松花江流域典型鱼类的游泳能力比较实验研究
2014-05-15王从锋刘德富寇方露莫伟均石小涛
熊 锋 王从锋,2 刘德富 寇方露 莫伟均 刘 伟 石小涛
(1.三峡大学 水利与环境学院,湖北 宜昌 443002;2.三峡地区地质灾害与生态环境湖北省协同创新中心,湖北 宜昌 443002;3.湖北工业大学 资源与环境学院,武汉 430068;4.三峡大学 化学与生命科学学院,湖北 宜昌 443002)
目前世界各国修建了大量的大坝、堤堰等水利工程,在防洪、发电、灌溉等方面凸显巨大作用,但其对生态环境也产生了极大的影响.特别是修建拦河大坝后,对鱼类生长、繁殖产生了严重的干扰,其中以对洄游、半洄游性鱼类的影响最大[1].随着人们对这种影响的逐步认识,过鱼设施的研究与建设成为减小这种影响的一种有效途径.而确定保护对象,并对其游泳能力和生态习性进行研究,成为研究和建设过鱼设施的首要工作.因此对不同鱼类游泳能力及其生态行为学的研究,不仅具有理论上的意义,还可为过鱼、拦鱼和诱鱼等设施的设计提供依据,保证其水电工程储能设施发挥较大的功能[2].鱼类游泳能力是指鱼类游泳的持续时间和强度[3,4],鱼类的游泳类型分为持续式游泳、耐久式游泳和爆发式游泳,对应衡量鱼类游泳能力的指标主要有临界游速、持久游速和突进游速[5].
临界游速亦称最大可持续游泳速度[6],主要用于评价鱼类有氧运动能力,目前应用最为广泛[7],主要原因是测定临界游速时间相对比较短,所需的实验鱼数目相对更少[4].临界游速也可用于估测不同环境因子对鱼类游泳能力的影响,用于评价栖息地环境条件改变可能导致的生态结果[8].突进游速是鱼类所能达到的最大速度,维持时间很短,通常小于20s[9],是鱼类逃脱捕获及逃避猎食者时采取的重要方式[10],对鱼类的生存有重要影响.
本研究利用自制的鱼类游泳能力研究实验装置,采用递增流速法分别测定了不同体长规格的3种鱼类的临界游速及突进游速,并系统分析了3种鱼类体长与临界游速及突进游速之间的关系,为研究鱼类游泳能力和鱼道设计提供基础数据与参考资料.
1 材料与方法
1.1 试验器具
本研究所用的试验装置为实验室自制的鱼类游泳能力测定装置(如图1所示).试验装置的工作原理是:假设在密闭空间内,鱼类的游速与水流速度相等,其中水流速度通过调频器改变电机的转速来实现,并利用多孔整流器使观察区截面各处水流速度近似均匀,通过计算水流速度获得鱼类的游泳速度[11].试验装置的外箱体积为54L(86cm×37cm×17cm),其中,观察区体积为4.5L(37cm×11cm×11cm).

图1 鱼类游泳能力测定装置
装置的水流速度由Vectrino小威龙点式流速仪(Nortek)测定.按4Hz的步长调节电动机频率,测量相应频率下的平均流速.建立流速(Y)与调节频率(X)之间的线性关系:Y=0.0329 X-0.0176,R2=0.9974.(如图2所示)
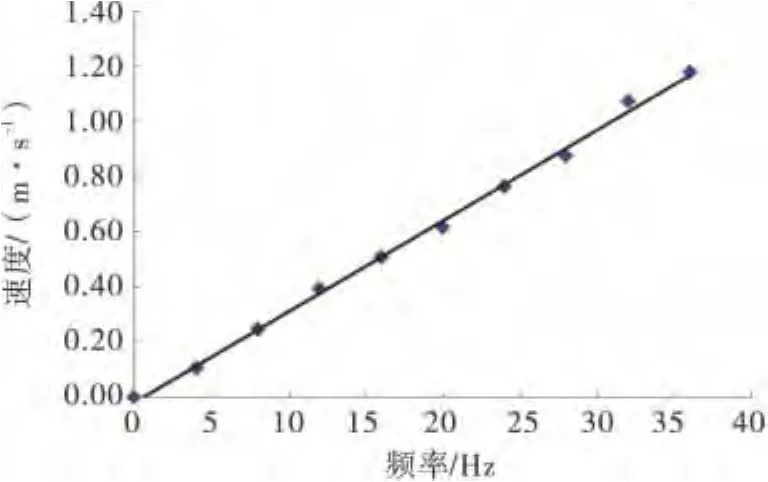
图2 速度与频率的关系
1.2 实验材料
实验鱼捕捞于松花江松花湖.试验前在循环控温水池中(水容量2.5m3)暂养5d.暂养期间每天投喂1次饵料,暂养用水为曝气自来水,光照为室内自然光,水温控制在(21±2)℃,溶氧含量不小于7mg/L,氨氮水平不大于0.01mg/L.
1.3 测定方法
1.3.1 临界游速
正式实验前先进行临界游速预试验,估测每种实验鱼的绝对临界游速(单位为cm·s-1),以此确定其Ucrit(单位为cm·s-1)的速度增量.选取1尾鱼置于水槽鱼类观察区内,在适应流速的基础上,以每2min以0.4BL·s-1的体长(BL,单位为cm)速度增加水流速度,直至实验鱼疲劳[11].此时的临界游速为估计值,作为正式试验的参考值.
实验前将单尾实验鱼放入观察区中,为消除转移过程对鱼体的胁迫,将鱼放置在低流速下适应1h.实验开始后,将流速增至60%Ucrit估计值[12],持续15 min后速度增加15%Ucrit估计值,以此方式不断提升流速,直至实验鱼达到运动力竭状态(即停留在观察区拦网处20s以上)不能继续游泳为止.记录实验鱼的力竭时刻并将其从观察区中取出,并测定其体重及体长.Ucrit计算公式[13]为
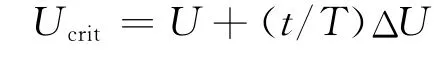
式中,U为顺利完成设定时间的最大游速(实验鱼力竭前一档流速),T为设定各速度梯度下的时间间隔(15min),t为鱼类疲劳前一档流速梯度的游泳时间(t<15min),ΔU 为速度增量(15%Ucrit估计值).实测即为,为消除体长对Ucrit的影响,更有效地比较3种鱼类的相关能力,本研究采用相对临界游速(Urcrit,单位为BL·s-1),其计算公式为
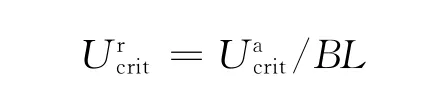
1.3.2 突进游速
本试验中,突进游速的测试也采用“递增流速法”,与临界游速测试方法基本一致,只是将流速提升时间间隔改为20s.突进游速计算公式与临界游速计算公式一致,本研究采用相对突进游速,单位为BL·s-1)计算公式为/BL,式中为绝对突进游速(cm·s-1).
1.4 数据处理
试验数据用Excel 2010进行常规计算后,用SPSS 17.0软件进行多重比较,分析3种鱼类临界游速和突进游速及其之间的差异.统计值用平均数±标准差(Mean±SD)描述,差异显著水平为P<0.05.通过回归分析对体长与临界游速、体长与突进流速的关系进行拟合.
2 结果与分析
2.1 临界游速
根据鱼类游速公式,经分析和计算得出长春鳊、怀头鲇、翘嘴鲌的临界游速(见表1).
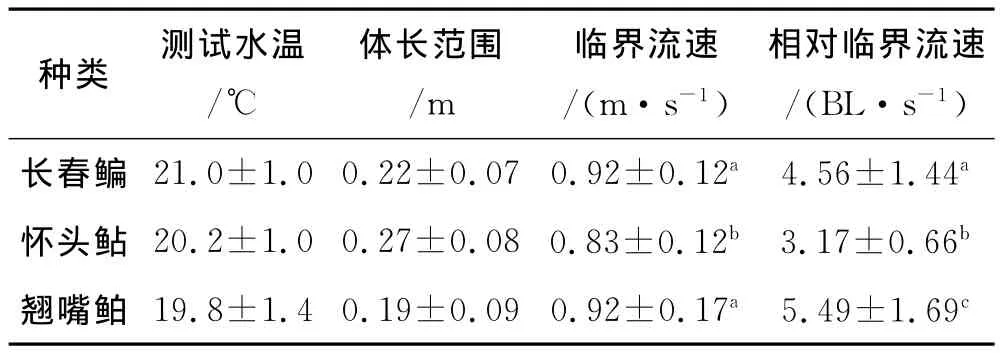
表1 3种鱼类的临界游速
经统计检验得出3种实验鱼中,怀头鲇与其他鱼类的相对临界游速存在显著性差异(P<0.05),其中翘嘴鲌的相对临界游速最高,长春鳊次之,怀头鲇最低.
从图3可见,3种鱼类的相对临界游速均随体长的增长呈递减趋势,其中翘嘴鲌的相对临界游速与体长的关系最为显著,其相对临界游速与体长的相关关系拟合方程为Y=-0.1789 X+8.8982(R2=0.8913).
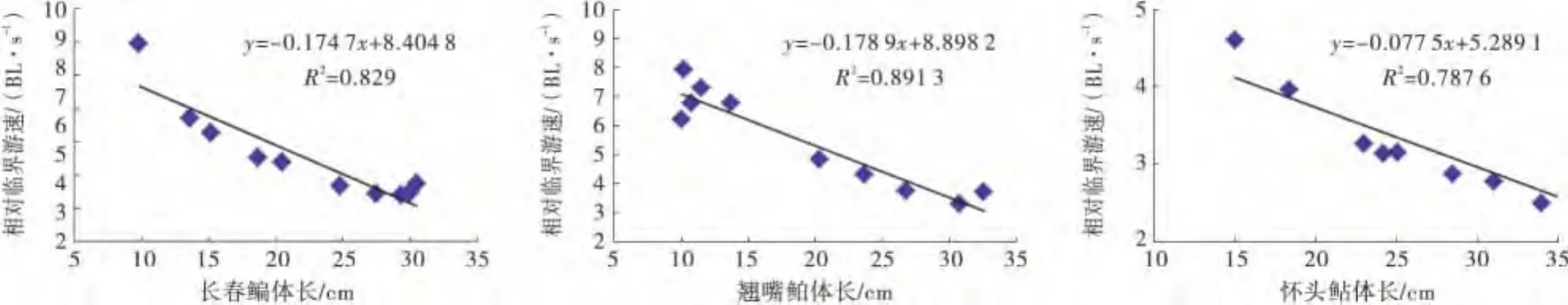
图3 体长与相对临界游速关系
2.2 突进游速
根据鱼类游速公式,经分析和计算得出长春鳊、怀头鲇、翘嘴鲌的突进游速(见表2).
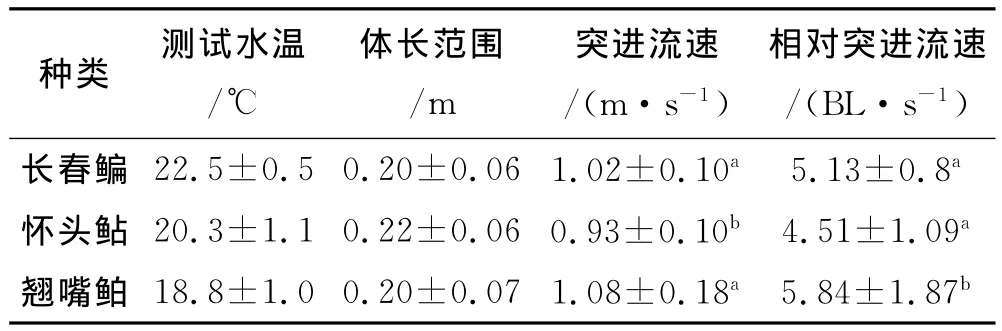
表2 3种鱼类的突进游速
经统计检验得出4种实验鱼中,翘嘴鲌与其他两种鱼的相对突进游速存在显著性差异(P<0.05),其中翘嘴鲌的相对突进游速最高,长春鳊次之,怀头鲇最低.
从图4可见,3种鱼类的相对突进游速均随体长的增长呈递减趋势,其中怀头鲇的相对突进游速与体长的关系最为显著,其相对临界游速与体长的相关关系拟合方程为Y=-0.1639 X+8.1267(R2=0.8810).
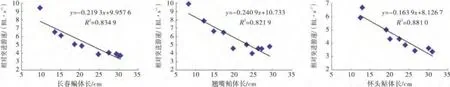
图4 相对突进游速与体长的关系
3 讨 论
本研究得出怀头鲇相对临界游速为3.17±0.66 BL·s-1(Mean±SD),这与鲜雪梅[12]对南方鲇的相对临界游速(3.14±0.11BL·s-1)研究结论相似.翘嘴鲌相对临界游速为5.84±1.87BL·s-1(Mean±SD),这与文献[14]报道的生活习性相似的鲢的相对临界游速(5.87±1.87BL·s-1)一致.文献[15]报道鳙的相对临界游速为4.57±0.56BL·s-1,与本研究得出长春鳊的相对临界游速为4.56±1.44BL·s-1(Mean±SD)结论相似.本研究中3种实验鱼的临界游速随鱼体长的增加而上升,相对临界游速随鱼体长的增加而下降,本结论与 Wardle[16]及官之梅等[17]分别对海水鱼和7种淡水鱼类的测试结果一致.
根据临界游速和体长的回归曲线,分析比较相同体长条件下不同鱼类的游速.本研究中的翘嘴鲌与类似规格的其他鱼类相比表现出不同的游泳能力,如15℃条件下体长为26.29cm鲫(Carassius auratus)的相对临界游速为3.36BL·s-1[18],20℃条件下体长为32.45cm的美国红鱼的相对临界游速为2.47 BL·s-1,均低于这两种鱼的相对临界游速.本研究中的翘嘴鲌出现异于其他鱼类的游泳能力,主要原因可能是由于游泳能力与鱼类的生活习性、体型及其所栖息的环境相关.翘嘴鲌为凶猛肉食性鱼类,大多在湖泊、水库和河流等大水体中,成鱼生活在敞水区的中上层,游泳迅速,善跳跃,性情暴躁,其较高的临界游速与其生活习性有着密切联系.
突进游泳是鱼类在环境应激反应下的游泳行为,也是穿越鱼道高流速区域的重要保障,对鱼类的生存有着重要的影响[19].由突进游速和体长的回归曲线可知,3种实验鱼的突进游速随体长的增加而增加,相对突进游速随体长的增加而减少,本研究的结论与Ming-Fong[20]的测试结果一致.根据测得的突进游速,以20s为突进时间可估测出最大突进距离,本研究中3种鱼类的突进游泳距离在20m左右,此长度则为这3种实验鱼在鱼道内最大流速区域内能顺利通过的距离,从而以此长度作为设计鱼道休息室的间隔距离.同时,改变温度基本上不会对鱼类的突进游泳能力产生显著的影响[21],因此,突进游速能更好的为过鱼设施设计提供参考数据.
影响鱼类游泳能力的因素,除了自身因素外还包括溶氧量、光照、温度等[22].当在一定程度上降低水域中氧的浓度或升高二氧化碳的浓度时,鱼类的游泳能力会随着降低[23-26].本研究中由于试验用水的溶氧量存在着±1mg·L-1的波动,因此对实验鱼的游泳能力测试存在着影响.对于实验鱼而言,水温越高,新陈代谢越强,肌肉收缩时间截短,故游速也越高[27].本研究中由于试验持续的时间较长,受实验室气温影响,循环水槽中的水温在18~23℃波动,这可能是导致实验鱼游速差异性显著的外界因素之一.
对于野生个体和养殖个体之间游泳能力是否存在显著差异,Schreer和Dunmall对野生和养殖的大西洋鲑鱼(Salmo Salar)的游泳能力进行了研究,实验结论表明两者间不存在明显差异[28].本研究的3种鱼类的野生与养殖个体间游泳能力是否存在差异,需要进行实验验证.另外,本研究所选取的鱼类捕捞于松花江流域,该地区气候条件异于其他流域,这3种鱼类的游泳能力是否异于其他流域的同种鱼类,仍需要进一步实验论证.
[1]涂志英,袁 喜,王从锋,等.亚成体巨须裂腹鱼游泳能力及活动代谢研究[J].水生生物学报,2012(4):682-688.
[2]涂志英,袁 喜,韩京成,等.鱼类游泳能力研究进展[J].长江流域资源与环境,2011(S1):59-65.
[3]He P,Wardle C S.Endurance at Intermediate Swimming Speeds of Atlantic Mackerel,Scomber Scombrus L.,Herring,Clupea Harengus L.,and Saithe,Pollachius Virens L.[J].Journal of fish biology,1988,33(2):255-266.
[4]Hammer C.Fatigue and Exercise Tests with Fish[J].Comparative Biochemistry and Physiology Part A:Physiology,1995,112(1):1-20.
[5]Plaut I.Critical Swimming Speed:Its Ecological Relevance[J].Comparative Biochemistry and Physiology-Part A:Molecular &Integrative Physiology,2001,131(1):41-50.
[6]Fisher R,Wilson S K.Maximum Sustainable Swimming Speeds of Late-Stage Larvae of Nine Species of Reef Fishes[J].Journal of Experimental Marine Biology and E-cology,2004,312(1):171-186.
[7]Day N,Butler P J.The Effects of Acclimation to Reversed Seasonal Temperatures on the Swimming Performance of Adult Brown Trout Salmo Trutta[J].Journal of Experimental Biology,2005,208(14):2683-2692.
[8]Zeng L,Cao Z,Fu S,et al.Effect of Temperature on Swimming Performance in Juvenile Southern Catfish[J].Comparative Biochemistry and Physiology-Part A:Molecular & Integrative Physiology,2009,153(2):125-130.
[9]郑金秀,韩德举,胡望斌,等.与鱼道设计相关的鱼类游泳行为研究[J].水生态学杂志,2010(5):104-110.
[10]Yanase K,Eayrs S,Arimoto T.Influence of Water Temperature and Fish Length on the Maximum Swimming Speed of Sand Flathead,Platycephalus Bassensis:Implications for Trawl Selectivity[J].Fisheries research,2007,84(2):180-188.
[11]石小涛,陈求稳,刘德富,等.胭脂鱼幼鱼的临界游泳速度[J].水生生物学报,2012(1):133-136.
[12]鲜雪梅,曹振东,付世建.4种幼鱼临界游泳速度和运动耐受时间的比较[J].重庆师范大学学报:自然科学版,2010(4):16-20.
[13]Brett J R.The Respiratory Metabolism and Swimming Performance of Young Sockeye Salmon[J].Journal of the Fisheries Board of Canada,1964,21(5):1183-1226.
[14]房 敏,蔡 露,高 勇,等.温度对鲢幼鱼游泳能力及耗氧率的影响[J].水生态学杂志,2013(3):49-53.
[15]蔡 露,涂志英,袁 喜,等.鳙幼鱼游泳能力和游泳行为的研究与评价[J].长江流域资源与环境,2012(S2):89-95.
[16]Wardle C S.Limit of Fish Swimming Speed[J].1975,255:725-727.
[17]官之梅,刘文郁,陈佩薰.鱼类学论文集[M].北京:科学出版社,1981:133-139.
[18]赵文文,曹振东,肖月吉,等.低温条件下鲫鱼幼鱼的游泳能力及其能量消耗[J].重庆师范大学学报:自然科学版,2011(1):13-17.
[19]叶 超,王 珂,黄福江,等.异齿裂腹鱼游泳能力初探[J].淡水渔业,2013(3):33-37.
[20]Yeh M,Ho C,Lee M.Burst Swimming Speeds of Two Minnows Acrossocheilus Paradoxus and Varicorhinus Barbatulus in Relation to Water Temperature and Body Size[J].台湾水产学会会刊,2010,37(1):37-48.
[21]Nikora V I,Aberle J,Biggs B,et al.Effects of Fish Size,Time-to-Fatigue and Turbulence on Swimming Performance:a Case Study of Galaxias Maculatus[J].Journal of Fish Biology,2003,63(6):1365-1382.
[22]Dickson K A,Donley J M,Sepulveda C,et al.Effects of Temperature on Sustained Swimming Performance and Swimming Kinematics of the Chub Mackerel Scomber Japonicus[J].Journal of Experimental Biology,2002,205(7):969-980.
[23]Sepulveda C,Dickson K A.Maximum Sustainable Speeds and Cost of Swimming in Juvenile Kawakawa Tuna(Euthynnus Affinis)and Chub Mackerel(Scomber Japonicus)[J].Journal of Experimental Biology,2000,203(20):3089-3101.
[24]Schurmann H,Steffensen J.Spontaneous Swimming Activity of Atlantic cod Gadus Morhua Exposed to Graded Hypoxia at Three Temperatures[J].Journal of Experimental Biology,1994,197(1):129-142.
[25]Domenici P,Steffensen J F,Batty R S.The Effect of Progressive Hypoxia on Swimming Activity and Schooling in Atlantic Herring[J].Journal of Fish Biology,2000,57(6):1526-1538.
[26]Beaumont M W,Butler P J,Taylor E W.Exposure of Brown Trout Salmo Trutta to a Sublethal Concentration of Copper in Soft Acidic water:Effects Upon Gas Exchange and Ammonia Accumulation[J].Journal of experimental biology,2003,206(1):153-162.
[27]乔云贵,黄洪亮,黄妙芬,等.不同淡水鱼类游泳速度的初步研究[J].湖南农业科学,2012(15):116-119.
[28]Dunmall K M,Schreer J F.A Comparison of the Swimming and Cardiac Performance of Farmed and Wild Atlantic Salmon,(Salmo salar),Before and after Gamete Stripping[J].Aquaculture,2003,220(1):869-882.
