草海流域水源功能区植物持水功能群划分
2014-05-05张建利喻理飞袁丛军严令斌
吴 华,张建利,喻理飞,袁丛军,严令斌
(1.贵州大学 喀斯特生态环境研究中心,贵阳55002;2.开阳县生态文明建设局,贵阳550300;3.贵州科学院 山地资源研究所,贵阳550001)
植物功能群是研究复杂生态系统结构和功能的有效途径,植物功能群以植物的功能行为和属性为特征,是沟通植物结构和功能与生态系统属性的桥梁,对植物功能群的研究已经成为理解复杂生态系统功能必要的一步[1]。功能群是被定义为对特定环境有相似反应的一类物种[2]。植物功能性状是植物与环境关系的演绎者,故其在生态学领域应用颇多[3-5]。植物与环境的相互关系一直是生态学研究的核心问题。早在l9世纪,生态学家及植物学家们就开始关注植物性状,并发现部分植物性状对环境变化有很好的响应和适应表现[6],目前功能群研究最核心的问题仍在于决定植物功能群划分的植物特征选择上[7-9]。
国内外生态学者对植物功能群做了大量开拓性的工作,研究者们尝试从不同的角度,用不同的方法,在不同的地点对不同的生态系统对植物进行功能群的划分,有关植物功能群的理论、方法逐渐形成,为植物功能群在生态系统动态和功能中的作用等进一步研究奠定了基础,但目前有关植物功能群研究尚存在的科学问题如下:① 研究层次不一,从全球尺度、群落尺度到分子尺度都有,对植物功能群形成的机制及其对生态系统的贡献等方面的研究需进一步深入;② 研究地点分布零星,应该有一个先主后次的研究计划;③研究方法各种各样,功能指标的选取不一,计算方法各有差异,无法对研究成果进行整合;④ 这些阶段性的研究是分散的,缺少一个整体的计划;⑤ 区域功能群研究没有能全面调查,一些研究只针对某一地点进行,这样势必会造成研究成果的不完整,对后继工作是一种浪费[10]。
综上所述,前人为植物功能群的划分多基于植物生理、生态适应性、生活史或对某一生态过程相关以及物种行为相联系的生物学特性与植物功能关系进行功能群划分,其研究对象多集中在草原生态系统[11-14],但通过揭示植物形态特征与植物服务功能关系,并划分植物功能群的研究少见报道。本文拟通过草海流域植物水源涵养功能区中植物形态特征与持水功能关系的揭示,划分植物持水功能群,既丰富植物功能群研究的内容,又可为该区域植物群落水源涵养功能的充分发挥、优化群落结构提供依据。
1 研究区概况
草海湖位于贵州省西部威宁县城的西南侧,处云贵高原中部乌蒙山脉之腹地,是贵州最大的淡水天然湿地湖泊,也是一个典型的高原喀斯特湿地生态系统。地理 位 置 为 北 纬 26°49′—26°05′,东 经 104°12′—104°18′[9-10]。区内属山地暖温带湿润季风气候,年平均气温10.6℃,7月份平均气温最高为l7.7℃,1月份最低为1.9℃,冬季长,无夏季。年平均降雨量950.9 mm,干湿季节明显,5—10月份为雨季,其降雨量占全年的88%,12月份至次年3月份为旱季,降雨量仅占全年的5%[15-16]。草海区域森林组成以中、幼龄为主,成熟林极少,灌丛稀疏。保护区森林覆盖率低,林下少有灌木、草本植物和地被物,森林和灌丛下土壤裸露,森林结构层次简单,水土保持性能差。
2 研究方法
2.1 树种选择
以生态学群落调查为基础,选取流域内22个样地中乔、灌层优势种为研究对象。研究中涉及的树种23种:云南松(P.yunnanensis)、华山松(P.armandii)、云 南 杜 鹃 (R.yunnanense)、白 栎 (Q.semicarpifolia)、马缨杜鹃 (R.delavayi)、小果南烛(L.ovalifolia)、高山栎(Q.semicarpifolia)、豆梨(P.calleryana)、川榛(C.heterophylla)、桤木(A.cremastogyne)、水红木(V.cylindricum)、大果蜡瓣花(L.chinense)、大白杜鹃(R.decorum)、菝葜(S.china)、古钟金花小檗(B.wilsonae)、贵州毛柃(E.kueichowensis)、胡颓子(E.pungens)、盐肤木(R.chinensis)、直角荚蒾(V.foetidum)、滇杨(P.calleryana)、栒子(C.horizontalis)、金丝桃(H.monogynum)、扁刺蔷薇(R.sweginzowii)。
2.2 取样及测定方法
在所确定的优势树种中,选取各年龄阶段的树种林冠上中下各摘健康叶20片,每树种选取9个重复。对取样叶片进行编号,用美国基因公司LI-3000C叶面积仪测叶片叶长(cm)、叶宽(cm)、叶面积(cm2)。奥豪丝上海公司电子天平(感重0.1g)称量每片叶鲜重(g),浸水待重力水滴尽后称叶片最大持水重(g)。叶片长度是叶片基部到叶尖的距离,叶片宽度是与叶脉垂直的最大距离,叶片最大持水率(%)是单片鲜叶的最大持水重占叶片鲜重的百分比。树皮最大持水率的计算与叶片方法一致,枝夹角是树干与一级枝的水平夹角,叶夹角是叶片与枝的水平夹角,冠幅是垂直投影的面积。实测数据见表1,表中各数据为各树种的各因子平均值。
2.3 植物特征描述
植物特征包括叶形特征和粗糙程度特征,本研究中叶形特征用叶片面积(cm2)、叶长(cm)、叶宽(cm)、长宽比、粗糙程度、叶片持水率(%)来表示,长宽比即长度和宽度的比值,可以反映叶片的宽窄程度;粗糙程度特征即叶片有无毛、毛多寡和叶片表面白粉、霜、蜡质等[17],本研究根据该特征将叶片分为低粗糙叶、中粗糙叶和高粗糙叶,粗糙度等级分别记为0,1,2。其中低粗糙叶的特征叶片双面无毛有白粉、霜、蜡,中粗糙叶的特征叶片单面有毛或少许蜡质或两面稀疏柔毛,高粗糙叶的特征叶片双面有密毛无蜡质。植物构型特征用ZJJ—枝夹角、YJJ—叶夹角;树皮特征用树皮持水率(SPCSL)来表示。

表1 树种持水因子实测数据
2.4 数据分析
采用Microsoft Excel 2007和SPSS 17.0统计分析软件对数据进行描述性统计分析、主成分分析和系统聚类分析。
3 结果与分析
3.1 优势种植物叶面积与持水能力的差异性分析
由于叶片对树种持水能力影响较大,因此对草海流域不同树种叶面积进行方差分析(如表2左下所示),结果表明,叶面积存在极显著性差异(P<0.01)的树种种对有150对,占物种叶面积总对数的68.19%;叶面积存在显著性差异(P<0.05)的树种有35对,占物种叶面积总对数的15.91%;不存在显著性差异(P>0.05)的乔木树种种对有35对,占物种叶面积总对数的15.91% 。由此可知,草海流域主要物种的叶片面积基本上都存在差异。
同时对叶片持水率进行方差分析(如表2右上所示),结果表明,叶片持水率存在极显著性差异(P<0.01)的树种种对有155对,占物种叶片持水率总对数的70.46%,其中物种1与22和其他20个种之间均存在极显著差异,物种3,4与20个种之间均存在显著性差异;叶面积存在显著性差异(P<0.05)的树种种对有26对,占物种叶片持水率总对数的11.82%;不存在显著性差异(P>0.05)的乔木树种种对有39对,占物种叶片持水率总对数的15.91%。说明在草海流域叶片持水率上是存在差异的。
3.2 植物形态特征与持水能力关系
植物形态特征与持水能力关系以树种为实体,以反映植物持水能力,反映形态特征的叶面积(Area)、叶长(Length)、叶宽(Width)、长宽比(L/W)、粗糙度(CC)、树皮持水率(SPCSL)、叶夹角(YJJ)、枝夹角(ZJJ)为属性,对表1数据进行PCA分析。
对草海流域常见种进行PCA分析,由表3可知,第三、四主成分累计贡献率达到75.60%,86.23%,有较理想的效果。表4可知,第一主成分叶面积、叶宽载荷为0.97,0.953,反映叶片面积大小;第二主成分长宽比、叶长、枝夹角载荷为0.893,0.869,0.724,反映叶片的宽窄和物种的外形,第三主成分叶片持水率的在载荷为0.921,反映叶片的持水能力,第四主成分叶片夹角的载荷为0.814,反映叶片对降雨的汇集能力。

表2 草海流域水源功能区优势植物种叶面积与持水能力方差分析
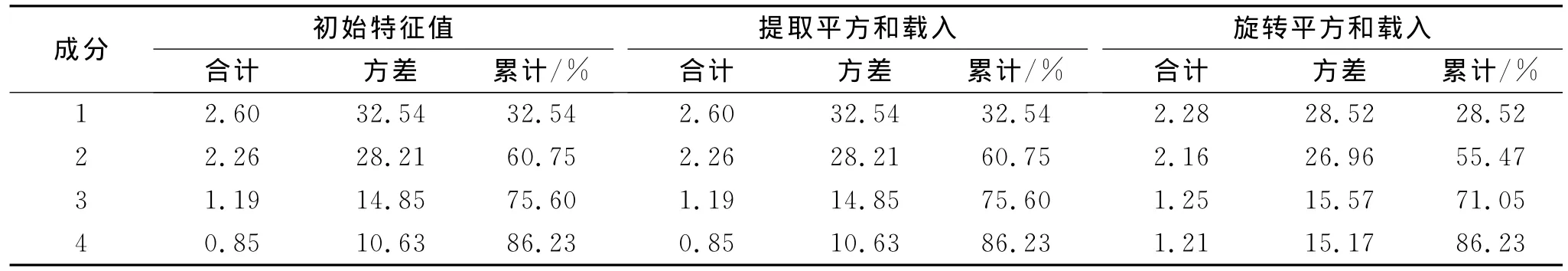
表3 PCA中解释的总方差

表4 旋转前后各因子载荷矩阵
3.3 树种综合持水能力评价
由PCA分析结果得出4个主成分分别表示叶面积大小、叶片宽窄、树木枝夹角(构型)、叶片夹角(叶片汇水能力)。采用公式计算各树种的持水能力。
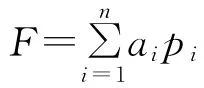
式中:F——某树种持水能力综合得分值;ai——第i个主成分特征值占百分比;pi——第i各主成分因子得分值。
由表5表明,各树种的持水能力差异显著,桤木持水能力最强,得分3.11,金丝桃最低,为-1.52。各树种持水的对策不同,将影响各自的持水能力。桤木、马缨杜鹃以叶大、叶片粗糙度高、分枝多、枝夹角大、树皮粗糙等原因来提高自身的持水能力,金丝桃持水偏低主要因叶片小且光滑无毛,植株分枝较少且叶夹角偏小等原因造成的。

表5 树种综合持水能力
3.4 植物持水功能群划分
对草海流域23各常见种进行PCA分析,得出决定植物持水功能因子主要表现为叶片大小、叶片宽窄、枝夹角(树木的构型)、叶夹角(叶片的汇水形式)4类,表5中树种综合持水能力得分并不能用于区分各树种在不同对策上获取持水能力的特点,固以树种为实体,对反映物种持水能力的4个主成分为属性进行聚类分析(图1),通过分析,将草海流域23个常见种分为5类持水功能群。其命名方法为:生活型+叶片大小+持水能力高低,如大白杜鹃、水红木、马樱杜鹃等可命名为常绿阔叶大叶片高持水功能群。
第一类PFGsI阔叶大叶高持水功能群包括川榛、桤木、马樱杜鹃、大白杜鹃、水红木;该类型功能群叶片较大,变动范围为34.0~57.10cm2,长宽比平均为2.13,枝夹角平均为45°,叶夹角平均为36.3°,综合得分0.94~3.11。
第二类PFGsII阔叶中叶中持水功能群,叶面积平均为14.42cm2,长宽比1.69,枝夹角为48.6°,叶夹角为33°,持水能力综合得分-0.51~0.24。主要物种有高山栎、胡颓子、小果南烛、滇杨、大果蜡瓣花。
第三类PFGsIII阔叶小叶中持水功能群,叶面积较小,平均在4.13cm2,长宽比为2.22,枝夹角为30°,叶夹角为5°,综合得分-0.53~-1.5。主要物种有扁刺蔷薇、栒子、云南杜鹃、直角荚蒾。
第四类PFGsIV针叶中持水功能群,华山松、云南松为针叶树种,综合得分-0.17~-0.72。
第五类PFGsV阔叶小叶低持水功能群,叶面积平均4.47cm2,长宽比为2.55,枝夹角为40°,叶夹角为26.3°,持水能力综合得分-1.09~-1.52。主要物种为豆梨、古钟金华小檗、贵州毛柃、金丝桃。

图1 不同树种系统聚类图
4 结论与讨论
(1)本研究区树种叶片持水能力主要受叶片大小、叶片宽窄、叶片的粗糙程度、枝夹角、叶夹角等形态特征影响,因此,通过上述5个植物形态指标,对草海流域水源功能区植物群落中23个主要树种的持水能力进行综合评价和能力排序。这种将植物形态特征与持水能力结合,采用定性与定量结合的测定方法,较以往仅通过树种叶片持水率分析持水能力更为客观,但对针叶树种的叶片形态与功能的关系研究缺乏,有待进一步探讨。
(2)草海流域水源功能区22个植物群落中主要树种分为5个叶片持水功能群,即阔叶大叶高持水功能群(PFGsI)、阔叶中叶中持水功能群(PFGsII)、阔叶小叶中持水功能群(PFGsIII)、针叶中持水功能群(PFGsIV)、阔叶小叶低持水功能群(PFGsV),将为研究区涵养水源林的营造和生态修复过程中的树种选择、结构优化提供理论依据。
(3)本文利用植物形态特征与植物持水能力的关系,开展了植物持水功能群的划分研究,即通过植物功能性状研究,揭示相应的功能。其结果既能得出植物群落中各树种的持水能力,又能揭示出各植物持水能力依据的主要形态对策,如阔叶大叶高持水功能群(PFGsⅠ)的桤木、川榛等,以大叶片、大枝(叶)夹角和树皮的高持水能力为对策使其植株持水能力最高。利用这种主要形态对策可用于指导树种选择,也为植物群落持水结构配置提供良好的依据,具有现实意义。利用叶片和植物形态特征进行叶片持水功能群研究,丰富了植物功能群研究的内容,也是一种可行的方法学探索,具有理论意义。但要全面评价树种的个体持水能力,还需从树冠构型、树干、树枝、枯落物等影响持水能力的形态指标与树种持水关系的深入研究,才能更好划分树种持水功能群。
[1] 杨晓慧,鲍雅静,韩国栋,等.植物功能群及其在生态学研究中的应用[J].大连民族学院学报,2009,11(5):397-400,409.
[2] 孙国钧,张荣,周立.植物功能多样性与功能群研究进展[J].生态学报,2003,23(7):1430-1435.
[3] 岳静,郝敏,张灵菲,等.物种多样性—生态系统功能关系的影响因素[J].草业科学,2011(04):532-535.
[4] 谭丽萍,周广胜.内蒙古羊草群落、功能群、物种变化及其与气候的关系[J].生态学报,2013,33(2):0650-0658.
[5] 邓福英,臧润国.海南岛热带山地雨林天然次生林的功能群划分[J].生态学报,2007,27(8):3240-3249.
[6] 冯秋红,史作民,董莉莉.植物功能性状对环境的响应及其应用[J].林业科学,2008,44(4):125-131.
[7] 胡楠,范玉龙,丁圣彦,等.陆地生态系统植物功能群研究进展[J].生态学报,2008,28(7):3302-3311.
[8] 鲍雅静,李政海.基于能量属性的植物功能群划分方法探索:以内蒙古锡林河流域草原植物群落为例[J].生态学报,2008,28(9):4540-4546.
[9] 毛伟,李玉霖,张铜会,等.不同尺度生态学中植物叶性状研究概述[J].中国沙漠,2012,32(1):33-38.
[10] 白永飞,张丽霞,张焱,等.内蒙古锡林河流域草原群落植物功能群组成沿水热梯度变化的样带研究[J].植物生态学报,2002,26(3):308-316.
[11] 赵建中,彭敏,刘伟,等.矮嵩草草甸不同功能群主要植物种生长特征与地表温度的相关性分析[J].西北植物学报,2012,32(3):559-565.
[12] 马建军,姚虹,冯朝阳,等.内蒙古典型草原区3种不同草地利用模式下植物功能群及其多样性的变化[J].植物生态学报,2012,36(1):1-9.
[13] 蔺雨阳,王得祥,雷瑞德,等.秦岭中段天然油松林生态种对和功能群划分[J].东北林业大学学报,2009,37(11).46-51.
[14] 毛培利,臧润国,丁易,等.海南岛热带山地雨林三种不同演替阶段典型植物苗期光合特性研究[J].林业科学研究,2011,24(5):563-571.
[15] 孔凡翠,杨瑞东,林树基.从威宁草海的演化分析0.73Ma来贵州威宁地区喀斯特环境的演变[J].地球与环境,2010(2):138-145.
[16] 齐建文,李矿明,黎育成,等.贵州草海湿地现状与生态恢复对策[J].中南林业调查规划,2012,31(2):39-40.
[17] 中国科学院植物研究所.中国高等植物图鉴[M].北京:科学出版社,1975.
