H9亚型禽流感病毒钦州株HA基因的克隆与遗传进化
2014-05-18陈进喜张华智郑彦梓翟成兵施开创
陈进喜,张华智,黄 梅,郑彦梓,翟成兵,熊 毅,郑 敏,施开创
(1.钦州市动物疫病预防控制中心,广西 钦州 535099;2.钦州市畜牧站,广西 钦州 535099;3.广西动物疫病预防控制中心,广西 南宁 530001)
禽流感(avian inf l uenza,AI)是由正黏病毒科的A型流感病毒(inf l uenza A viruses)引起的一种禽类的感染和/或疾病综合症,对家禽造成严重的危害。自1966年首次分离到H9N2亚型禽流感病毒以来,该型病毒目前已经普遍存在于世界各国的禽群中,流行毒株可分为北美及欧亚两种系[1]。1994年陈伯伦等[2]报道了中国大陆的首例禽流感病毒感染并从广东鸡群中分离到了H9N2亚型禽流感毒株。2003年分离到的人H9N2禽流感病毒(A/Hong Kong/2108/03)是一个新的重组体,很可能来源于当地的活禽市场[3]。AIV存在着广泛的抗原漂移和抗原转变现象,其抗原变异频率很高,而在AIV所编码的各种蛋白质中,又以HA基因的突变频率最高;HA还是AIV中最主要的保护性抗原。本研究采集活禽交易市场临床健康鸡的气管和泄殖腔的棉拭子,对其进行H9亚型禽流感检测,并扩增HA基因序列,力图阐述钦州地区H9N2禽流感HA基因的分子遗传规律。为研究禽流感病毒的分布以及控制禽流感的流行、蔓延提供数据支持。
1 材料与方法
1.1 病毒
研究样品采自钦州市活禽交易市场临床健康鸡的气管和泄殖腔棉拭子,由广西区动物疫病预防控制中心实验室保存; 阳性对照:禽流感(H5+H9)二价灭活疫苗,青岛易邦生物工程有限公司产品。
1.2 SPF鸡胚
9-11日龄无特定病原(SPF)鸡胚由山东省农科院家禽研究所禽病研究中心提供。
1.3 引物
参考GenBank上已发表并上传的H9N2亚型基因序列,选择保守区域应用Oligo6.0软件设计并合成特异性引物:H9亚型分型用的引物:H9F:5’-CTYCACACAGARCACAATGG-3’;H9R:5’-GCCACACTTGHGGTRTC-3’;扩增HA基因片段全长用引物:HA1:5’-GGGGAAT TTCACAACCACTCAAG-3’;HA2:5’-TTGAGT AGAAACAAGGGTGTTTC-3’;反转录用引物Uni12:5’-AGCRAAAGCA-GG-3’。
1.4 病毒RNA的提取及RT-PCR
按试剂盒操作说明提取病毒总RNA。以总RNA为模板,采用反转录引物Uni12进行反转录,获得cDNA。以cDNA为模板,应用设计的特异性引物,进行PCR扩增。反应体系总体积25 μL: 内含模板2.0 μL,加10×Reaction Buffer 2.5 μL,dNTP(2.5 mmol/μL)2.0 μL,Taq DNA酶(5 U/μL)0.25 μL,MgCl2(25 mmol/L)1 μL,上下游引物(25 pmol/μL)各0.5 μL,加ddH2O至25 μL,置PCR仪中。反应程序:94 ℃ 5 min;94 ℃ 45 s,(H9亚型分型用57 ℃、扩增HA基因片段全长用54 ℃)45 s,72 ℃ 1 min,35 个循环;72 ℃ 10 min。反应结束后,取PCR产物7 μL在1.5%琼脂糖凝胶上电泳,置凝胶成像系统中观察结果。
1.5 PCR产物的克隆及序列分析
回收PCR产物中目的片段,回收目的DNA按连接试剂盒说明书进行连接,然后将连接产物转化到感受态细胞DH5α中,涂平板,培养16 h,挑斑接种于LB液体培养基中,置37 ℃摇床培养16 h;取1.0 mL培养物抽提重组质粒,进行酶切及PCR鉴定,将阳性菌株送宝生物工程(大连)公司进行测序。将测序结果与已发表的基因序列进行分析。
2 结果
2.1 H9亚型禽流感病毒PCR检测结果
用H9亚型禽流感病毒分型引物对临床健康的鸡棉拭子进行检测,出现488 bp大小的片段确定为H9亚型禽流感病毒阳性(图1)。所采集的样品中有1份为H9亚型禽流感病毒阳性(命名为:A/Chicken/Guangxi/qz40/2009,简称qz40)。

图1 H9亚型禽流感病毒PCR检测结果图
2.2 病毒的增殖和纯化
将H9亚型禽流感病毒PCR检测阳性的棉拭子经双抗处理后8000 r/min离心10 min,取上清液接种9 ~ 11日龄SPF鸡胚绒毛尿囊膜(0.2 mL/胚),37 ℃孵育,间隔6 ~ 12 h照胚一次,死胚置4 ℃过夜,无菌收集24 ~ 96 h死亡的鸡胚尿囊液,用1%的鸡红细胞测定其血凝效价。然后根据血凝效价进行10-1000倍有限稀释后继续接种SPF鸡胚,至少传3代,收获血凝滴度≥7log2的尿囊液于 -70℃保存备用。
2.3 HA基因的PCR扩增
应用HA1/HA2引物扩增出qz40株的HA基因全长,PCR产物经10 g/L琼脂糖凝胶上电泳分析表明,与预期结果相符(图2)。回收PCR产物中目的片段,连接、转化、挑斑,将酶切及PCR鉴定阳性的菌株进行测序。
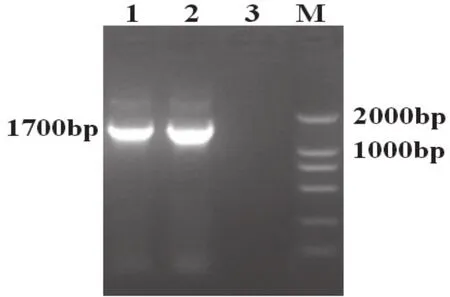
图2 HA基因的PCR产物电泳图
2.4 序列测定及同源性分析
将所获qz40株的HA基因核苷酸序列的测序结果拼接后(图3),HA基因的cDNA包括一个完整的开放阅读框,编码区由1683 bp组成,共编码560个氨基酸,包括信号肽(1aa – 18aa)、HA1(19aa – 337aa)、HA2(339aa – 560aa) 和一个精氨酸(338aa)。经DNAStar软件将所获毒株的HA基因序列与从GenBank中获得的26个H9N2亚型禽流感病毒HA基因核苷酸序列及其推导的氨基酸序列进行同源性比较,由表1可知,qz40株与A/chicken/Guangxi/21/2006株的核苷酸同源性最高,达99.9%,与A/Duck/HongKong/Y439/97株的核苷酸同源性最低,仅为82.8%;推导的氨基酸同源性介于87.5% ~ 99.6%之间。
2.5 功能位点分析
推导的氨基酸序列分析表明,qz40株含有7个潜在的N-糖基化位点(Asn-X-Ser/Thr,X为除Pro以外的任意氨基酸),其中5个位于HA1部分,为NST(29aa-31aa)、NVS(141aa-143aa)、NRT(218aa-220aa)、NTT(298aa-300aa)、NVS(305aa-307aa);2个 位 于 HA2部 分, 为NGT(492aa-494aa)、NGS(55laa-553aa)。 裂解位点序列(335aa-341aa)分析发现,该毒株为RSSR↓GLF,为非高致病力毒株的典型特征,对禽类为低致病力。构成受体结合位点的氨基酸位于 109aa、161aa、163aa、191aa、198aa、202aa、203aa(qz40株的受体结合位点的氨基酸依次YWTNTLY),通过与其他参考毒株相比,198位氨基酸变化较大,其中本研究的qz40株为T,而A/Chicken/Beijing/1/94为 V,A/Quail/HongKong/G1/97和A/Duck/HongKong/Y439/97都为E,其它位点的氨基酸相对保守。

图3 qz40株HA基因测序结果
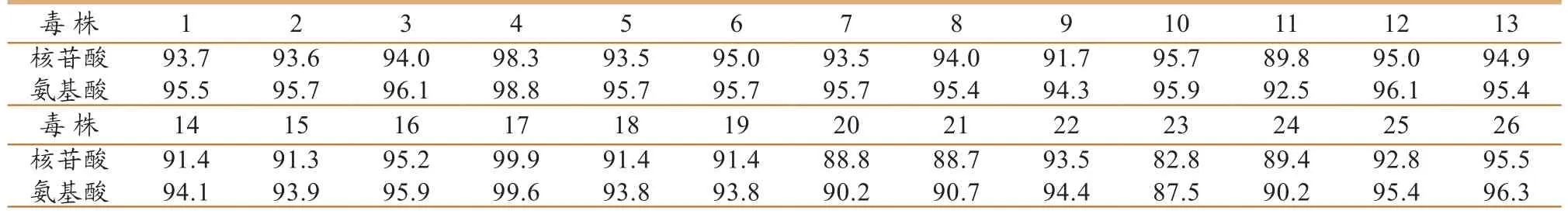
表1 qz40株HA基因的核苷酸序列及其推导的氨基酸序列与其他H9N2毒株的比对(%)
2.6 进化树分析
基于H9亚型禽流感病毒HA基因核苷酸序列绘制的系统发育进化树(图4)。从进化树可以分为3个群系,分别为:A/Chicken/Beijing/1/94(Ck/Bei-like)、A/Quail/HongKong/G1/97(G1-like)和A/Duck/HongKong/Y439/97(Y439-like)。从本研究建立的基于H9亚型HA基因进化树发现,钦州分离的qz40株属于A/Chicken/Beijing/1/94(Ck/Bei-like)群系,与国内参考株的亲缘关系较近。而与A/Duck/HongKong/Y439/97(Y439-like)群系的亲缘关系较远。

图2 HA基因的PCR产物电泳图
3 讨论
禽流感病毒粒子表面有二种主要的纤突,分别为血凝素(HA)和神经氨酸酶(NA)。HA在病毒吸附及穿膜过程中起关键作用,能刺激机体产生中和抗体从而中和病毒的感染[4]。目前,在世界范围内的各种家禽和野禽中,按照HA的表面抗原来区分,有15种特异的HA(H1-H15)[5]。
本研究运用H9亚型分型引物对采集的棉拭子进行PCR筛选,选取PCR检测阳性的样品,接种SPF鸡胚进行病毒的增殖,通过有限稀释克隆法纯化3代后,获得相对稳定、血凝滴度较高(≥71og2)的种毒,分装后保存于 -70℃备用。本研究从采集自活禽交易市场的临床健康的鸡棉拭子中获得1份阳性样品,将其命名为A/Chicken/Guangxi/qz40/2009(简称qz40),在运用特异引物扩增包含HA全基因在内的序列,并对其进行序列的测定,测序结果拼接后显示HA基因的cDNA包括一个完整的开放阅读框,编码区由1683 bp组成,共编码560个氨基酸。经DNAStar软件将所获毒株的HA基因序列与从GenBank中获得的26个H9N2亚型禽流感病毒HA基因核苷酸序列及其推导的氨基酸序列进行同源性比较,结果显示该毒株与参考毒株的核苷酸序列同源性为82.8%~99.9%,与A/chicken/Guangxi/21/2006株的核苷酸同源性最高;推导氨基酸同源性介于87.5%~99.6%之间。
HA上潜在的糖基化位点对病毒的受体亲和力、病毒毒力等多种生物学特性有影响[6]。本研究中qz40株含有7个潜在的N-糖基化位点,其中5个位于HA1部分、2个位于HA2部分,与其他参考株没有太大的差异。本研究所得的qz40株的HA的裂解位点为RSSR↓GLF,没有多个连续的碱性氨基酸插入,具有典型的低致病性禽流感病毒的基因特征[7]。流感病毒HA受体结合位点的特异性决定其感染的宿主范围。当病毒适应新宿主时,决定受体结合特异性的234位氨基酸通常首先发生变化[8]。本研究的病毒在该位置为亮氨酸(L),而香港人源H9N2毒株该处氨基酸也为亮氨酸(L)。
研究表明H9N2禽流感主要由2个大的分支,欧亚种系和北美种系。欧亚种系又分为3个群系,分 别 为:A/Chicken/Beijing/1/94(Ck/Bei-like)、A/Quail/HongKong/G1/97(G1-like) 和 A/Duck/HongKong/Y439/97(Y439-like)[1]。本研究中的钦州分离毒株qz40株属于欧亚种系的A/Chicken/Beijing/1/94(Ck/Bei-like)群系,与国内参考株的亲缘关系较近,特别是与分离自广西的毒株A/chicken/Guangxi/21/2006株处于同一小分支上,这与核苷酸同源性比对结果相符。而与A/Duck/HongKong/Y439/97(Y439-like)群系的亲缘关系较远。
H9N2亚型禽流感病毒虽在亚洲的许多国家流行,各毒株的亲缘关系也较近,但有很多因素能影响其HA基因的变异,从而影响其致病力。已有研究表明来自于禽类的H9N2禽流感病毒能偶尔从禽类传染给哺乳动物,包括人和猪[9]。这种跨种间传播表明目前的H9N2禽流感病毒对人仍有潜在的感染性。因人类与家禽的接触机会历来很多,人类与禽类流感之间的联系自然非常密切,加之1997年香港禽流感病毒对人类的直接感染[10],目前对禽流感的公共卫生意义的认识日益受到重视。在禽流感所编码的各种蛋白质中,又以HA基因的突变频率最高, HA还是禽流感中最主要的保护性抗原,因此本研究就HA基因序列的测定,为H9N2亚型禽流感广西分离株遗传变异分析和分子流行病学调查提供了重要材料,也为制订H9亚型禽流感的综合防制措施提供了依据,因此建议临床上在重视H5亚型禽流感感染控制的同时,还应该加强对H9亚型禽流感感染的防控。
[1]Xu K M, Smith G J D, Bahl J, et al.The genesis and evolution of H9N2 inf l uenza viruses in poultry from southern China, 2000 to 2005[J].J Virol, 2007, 81(19): 10389-10401.
[2]陈伯伦,张泽纪,陈伟斌.禽流感研究I:鸡A型禽流感病毒的分离与血清学初步鉴定[J].中国兽医杂志,1994,22(10):3-5.
[3]Butt K M, Smith G J D, Chen H, et al.Human infection with an avian H9N2 inf l uenza A virus in Hong Kong in 2003[J].J Clin Microbiol, 2005, 43(11): 5760-5767.
[4]甘孟侯.禽流感[M].北京:北京农业大学出版,1995:27-38.
[5]殷震.刘景华.动物病毒学[M]2版.北京:科技出版社,1997.
[6]Li C, Yu K, Tian G, et al.Evolution of H9N2 influenza viruses from domestic poultry in Mainland China[J].Virol,2005, 340(1): 70-83.
[7]Liu H Q, Liu X F, Cheng J, et al.Phylogenetic Analysis of the Hemagglutinin Genes of Twenty-Six Avian Influenza Viruses of Subtype H9N2 Isolated from Chickens in China During 1996–2001[J].Avian Dis, 2003, 47(1): 116-127.
[8]Matrosovich M, Tuzikov A, Bovin N, et al.Early alteration of the receptor-binding properties of H1, H2 and H3 avian influenza virus hemagglutinins after their introduction into mammals[J].J Virol, 2000, 74(18): 8502-8512.
[9]Lin Y P, Shaw M, Gregory V, et al.Avian-to-human transmission of H9N2 subtype inf l uenza A viruses: Relationship between H9N2 and H5N1 human isolates[J].Proc Natl Acad Sci USA,2000 , 97 (17):9654-9658.
