Temporal changes in nitrogen acquisition of Japanese black pine (Pinus thunbergii) associated with black locust (Robinia pseudoacacia)
2014-04-19LopezMizotaNoboriSasakiYamanaka
M.L.Lopez C.• C.Mizota • Y.Nobori • T.Sasaki • T.Yamanaka
Introduction
The nitrogen-fixing tree black locust (Robinia pseudoacacia) is the second most abundant deciduous tree species in the world (Malcolm et al.2008).This competitive, early-successional species native to the Appalachian uplands in USA has rapidly invaded several countries (Malcolm et al.2008).Nitrogen-fixing species can impact the structure and function of plant communities by contributing a high annual budget of N-rich litter of up to 100 kg·ha-1·a-1(Binckley et al.1985; Moon and Haruki 1999; Baddey et al.2000; Schulze 2000; Rice et al.2004).Invasion of these species elevates soil N concentration and increases rates of subsequent N cycling, including mineralization and nitrification (and probably denitrification) in forest ecosystems (Binckley et al.1985; Kawata 1987; Shulze et al.1991; Rice et al.2004; Malcolm et al.2008).
In Japan, black locust was first introduced in the middle of the 19th century to stabilize Japanese black pine (Pinus thunbergii) plantations on coastal sand dunes along the Sea of Japan where N-poor habitat prevails.The legume species has recently invaded gaps caused by the spread of pine nematode (Taniguchi et al.2007).Taniguchi et al.(2007) reported that the invasion reduces N acquisition by the associated black pine due to elevated concentration of nitrate-N.Temporal changes in the process of N acquisition by black locust and associated black pine are expected to be recorded in the annual rings and to represent past interactions (Poulson et al.1995; Sauer et al.2004; Savard 2010).
Use of the naturally abundant N isotope composition (δ15N/14N ratio), commonly designated as δ15N, (a per mil variation relative to atmospheric nitrogen, where δ15N=0) has been recognized as a method of tracing N nutrition in studies of forest ecosystems (Natelhoffer and Fry 1988; Gebauer and Shulze 1991; Robinson 2001; Hart and Classen 2003; Lopez et al.2010; Mizota et al.2011).Under humid and temperate climatic conditions, δ15N values for wood of black locust are expected to be around zero per mil because limited N isotope fractionation results from biological nitrogen fixation (Robinson 2001).Black pine grown in humid and temperate climates such as those of central to southern Japan has δ15N values in a narrow range for any geographic region.This reflects the sole N source (NH4+and NO3--N) in meteoric precipitation in open soil systems (Lopez et al.2010; Mizota et al.2011).In forest gaps newly formed after nematode disease outbreaks, e.g.those in forests along the Sea of Japan coast, black locust invades, resulting in the formation of mixed forests (Taniguchi et al.2007).In any forest ecosystem, when other nutrients are not limiting, increases in N availability can enhance tree growth (Bouillet et al.2008).Black locust is a fast growing species with high biomass production.Because of its N fixation ability it can be used to reduce the demand for external fertilization, especially if the timing for maximum N transfer from black locust to black pine is determined.The aim of this study was to investigate N acquisition by Japanese black pine associated with black locust by using δ15N analyses of annual tree rings (resolution of one year) at two locations in Japan.We discuss temporal changes in δ15N in relation to the growth of associated N-fixing legumes.
Materials and methods
Description of the study sites
Two study sites were selected to represent different durations of black locust establishment in mixed forest with black pine.Recently invaded forest was represented by the Shohnai site in northeast Japan, while longer black locust presence was represented by the Kita-Kyushu site in southwest Japan.Soils at both study sites were classified as Dystric Regosols (FAO/UNESCO system) developed on coastal dune sands.Mean annual air temperature and annual rainfall at the nearest meteorological stations (Sakata and Fukuoka, respectively) are 12.1 and 16.2°C, and 1857 and 1604 mm, respectively.A brief description of the sites is given below.
Shohnai site
This study site is representative of temperate-humid regions in Yamagata Prefecture, northeast Japan.The Shohnai (38°49'14"N, 139°47'47"E) coastal areas in northern Yamagata are subject to strong seasonal winds during winter.For more than 150 years, black pine has been planted along the coast to prevent sand movement, which causes serious damage to agricultural crops in back-dune areas.The site selected for sample collection was located on the west-facing slope of a coastal sand dune where black locust recently (early 1990s) invaded clearings in black pine forests.The growth of black locust is faster than that of the associated black pine (Kawata 1987; Taniguchi et al.2007) and contributes to the increase of N input to soils, providing a new N source for the surrounding black pine.The densities of Pinus thunbergii and Robinia pseudoacacia on the study area was estimated at 2,100 and 2,400 trees per hectare, respectively.Kita-Kyushu site
This study site represents the warm and humid regions in Fukuoka in southwest Japan.The site (33°39'53" N, 130°21'21" E) was along the Genkai Sea at the location of the flat or nearly flat National Reserve of Marine Park.This area was also invaded by black locust after clearings formed in black pine forests infested by pine nematodes, but invasion occurred about 20 years earlier here (early 1970s) than at the Shohnai site (Gyokusen et al.1991).At this site, decaying large trunks of black locust are common, suggesting that black locust has occurred here for at least two generations.At this site there was no pure black locust stand.The densities of P.thunbergii and R.pseudoacacia during the present study site were estimated at 1760 and 4750 trees per hectare, respectively (Gyokusen et al.1991).
Selection of annual tree ring samples
Representative trees were carefully selected based on observations of their spatial distribution.Tree cores were collected at both sites from Japanese black pine (soft wood) and black locust (porous wood) trees, using an increment borer (diameter of 12 mm, Haglof, Langsele, Sweden).Core samples were collected as follows: One core sample from a pure black pine stand, three core samples from three black pine trees in a mixed forest with black locust (at the Kita-Kyushu site only one sample was collected) and one core sample from a pure black locust stand.The increment borer was inserted into the tree from one side to the other side of the trunk (thus two sides were collected at once with the pith included).All samples were collected at breast height (1.3 m above ground level).The cores were taken to a laboratory for sampling of annual tree-rings (resolution of one year) to quantify δ15N values.One of the core samples from black pine in the mixed forest was analyzed from 1992 while the other two were analyzed from 1999 to 2009.The tree ring series ranged from the first year (1990/1992) to 2009 except for the sample from a pure black pine stand from Shohnai that was approximately 42 years old.All analyzed tree rings corresponded to the sapwood area.Each tree ring was systematically dissected using a sharp, high-quality, stainless blade, and dried at 55 °C overnight.Each ring sample was then cut into small pieces using a nail cutter.
Nitrogen isotope analysis
Extraction of labile N was not carried out because there were negligible changes in δ15N values in tree rings after extraction of labile N in previous studies (Couto-Vazquez and Gonzales Prieto 2010; Doucet et al.2011; Lopez et al.2011).δ15N values for dry ring samples were estimated using a CF-IRMS (continuous-flow type mass spectrometer: Iso Prime mass spectrometer, GV Instruments, UK) installed at the Faculty of Science, Okayama University.The evolved gas was first passed through a column packed with solid CaO-NaOH reagent (soda lime) to eliminate excess CO2and then subjected to gas chromatography to separate N2followed by mass spectrometric measurement of15N/14N ratios.The isotopic compositions of samples were expressed relative to atmospheric N2(δ15N=0) on scales normalized to the known δ15N values of laboratory working standards for glycine (δ15N=-0.3‰), which was normalized to L-glutamic acid distributed as USGS-40 (δ15N=-0.2‰) by SI Science Inc., Japan.The working standard was analyzed after every eight to ten samples during CF-IRMS runs to assess the replicability of the isotope measurements and normalization.One pulse of pure N2reference gas from a tank reservoir (δ15N=-2.5‰) was discharged into the IRMS at the beginning of each chromatogram for both standards and samples.The accuracy obtained for standards and samples during the overall analytical procedure was better than ±0.2‰for sample sizes of ≥10 µg N.
Results and discussion
Temporal changes in tree ring nitrogen isotope composition
It is well documented that tree-rings of Japanese black pine retain their intrinsic record of N acquisition (Lopez et al.2010; Mizota et al.2011; Lopez et al.2011).Temporal changes in nitrogen isotope composition of tree-rings grown after 1992/1994 from Shohnai and Kita-Kyushu sites are shown in Fig.1 and 2, respectively.Clear differences were recorded for δ15N values of black locust and black pine.

Fig.1: Temporal changes in δ15N values of annual tree-rings from Shohnai site.The standard deviation of black pine in the mixed stand corresponds to three core samples per tree-ring.Higher variation in δ15N value of ring samples from black pine, relative to those of black locust is also noticeable.For detailed explanation of the plots described in the legends, see the section Materials and methods.
Shohnai site
δ15N values of black locust tree rings from the pure stand varied in a narrow range from -1.1‰ to -0.5‰ (average= -0.9‰, n=18).Such values approximate those of atmospheric N2(δ15N= 0).Because limited N isotope fractionation is associated with N fixation (Robinson 2001), this result confirms the substantial contribution by N fixation in this woody legume.δ15N values of tree rings from the pure black pine stand ranged from -5.3‰ to -2.3‰, averaging -3.9‰.Nitrogen isotope values were similar to those of inorganic nitrogen input from meteoric precipitation as observed at a nearby site (δ15N=-5.5‰ to -1.7‰) (Fukuzaki and Hayasaka 2009) during May 2001 to November 2002.
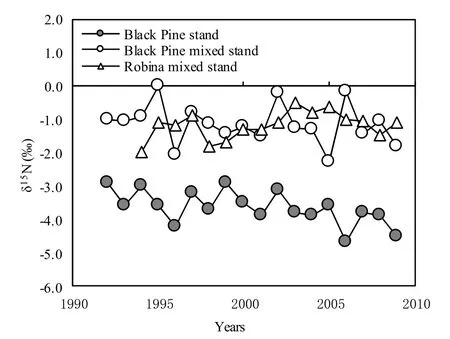
Fig.2: Temporal changes in δ15N values of annual tree-rings from Kita-Kyushu site.The δ15N values of ring samples from black pine are more variable, relative to those of black locust.For detailed explanation of the plots described in the legends, see the section Materials and methods.
δ15N values for tree rings from black pine in the mixed stand were markedly higher than those from the pure stand.A steady increase was observed from 1992 (-2.6‰) to 2009 (-2.1‰±0.6‰).Tree ring δ15N values in 1999 and 2004 were (-3.2±1.4)‰ and (-1.4±0.4)‰, respectively.The δ15N values for 2002 to 2009 approached those of the associated black locust.
δ15N values of tree rings of black pine for the first and second years (1992 to 1993) in the mixed stand showed clearly higher values (-2.7‰ and -2.6‰) than for the corresponding years for the pure black pine stand (-4.5‰ and -4.3‰).This could indicate that somewhat elevated δ15N values resulted from the N input from past invasion of black locust prior to the present stand of Japanese black pine (Fig.1).
Kita-Kyushu site
The overall temporal trend of δ15N values at the Kita-Kyushu site was different from that observed for the Shohnai site.From 1994, δ15N values of black locust tree rings in the mixed stand ranged from -2.0‰ to -0.5‰, averaging -1.2‰.
δ15N values for black pine tree rings from a pure stand ranged from -4.7‰ to -2.9‰ for the period 1992−2009, averaging -3.7‰.These values were nearly identical to those for the Shohnai site (δ15N=-3.9‰).In contrast, δ15N values for tree rings from black pine in the mixed stand showed higher values that ranged from -2.3‰ to 0 and averaged -1.2‰ for the entire growth period, suggesting incorporation of N derived from N2 fixation from the very first year of Japanese black pine establishment in this area.This indicates that several generations (more than 30 years) of black locust in this area have enriched soil N to the point that it is readily available to newly established Japanese black pine (Fig.2).
Potential use of nitrogen stable isotope composition in analysis of the acquisition of nitrogen in mixed forests
From the temporal fluctuations and elevated isotope values observed in tree rings of black pines on both sites, it is clear that the main cause of this change is the acquisition of N derived from N2fixation through association with black locust.To a lesser degree, however, the roles of processes such as mineralization, nitrification and denitrification must also be considered.
It is well known that woody legumes fix variable amounts of atmospheric N2.Estimates range from nearly 0 to 204 kg·ha-1·a-1(Bouillet et al.2008).The highest value reported for black locust was 110 kg·ha-1·a-1(calculated contribution of atmospheric N2relative to whole N absorbed from the environment = 90%) (Danso et al.1955).The nitrogen isotope analysis for annual tree rings can be applied to forest management practices in which woody legumes are mixed with other species and, as in the case of this study, can enhance growth of black pine forests that provide windbreaks in coastal areas along the Sea of Japan.However, the competition between the two species could counterbalance the positive effect of black locust on the growth of black pine as reported for red alder-Douglas fir forest (Binckley et al.1985).Malcolm et al.(2008) reported that black locust was completely removed prior to the reestablishment of pine-oak forest because of the enhanced growth of these species to soils enriched by high levels of N resulting from the prior presence of black locust.However, this is apparently not the case for black pine since it was not affected by high levels of N in the soil in forest receiving high input of avian N (Lopez et al.2011; Mizota et al.2011).
Taniguchi et al.(2007) reported that black pine relies on ectomycorrhizal (ECM) colonization for N uptake from the soil.Since the ECM community changes because of N enrichment in the soil, the N enrichment effect of black locust on black pine trees is negative.Nevertheless, black pine trees exposed to high inputs of avian N grow without limitation in other areas of Japan (Lopez et al.2011; Mizota et al.2011), suggesting that even when ECM symbiosis is modified, black pine trees can uptake available N directly from the soil.Changes in ECM functions related to increases in N input remain, however, a topic for future studies.
Our study results contribute to understanding the positive role of black locust in forest management practices for black pine in coastal areas of the Sea of Japan.Elevation of δ15N values in tree rings of Japanese black pine resulting from biological N fixation from associated black locust saturates within ca.10 years after the black locust invasion (Fig.1).This estimate enables improved scheduling of the removal of black locust during the early growth stages of black pine plantations.Such practices have not yet been implemented, despite the role of black locust in N fixation under low soil fertility conditions.
Acknowledgement
We are grateful to two anonymous reviewers for providing constructive comments and to Dr A.L.Cronin for improving the English in the paper.
Boddey RM, Peoples MB, Palmer B, Dart P.2000.Use of the15N natural abundance technique to quantify biological nitrogen fixation by woody perennials.Nutr Cycl Agroecosys, 57: 235−270.
Bouillet JP, Laclau JP, Gonçalves JLM, Moreira MZ, Trivelin PCO, Jourdan C, Silva EV, Piccolo MC, Tsai SM, Galiana A.2008.Mixed-species plantations of Acacia mangium and Eucalyptus grandis in Brazil.2: Nitrogen accumulation in the stands and biological N2fixation.For Ecol Manag, 255: 3918−3930.
Couto-Vazquez A, Gonzalez-Prieto SJ.2010.Effects of climate, tree age, dominance and growth on δ15N in young pinewoods.Trees, 24: 507−514.
Danso SKA, Zapata F, Awonaike KO.1955.Measurement of biological N2fixation in field-grown Robinia pseudoacacia L.Soil Biol Biochem, 27: 415−419.
Doucet A, Savard MM, Begin C, Smirnoff A.2011.Is wood pre-treatment essential for tree-ring nitrogen concentration and isotope analysis? Rapid Commu Mass Spectr, 25: 469−475.
Fukuzaki N, Hayasaka H.2009.Seasonal variations of nitrogen isotope ratios of ammonium and nitrate in precipitations collected in the Yahiko-Kakuda mountains area in Niigata prefecture, Japan.Water, Air, Soil Pollut, 203: 391−397.
Gebauer G, Schulze ED.1991.Carbon and nitrogen isotope ratios in different compartments of a healthy and declining Picea abies forest in the Fichtelgebirge, NE Bavaria.Oecologia, 87: 198−207.
Gyokusen K, Iijima Y, Yahata H.1991.Spatial distribution and morphological features of root systems in Niseakashia (Robinia pseudo-acacia L.) growing under a coastal black pine forest.Bull Kyushu Univ Forests, 64: 13−28.(In Japanese)
Hart SC, Classen AT.2003.Potential for assessing long-term dynamics in soil nitrogen availability from variations in δ15N of tree rings.Isotopes Environ Health Stud, 39: 15−28.
Kawata H.1987.Difference of nutrient amounts returned to soil by litter falls and their effects on soil properties between Pinus thunbergii and Robinia pseudoacacia forests in coastal sand dune.Bull Niigata Univ For, 20: 51−56.(In Japanese)
Lopez CML, Mizota C, Yamanaka T, Nobori Y.2010.Temporal changes in tree-ring nitrogen of Pinus thunbergii trees exposed to Black-tailed Gull (Larus crassirostris) breeding colonies.Appl Geochem, 25: 1699−1702.Lopez CML, Mizota C, Yamanaka T, Nobori Y.2011.Effects of pre-treatment on the nitrogen isotope composition of Japanese black pine (Pinus thunbergii) tree-rings as affected by high N input.Rapid Commu Mass Spectr, 25: 3298−3302.
Malcolm GM, Bush DS, Rice SK.2008.Soil nitrogen conditions approach preinvasion levels following restoration of nitrogen-fixing black locust (Robinia pseudoacacia) stands in a pine-oak ecosystem.Restor Ecol, 16: 70−78.
Mizota C, Lopez CML, Yamanaka T, Nobori Y.2011.Differential response of two Pinus spp.to avian nitrogen input as revealed by nitrogen isotope analysis for tree-rings.Isot Environ Health Stud, 47: 62−70.
Montagnini F, Haines B, Boring L, Swank W.1986.Nitrification potentials in early successional black locust and in mixed hardwood forest stands in the southern Appalachians, USA.Biogeochemistry, 2: 197−210.
Moon HS, Haruki M.1999.Nutrient distribution and content in plant tissue, forest floor, and litterfall in four pioneer stands on volcano Mt.Showa-Shinzan.J For Environ, 41: 7−11.
Natelhoffer KJ, Fry B.1988.Controls on natural nitrogen-15 and carbon-13 abundances in forest soil organic matter.Soil Sci Soc Am J, 52: 1633−1640.
Poulson SR, Chamberlain CP, Friedland AJ.1995.Nitrogen isotope variation of tree rings as a potential indicator of environmental change.Chem Geol (Iso.Geosci.Sec.), 125: 307−315.
Rice SK, Westerman B, Federici R.2004.Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen-cycling in a pine-oak ecosystem.Plant Ecol, 174: 97−107.
Robinson D.2001.δ15N as an integrator of the nitrogen cycle.Trends Ecol Evol, 16: 153−162.
Sauer M, Cherubini P, Ammann M, De Cinti B, Siegwolf R.2004.First detection of nitrogen from NOxin tree rings: a15N/14N study near a motorway.Atmosp Environ, 38: 2779−2787.
Savard MM.010.Tree-ring stable isotopes and historical perspectives on pollution – An overview.Environ Pollu, 158: 2007−2013.
Schulze ED, Gebauer G, Ziegler H, Lange OL.1991.Estimates of nitrogen fixation by trees on an aridity gradient in Namibia.Oecologia, 88: 451−455.
Schulze ED.2000.Carbon and nitrogen cycling in European forest ecosystems.Ecological Studies, 142: 3−13.
Taniguchi T, Tamai S, Yamanaka N, Futai K.2007.Inhibition of the regeneration of Japanese black pine (Pinus thunbergii) by black locust (Robinia pseudoacacia) in coastal sand dunes.J For Res, 12: 350−357.
杂志排行

Journal of Forestry Research的其它文章
- Ethno-medicinal plants used by Bengali communities in Tripura, northeast India
- Litter production, decomposition and nutrient mineralization dynamics of Ochlandra setigera: A rare bamboo species of Nilgiri Biosphere Reserve, India
- Plant diversity at Chilapatta Reserve Forest of Terai Duars in subhumid tropical foothills of Indian Eastern Himalayas
- Floristic composition and management of cropland agroforest in southwestern Bangladesh
- The changing landscape of mangroves in Bangladesh compared to four other countries in tropical regions
- The effect of fire disturbance on short-term soil respiration in typical forest of Greater Xing’an Range, China