抗体治疗可以在不影响T、B细胞适应性免疫反应的情况下,通过选择性的消耗反应性B细胞来发挥抗炎和诱导免疫耐受作用[24]。
4 展望
T、B淋巴细胞以及固有免疫细胞功能异常是自身免疫病的重要病理机制,研究显示mIgD和sIgD都与上述免疫细胞的功能紧密相关,IgD在自身免疫病发生发展的可能作用机制假设如下:(1)B细胞上mIgD所转导的信号介导了B细胞的过度活化,导致自身反应性增强、自身抗体产生增多。(2)病理状态下,sIgD相对增多,sIgD与T细胞或其他免疫细胞上的IgD-R结合,介导了免疫细胞的异常活化或凋亡减少,导致机体异常免疫应答。
IgD作为一类免疫球蛋白,自发现、至今已近50年,然而其在免疫系统中的作用和机制仍是一个谜。近年来,一些研究提示了IgD在自身免疫病的发生发展中可能扮演重要角色。相信随着对IgD与IgD-R以及相关信号通路的不断研究,将会揭示其在自身免疫病中的作用和机制,为自身免疫病的合理治疗和新药研发提供新的思路。
参考文献:
[1]Koelsch K,Zheng N Y,Wilson P C,et al.Mature B cells class switched to IgD are autoreactive in healthy individuals[J].J Clin Invest,2007,117(6):1558-65.
[2]Stavnezer J,Amemiya CT.Evolution of isotype switching[J].Semin Immunol,2004,16(4):257-75.
[3]Edholm E S,Bengten E,Wilson M,et al.Identification of two IgD+B cell populations in hannel catfish,Ictalurus punctatus[J].J Immunol,2010,185(7):4082-94.
[4]Amin A R,Tamma SM,Coico R F,et al.The immunoaugmenting properties of murine IgD reside in its C delta 1 and C delta 3 regions:potential role for IgD-associated glycans[J].Int Immunol,1993,5(6):607-14.
[5]Abney E R,Parkhouse R M.Candidate for immunoglobulin D present on murine B lymphocytes[J].Nature,1974,252(5484):600-2.
[6]Kim K M,Reth M.The B cell antigen receptor of class IgD induces a stronger and more prolonged protein tyrosine phosphorylation than that of class IgM[J].J Exp Med,1995,181(3):1005-14.
[7]Ales-Martinez J E,Warner G L,Scott D W.Immunoglobulins D and M mediate signals that are qualitatively different in B cells with an immature phenotype[J].Proc Natl Acad Sci USA,1988,85(18):6919-23.
[8]Peckham D,Andersen-Nissen E,Ashman R F,et al.Difference in apoptosis induction between surface IgD and IgM[J].Int Immunol,2001,13(5):285-95.
[9]Geisberger R,Crameri R,Achatz G.Models of signal transduction through the B-cell antigen receptor[J].Immunology,2003,110(4):401-10.
[10]Roes J,Rajewsky K.Immunoglobulin D(IgD)-deficient mice reveal an auxiliary receptor function for IgD in antigen-mediated recruitment of B cells[J].J Exp Med,1993,177(1):45-55.
[11]Geisberger R,Lamers M,Achatz G.The riddle of the dual expression of IgM and IgD[J].Immunology,2006,118(4):429-37.
[12]Loset G A,Roux K H,Sandlie I,et al.Differential segmental flexibility and reach dictate the antigen binding mode of chimeric IgD and IgM:implications for the function of the B cell receptor[J].J Immunol,2004,172(5):2925-34.
[13]Sjöberg O.Presence of receptors for IgD on human T and non-T lymphocytes[J].Scand J Immunol,1980,11(4):377-82.
[14]Tamma S M,Coico R F.IgD-receptor-positive human T lymphocytes.Ⅱ.Identification and partial characterization of human IgD-binding factor[J].J Immunol,1992,148(7):2050-7.
[15]Tamma SM,Coico R F.IgD-receptor(IgD-R)cross-linking partially protects murine T cells from dexamethasone-induced apoptosis[J].J Leukoc Biol,2003,73(6):764-70.
[16]Chen K,Xu W,Cerutti A,et al.Immunoglobulin D enhances immune surveillance by activating antimicrobial,proinflammatory and B cell-stimulating programs in basophils[J].Nat Immunol,2009,10(8):889-98.
[17]Chen K,Cerutti A.The function and regulation of immunoglobulin D[J].Curr Opin Immunol,2011,23(3):345-52.
[18]Komatsu N,Takayanagi H.Autoimmune arthritis:the interface between the immune system and joints[J].Adv Immunol,2012,115:45-71.
[19]Irmler I M,Gajda M,Bräuer R.Exacerbation of antigen-induced arthritis in IFN-gamma-deficient mice as a result of unrestricted IL-17 response[J].J Immunol,2007,179(9):6228-36.
[20]Chen K,Cerutti A.New insights into the enigma of immunoglobulin D[J].Immunol Rev,2010,237(1):160-79.
[21]Yurasov S,Tiller T,Nussenzweig M C,et al.Persistent expression of autoantibodies in SLE patients in remission[J].J Exp Med,2006,203(10):2255-61.
[22]Fields M L,Nish SA,Erikson J,et al.The influence of effector T cells and Fas ligand on lupus-associated B cells[J].J Immunol,2005,175(1):104-11.
[23]O′Neill SK,Shlomchik M J,Finnegan A,et al.Antigen-specific B cells are required as APCs and autoantibody-producing cells for induction of severe autoimmune arthritis[J].J Immunol,2005,174(6):3781-8.
[24]Nguyen T G,Little CB,Morris JM,et al.Anti-IgD antibody attenuates collagen-induced arthritis by selectively depleting mature B-cells and promoting immune tolerance[J].J Autoimmun,2010,35(1):86-97.
