普生轮藻浸提液对两种淡水藻类的化感抑制作用及其数学模型
2014-05-14何宗祥张庭廷
何宗祥,刘 璐,李 诚,张庭廷,*
(1.安徽师范大学数学计算机科学学院,芜湖 241000;2.安徽师范大学生命科学学院,芜湖 241000)
湖泊富营养化及其导致的蓝藻水华暴发是目前较为突出的环境问题之一。蓝藻水华肆虐,不仅对生态环境造成严重危害,其释放的蓝藻毒素可被水生生物吸收和累积,并随食物链传递,对人类的健康造成极大威胁[1-2],因此,对蓝藻水华的预防和治理是当前迫切需要解决的重要课题。传统的除藻方法有的存在着二次环境污染,如化学法除藻;有的操作时间长、难度大、费用高,如物理法除藻[3-4]。作为淡水水体中主要生产者的高等水生植物,对维持水体生态平衡有着其无与伦比的优势。一方面高等水生植物可以吸收和降低水体中氮、磷等营养盐;同时,不少高等水生植物还能够分泌和释放抑制藻类生长的化感物质,这些物质多为植物次生代谢物,在自然水体中易于降解,不会在生态系统中长期积累[5],因此,利用高等水生植物化感作用及其化感物质来进行抑藻受到了广泛的关注[4-5]。
目前,已有数十种高等水生植物被报道具有化感抑藻作用,如穗花狐尾藻(Myriophyllum spicatum)、金鱼藻(Ceratophyllum demersum)、凤眼莲(Eichharnia crassipes)、芦苇(Phragmites communis)等被认为是具有较强化感抑藻活性的植物[5-7]。已有大量文献报道轮藻与其它大型水生植物相比,更能保持水体的清澈透明[8-9]。Van Donk 等[10]通过调查提出轮藻植物对浅水区域水体透明度的保持,起着至关重要的作用;Steemann-Nielsen[11]认为在微咸的水域中,由于普生轮藻植物的存在,浮游植物只有极低的生物量。但迄今为止有关轮藻化感作用研究报道还相对较少。普生轮藻是广布性种类,几乎遍布我国绝大部分省区[12],本课题研究人员在芜湖地区调查发现:凡普生轮藻生长的区域,水体清澈,未见水华蓝藻生长。为了深入研究普生轮藻的化感抑藻效应及其规律,包括普生轮藻对单一毒性铜绿微囊藻以及混合藻类的抑制作用、其抑藻的时-效与量-效关系等,本文就普生轮藻浸提液的化感抑藻作用及其数学模型的建立进行了较深入研究。由于经典的描述单个种群增长的 Logistic模型只能表征种群在一定空间范围内的种群增长趋势,不能定量计算种群在逆境胁迫条件下任何一个时间节点的生物量;Lotka-Volterra模型描述了两种群在竞争状态下的数量关系,但在有抑制剂等其他因素干扰等限制条件下群落类的排斥与共存问题尚未彻底解决,而使得任何一个时间节点生物量的获得难以实现。本项目组曾对单一化感物质对单一藻类或两种化感物质对单一藻类的抑制作用模型进行了探讨[13-14],但两种藻类在竞争状态下的化感模型研究尚付阙如。因此本课题在传统的阻滞增长模型即逻辑斯特模型(Logistic模型)和种间竞争模型即Lotka-Volterra模型基础上,通过微元法建立了普生轮藻浸提液对单纯产毒铜绿微囊藻、单纯斜生栅藻以及两藻混合时的抑制数学模型。该研究将可为利用数学和生物学的有机结合来解决复杂多变的生物学问题以及为我国的蓝藻水华治理等提供重要的理论与实验依据。
1 实验材料、方法
1.1 实验材料
普生轮藻(Chara vulgaris)采集于安徽师范大学荷花塘;试验藻种产毒铜绿微囊藻(Microcystis aeruginosa)和斜生栅藻(Scenedesmus obliquus)均购自中国科学院水生生物研究所,产毒铜绿微囊藻采用BG-11培养基培养;斜生栅藻采用 HB-4培养基培养。
1.2 实验方法
1.2.1 藻种的培养
于无菌条件下,在已洗净消毒的500 mL三角锥形瓶中各加入100 mL培养基,接入藻种,摇匀,置光照培养箱中进行培养。培养条件:光照度4000 Lx,光周期12 L∶12 D,温度(24±2)℃,pH 值7.0。每天摇动锥形瓶4次,并加入适量培养基,使藻细胞处于呈几何级数增长的对数增长期[15]。
1.2.2 普生轮藻浸提液制备
取普生轮藻整株,洗净泥沙,用蒸馏水淋洗,经滤纸吸干其表面水分后,称其鲜重50 g,剪成2 cm长的小段,放入大烧杯中用500 mL蒸馏水将其完全浸泡,在4℃下放置96 h后,先用定性滤纸过滤出浸提液,然后减压浓缩至5 mL,再将其用0.22 μm微孔滤膜过滤,除去微生物[16-17]。
1.2.3 普生轮藻浸提液抑藻实验
(1)对单纯毒性铜绿微囊藻或斜生栅藻的抑制实验
在已灭菌的三角烧瓶中,分别接入50 mL处于对数增长期的铜绿微囊藻或斜生栅藻藻液,藻液起始浓度为2.4—2.5×105个/mL,然后加入普生轮藻浸提液使其起始浓度分别为 0.00,0.20,0.40,0.60,0.80,1.00 ml/L[相当于 0.00,2.00,4.00,6.00,8.00,10.00 g植物(鲜重)/L],每组3个平行。放在光照培养箱中按藻种培养条件进行培养,每隔24 h用血球计数板在OLYMPUS双筒显微镜下计数藻细胞[18]。
(2)对毒性铜绿微囊藻和斜生栅藻共培养条件下的抑制实验
在已灭菌的三角烧瓶中,分别接入25mL处于对数增长期的铜绿微囊藻和斜生栅藻藻液,两藻液起始浓度均为 1.2—1.3×105个 /mL(混合藻密度为2.4—2.5×105cells/mL),然后加入普生轮藻浸提液使其起始浓度分别为 0.00,0.20,0.40,0.60,0.80,1.00 ml/L,每组3个平行。放在与单一藻抑藻实验相同条件下进行培养,每隔24 h进行藻细胞计数[19]。
2 实验结果与模型建立
2.1 普生轮藻浸提液的抑藻效应
普生轮藻浸提液的抑藻实验结果,见图1—图4。

图1 不同浓度普生轮藻浸提物对产毒铜绿微囊藻生长的影响Fig.1 Abundance of toxic Microcystis aeruginosa after 120 h incubations with Chara vulgaris extracts concentrations of 0.00,0.20,0.40,0.60,0.80 and 1.00 ml/L.Error bars represent the standard deviation for triplicate treatments

图2 斜生栅藻在不同浓度普生轮藻浸提物作用下藻密度的实验值Fig.2 Abundance ofScenedesmus obliquus after 120 h incubations with Chara vulgaris extracts concentrations of 0.00,0.20,0.40,0.60,0.80 and 1.00 ml/L.Error bars represent the standard deviation for triplicate treatments
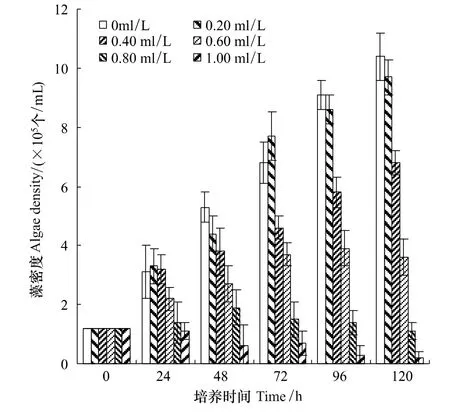
图3 共培养时不同浓度普生轮藻浸提物对产毒铜绿微囊藻生长的影响Fig.3 The growth inhibitory effect of different concentrations of Chara vulgaris extracts on toxic M.aeruginosa while the two algal species co-incubations.Error bars represent the standard deviation for triplicate treatments
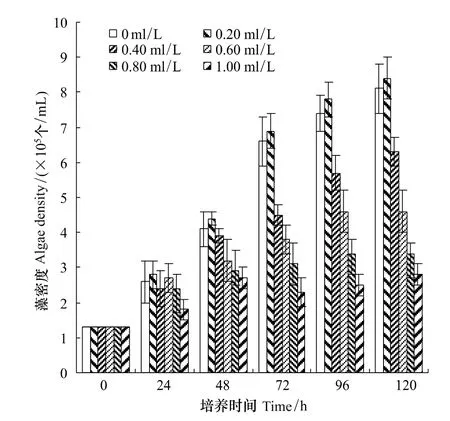
图4 共培养时不同浓度普生轮藻浸提物对斜生栅藻生长的影响Fig.4 The growth inhibitory effect of different concentrations of Chara vulgaris extracts on S.obliquus while the two algal species co-incubations;Errorbars representthestandard deviation for triplicate treatments
由图1—图4可知,普生轮藻浸提液对单独培养的毒性铜绿微囊藻、斜生栅藻以及共生的毒性铜绿微囊藻和斜生栅藻均有抑制作用。但是无论是单独还是两藻共生,普生轮藻浸提液对毒性铜绿微囊藻的抑制作用均明显强于对斜生栅藻。该结论来自以下三方面的分析:(1)将单独培养的铜绿微囊藻和斜生栅藻对照组藻密度进行比较,尽管两藻的起始密度相同,但第24小时开始两藻密度就有明显差异,铜绿微囊藻密度明显较斜生栅藻高,经配对t-检验,P<0.05,这种差异一直持续到第120小时;同样,该差异在两藻共培养的对照组中更为明显,经配对t-检验,P<0.05或0.01(第96以及第120小时)。(2)实验组中,无论是单独培养还是两藻共培养,从第96小时开始,同期铜绿微囊藻密度明显低于同期斜生栅藻,经配对 t-检验,P<0.01。总结(1)与(2),不难看出这种完全不同的变化趋势,反映出普生轮藻对铜绿微囊藻的强烈抑制作用。(3)从抑制率更能直观地说明以上结论,以最大抑制率第120小时为例,对铜绿微囊藻单独和共培养时分别为96.77%和98.08%,而对斜生栅藻则为67.71% 和65.43%。普生轮藻的这一特性对治理以铜绿微囊藻为优势种群的蓝藻水华具有重要的应用意义。
2.2 数学模型的确立
2.2.1 在不同初始浓度普生轮藻浸提液作用下,单一藻(铜绿微囊藻、斜生栅藻)藻密度随时间变化的数学模型
根据铜绿微囊藻在其种群生长过程中,其总量具有有限增长[18]的特点,以及在普生轮藻浸提液胁迫下铜绿微囊藻生长的特征,在传统的Logistic模型基础上,通过微元分析法,增加了普生轮藻浸提液对铜绿微囊藻的化感抑制作用项[13-14](可理解为由于普生轮藻浸提液的化感抑制作用,从而使得铜绿微囊藻细胞死亡率增高或增殖率减低,相当于振动系统中的阻尼项)。根据产毒铜绿微囊藻在不同浓度普生轮藻浸提液环境中藻密度在不同时间的实验值(图1)进行拟合(由于可采用的拟合方法有很多,在这里所采用的是惠普公司生产的H38G图形计算器)可得:在不同浓度普生轮藻浸提液环境中,产毒铜绿微囊藻藻密度随时间变化的模型为:
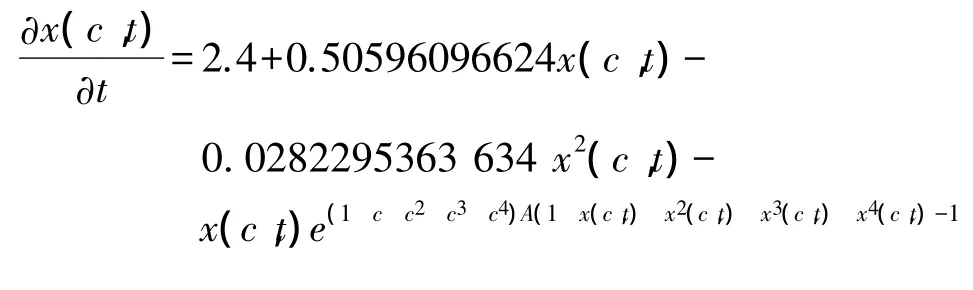

式中,c是普生轮藻浸提液的原始浓度 (ml/L);x(c,t)是产毒铜绿微囊藻在原始浓度为c(ml/L)的普生轮藻浸提液环境中,t时刻的藻密度值。
通过模型(1),可以计算出在图1相应的时间节点上产毒铜绿微囊藻藻密度的近似值(共30个数值)。通过计算可得图1所标的、通过实验得到的在不同原始浓度c(ml/L)普生轮藻浸提液环境中,产毒铜绿微囊藻的观察数据与利用上述模型(1)计算出的相对应的藻密度近似值之间的可决系数

根据模型(1)还可以得到在不同原始浓度c(ml/L)的普生轮藻浸提液环境中,产毒铜绿微囊藻随时间变化的趋势图(图5),从图5中可以比较方便、快捷地得到不同时间节点上普生轮藻浸提液的半抑制浓度(EC50)、最小有效浓度(MIC)等指标的预测值。例如,第120小时普生轮藻浸提液对产毒铜绿微囊藻的半抑制浓度(EC50)的预测值为0.483 ml/L,最小有效浓度(MIC)的预测值为0.738 ml/L、第72小时普生轮藻浸提液的半抑制浓度(EC50)的预测值为0.658 ml/L,最小有效浓度(MIC)的预测值为0.714 ml/L。
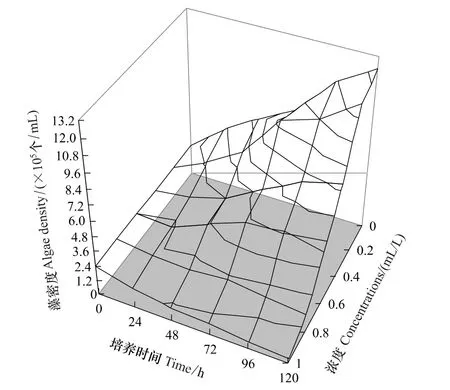
图5 在不同浓度的普生轮藻浸提液环境下产毒铜绿微囊藻藻密度的模型预测值Fig.5 The model predicted value of toxic M.aeruginosa density under the pressure of different initial concentrations of Chara vulgaris extracts
同理,根据斜生栅藻在不同浓度普生轮藻浸提液环境中藻密度在不同时间的实验值(图2)进行拟合(这里仍采用惠普公司生产的H38G图形计算器)可得:在不同浓度的普生轮藻浸提液环境中,斜生栅藻藻密度随时间变化的模型为:

式中,c是普生轮藻浸提液的原始浓度 (ml/L);y(c,t)是斜生栅藻在原始浓度为c(ml/L)的普生轮藻浸提液环境中,t时刻的藻密度值。
通过模型(2),可以计算出在图2相应的时间节点上斜生栅藻藻密度的近似值(共30个数值)。通过计算可得图2所标的、通过实验得到的在不同原始浓度c(ml/L)普生轮藻浸提液环境中,斜生栅藻的观察数据与利用上述模型(2)计算出的相对应的藻密度的近似值之间的可决系数R2为0.993。该可决系数反映了所拟合的在不同原始浓度c(ml/L)的普生轮藻浸提液环境中,斜生栅藻变化的数学模型(2),具有比较好的拟合度,是合理的、有效的、可靠和有意义的,基本能反映斜生栅藻变化的的实际情况。

图6 在不同浓度普生轮藻浸提液环境下斜生栅藻藻密度的模型预测值Fig.6 The model predicted value of toxic S.obliquus density under the pressure of different initial concentrations of Chara vulgaris extracts
根据模型(2)还可以得到的在不同原始浓度c(ml/L)的普生轮藻浸提液环境中,斜生栅藻随时间变化的趋势图(图6)。从图6可比较方便、快捷得到不同时间节点上普生轮藻浸提液对斜生栅藻的EC50、MIC等指标的预测值。例如,第120小时普生轮藻浸提液的EC50的预测值为0.655 ml/L、第72小时普生轮藻浸提液的EC50的预测值为0.779 ml/L。
2.2.2 在不同初始浓度普生轮藻浸提液环境中,铜绿微囊藻、斜生栅藻共生情况下,铜绿微囊藻、斜生栅藻藻密度随时间变化的数学模型
根据在不同初始浓度的普生轮藻浸提液环境中,铜绿微囊藻、斜生栅藻共生情况下,两个种群生活在同一空间,彼此竞争同一营养来源的特点,以及普生轮藻浸提液对共生藻的化感抑制作用,在传统的Lotka-Volterra模型的基础上,通过微元分析法,增加了普生轮藻浸提液对铜绿微囊藻、斜生栅藻的化感抑制作用项(可理解为由于普生轮藻浸提液的化感抑制作用,从而使得铜绿微囊藻细胞、斜生栅藻细胞死亡率增高或增殖率减低,相当于振动系统中的阻尼项)。根据在不同初始浓度的普生轮藻浸提液环境中,铜绿微囊藻、斜生栅藻共生情况下,铜绿微囊藻、斜生栅藻藻密度在不同时间的实验值(图3,图4)进行拟合(这里仍采用惠普公司生产的H38G图形计算器)得到:在不同初始浓度的普生轮藻浸提液环境中,在铜绿微囊藻、斜生栅藻共生情况下藻密度随时间变化的模型为:

式中,c是普生轮藻浸提液的原始浓度(mg/L);x(c,t)是产毒铜绿微囊藻在原始浓度为c(ml/L)的普生轮藻浸提液环境下,t时刻的藻密度值;y(c,t)是斜生栅藻在原始浓度为c(ml/L)的普生轮藻浸提液环境下,t时刻的藻密度值。


通过模型(3),可以计算出在图3、图4相应的时间节点上产毒铜绿微囊藻、斜生栅藻藻密度的近似值(共60个数值)。通过计算可得图3所标的、通过实验得到的在不同初始浓度的普生轮藻浸提液环境中,铜绿微囊藻、斜生栅藻共生情况下,产毒铜绿微囊藻的观察数据与利用上述模型(3)计算出的相对应的藻密度的近似值(共30个数值)之间的可决系数R2约为0.998。同理,通过计算可得图4所标的、通过实验得到的在不同初始浓度的普生轮藻浸提液环境中,铜绿微囊藻、斜生栅藻共生情况下,斜生栅藻的观察数据与利用上述模型(3)计算出的相对应的藻密度的近似值(共30个数值)之间的可决系数R2约为0.995。该两个可决系数反映了所拟合的在不同初始浓度普生轮藻浸提液环境中,铜绿微囊藻、斜生栅藻共生情况下,铜绿微囊藻、斜生栅藻藻密度随时间变化的模型(3)是合理的、有效的、可靠和有意义的,基本能反映在共生情况下铜绿微囊藻、斜生栅藻藻密度随时间变化的实际情况。
通过模型(3),还可以计算得出在不同初始浓度的普生轮藻浸提液胁迫下,铜绿微囊藻、斜生栅藻共生时,随着时间t→∞,铜绿微囊藻、斜生栅藻在小生境中相对稳定存在时的预测值(表1)。这类预测值在实际抑藻过程中,为抑藻方案的制定和实施提供了有价值的数据支撑,具有重要的实际意义。
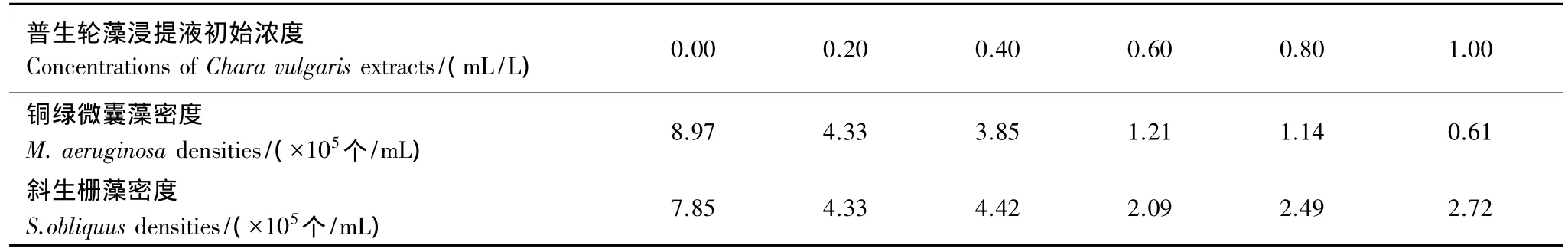
表1 不同初始浓度浸提液胁迫下共生的铜绿微囊藻与斜生栅藻在小生境中相对稳定时的预测值Table 1 The predicted abundance of S.obliquus and toxic M.aeruginosa while coexist under different concentrations of Chara vulgaris extracts
3 结论与讨论
3.1 普生轮藻浸提液对单独和共生状态的毒性铜绿微囊藻和斜生栅藻均具有抑制作用
高等水生植物分泌的化感物质抑藻时常具有种群特异性,这已有很多报道[19-20]。本课题组在研究脂肪酸类化感物质抑藻效应时发现亚油酸要比棕榈酸的抑藻效果好,而亚油酸对铜绿微囊藻(Microcystis aeruginosa)的抑藻效果又比对斜生栅藻(Scenedesmus obliquus)好[19];对多酚类化感物质的抑藻作用研究发现:对羟基苯甲酸对水华鱼腥藻(Anabaena flos-aquae)的抑制作用明显强于对蛋白核小 球 藻 (Chlorella pyrenoidosa)[21]; 从 芦 苇(Phragmites communis)中分离鉴定出化感物质2-甲基乙酰乙酸乙酯,对铜绿微囊藻和蛋白核小球藻具有很强的化感抑制作用,而对普通小球藻(Chlorella vulgaris)的抑制作用不明显[5];穗状狐尾藻对铜绿微囊藻、被甲栅藻(Scenedesmus armatus)等具有化感抑制作用,而对水华鱼腥藻几乎没有作用[22]。
本研究表明普生轮藻浸提液无论对单独的毒性铜绿微囊藻或斜生栅藻还是共生状态的毒性铜绿微囊藻和斜生栅藻均有很强抑制作用,且对毒性铜绿微囊藻的抑制作用要显著高于对斜生栅藻。由于水华发生时通常都是以微囊藻为优势种同时也存在一些诸如斜生栅藻等绿藻的多种藻类的混合体系,因此,该研究对如何合理利用高等水生植物化感作用来进行富营养化水体生态修复、水华藻类的防治等无疑具有重要的理论与应用意义。
3.2 所建立的化感物质抑藻模型具有普遍性意义
关于化感物质抑藻的数学模型研究目前相对较少。本研究在传统的Logistic模型基础上,通过微元分析法,增加了化感物质对单纯藻类的化感抑制作用项[12-13]建立了普生轮藻浸提液对单纯铜绿微囊藻或单纯斜生栅藻抑制作用的数学模型;同时在传统的Lotka-Volterra模型与Logistic模型的基础上,通过微元分析法,增加了普生轮藻浸提液对铜绿微囊藻和斜生栅藻共生时的化感抑制作用项,建立了化感物质对复合藻类抑制作用的数学模型。通过对同一时间节点上实验数据与所建立模型计算值的比较可得:增加了胁迫项的数学模型比传统的Logistic模型以及Lotka-Volterra模型能更好地表征单纯或复合藻类在普生轮藻浸提液胁迫下的生长,此与参考文献[13-14]中所讨论的结果具有一致性。另外,这几种模型均可有效表征和预测在一定范围内,产毒铜绿微囊藻、斜生栅藻及其混合体系在普生轮藻浸提液胁迫下藻密度随时间变化的规律;通过这些模型可方便地计算出在任何不同时间节点上普生轮藻浸提液抑藻的EC50、MIC等指标的预测值,该模型对指导和利用普生轮藻浸提液以及其他化感物质抑藻、为开发出有效的生物抑制剂以及设计和制定出合理、满足要求且具有经济参考的除藻方案等有重要参考价值。
[1]Jiang J L,Song R,Ren J H,Wang X R,Yang L Y.Advances in pollution of cyanobacterial blooms-producing Microcystins and their ecotoxicological effects on aquatic organisms.Progress in Chemistry,2011,23(1):246-253.
[2]Xie P.A review on the studies related to the effects of microcystins on human health.Journal of Lake Sciences,2009,21(5):603-613.
[3]Jeong J H,Jin H J,Sohn C H,Suh K H,Hong Y K.Algicidal activity of the seaweed Corallina pilulifera against red tide microalgae.Journal of Applied Phycology,2000,12(1):37-43.
[4]Jin Q,Dong S L.Comparative studies on the allelopathic effects of two different strains of Ulva pertusa on Heterosigma akashiwo and Alexandrium tamarense.Journal of Experimental Marine Biology and Ecology,2003,293(1):41-55.
[5]Li F M,Hu H Y.Allelopathic effects of disfferent macrophytes on the growth of Microcystis aeruginosa.Allelopathy Journal,2005,15(1):145-l52.
[6]Zhu J,Liu B Y,Wang J,Gao Y N,Wu Z B.Study on the mechanism ofallelopathic influence on cyanobacteria and chlorophytes by submerged macrophyte(Myriophyllum spicatum)and its secretion.Aquatic Toxicology,2010,98(2):196-203.
[7]Hilt S,Ghobrial M G N,Gross E M.In situ allelopathic potential of Myriophyllum verticillatum(Haloragaceae)against selected phytoplankton species.Journal of Phycology,2006,42(6):1189-1198.
[8]Peeter N,Tuvikene L,Feldmann T,Tõnno L,Künnap H,Luup H,Salujõe J,Nõges T.The role of charophytes in increasing water transparency:a case study of two shallow lakes in Estonia.Hydrobiologia,2003,506-509(1/3):567-573.
[9]Mulderij G,van D E,Roelofs J G M.Differential sensitivity of green algae to allelopathic substances from Chara.Hydrobiologia,2003,491(1/3):261-271.
[10]Van Donk E,Van Dund W J.Impact of submerged macrophytes including charophytes on phyto-and zooplankton communities:allelopathy versus other mechanisms.Aquatic Botany,2002,72(3/4):261-274.
[11]Steemann N E.Prevention of Hydroblogi-Polyteknisk Forlag.Lyngby,1984,36:25-37.
[12]Qiu L C,Ling Y J.Studies on Distribution of Characeae in China.Acta Hydrobiologica Sinica,2007,31(5):756-759.
[13]He Z X,Zhang T T.Mathematical model design of time-effect relationship analysis about the inhibition of four eighteen-cabon fatty acids on toxic Microcystis aeruginosa.Acta Ecologica Sinica,2011,31(23):7235-7243.
[14]He Z X,Zhang T T.Model development and optimization on the combined algal growth inhibition effects of two allelochemicals.Acta Scientiae Circumstantiae,2012,32(8):1894-1901.
[15]Zhang T T,Wang L L,He Z X,Zhang D.Growth inhibition and biochemicalchangesofcyanobacteria induced by emergent macrophyte Thalia dealbata roots.Biochemical Systematics and Ecology,2011,39(2):88-94.
[16]Nakai S,Inoue Y,Hosomi M.Allelopathic effects of polyphenols released by Myriophyllum spicatum on the growth of cyanobacterium Microcystis aeruginosa.Allelopathy J,2001,8:201-210.
[17]Nakai S,Yamada S,Hosomi M.Anti-cyanobacterial fatty acids released from Myriophyllum spicatum.Hydrobiologia,2005,543(1):71-78.
[18]Wang G.Practical Computer and Mathematical Modeling.Hefei:Anhui University Press,2000.
[19]Zhang T T,Zheng C Y,He M,Wu A P,Nie L W.Inhibition on algae of fatty acids and the structure-effect relationship.China Environmental Science,2009,29(3):274-279.
[20]Hilt S,Gross E M.Can allelopathically active submerged macrophytes stabilize clear-water states in shallow lakes?Basic and Applied Ecology,2008,9(4):422-432.
[21]Zhang T T,He M,Wu A P,Liu W N.Allelopathic inhibition of p-hydroxybenzoic acid on Microcystis aeruginosa Kueitz with no toxicological effects on Cyprinus carpio Linnaeus.Acta Scientiae Circumstantiae,2008,28(9):1887-1893.
[22]Körner S,Nicklisch A.Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes.Journal of Phycology,2002,38(5):962-971.
参考文献:
[1]姜锦林,宋睿,任静华,王晓蓉,杨柳燕.蓝藻水华衍生的微囊藻毒素污染及其对水生生物的生态毒理学研究.化学进展,2011,23(1):246-253.
[2]谢平.微囊藻毒素对人类健康影响相关研究的回顾.湖泊科学,2009,21(5):603-613.
[12]邱丽氚,凌元洁.中国轮藻植物分布研究.水生生物学报,2007,31(5):756-759.
[13]何宗祥,张庭廷.四种十八碳脂肪酸抑藻时-效关系分析的数学模型设计.生态学报,2011,31(23):7235-7243.
[14]何宗祥,张庭廷.两种化感物质联合抑藻的模型建立及数学最优化.环境科学学报,2012,32(8):1894-1901.
[18]王庚.实用计算机数学建模.合肥:安徽大学出版社,2000.
[19]张庭廷,郑春艳,何梅,吴安平,聂刘旺.脂肪酸类物质的抑藻效应及其构效关系.中国环境科学,2009,29(3):274-279.
[21]张庭廷,何梅,吴安平,聂刘旺.对羟基苯甲酸对铜绿微囊藻的化感效应以及对鲤鱼的毒性作用.环境科学学报,2008,28(9):1887-1893.
