不同光照条件下三七幼苗形态及生长指标的变化
2014-04-09匡双便张广辉陈中坚魏富刚杨生超陈军文
匡双便, 张广辉, 陈中坚, 魏富刚, 杨生超, 陈军文,①
(1. 云南农业大学: a. 云南省优势中药材规范化种植工程研究中心, b. 农学与生物技术学院, 云南 昆明 650201;2. 文山学院三七研究院, 云南 文山663000;3. 云南文山苗乡三七实业有限公司, 云南 文山 663000)
五加科(Araliaceae)人参属(PanaxLinn.)多年生草本植物三七〔Panaxnotoginseng(Burk.) F. H. Chen〕的根、茎、叶和花均可入药,具有散瘀止血、消肿定痛的功效[1],还具有抗血栓、抗心肌缺血、抗心律失常、抗炎、抗衰老及抗肿瘤等作用[2]。三七的药用及栽培历史悠久,历来就有“北人参、南三七”的说法[3]。三七主要分布于云南和广西交界及相邻的一些地区,主产于云南文山州的文山、砚山、丘北和马关等地,种植面积和产量分别占全国三七总种植面积和总产量的98%以上[4-5],已成为文山州乃至云南省最具特色和最具竞争力的生物支柱产业[6]。
光是影响植物生态建成的最重要的环境因子,随光照环境的变化植物会发生一系列的适应性改变[7],而植物宏观形态学特征对光的适应性表现最易观察也最直观。当环境中的光照强度降低时,植物地上部分的形态特征会朝着有利于捕获更多光能的方向变化,例如植株冠幅增大、单叶面积增加、分枝数减少、单株叶片总数减少等[8-10];环境中的光照强度对植物地下部分的形态发生和生长也有影响[11-13]。
三七是典型的喜阴药用作物[14],应在遮阳条件下培育,且荫棚透光度对三七幼苗的生长及发育均有影响[15]。一般来说,在30%自然光照条件下三七才能正常生长发育,故三七荫棚有“三成透光,七成蔽荫”之说;而在三七产区从事种植技术研发的科技人员认为:遮阳棚透光率为8%~12%最适宜,透光率超过17%对三七生长不利。因此,荫棚透光度的合理调整是三七种苗培育过程中的一个关键环节。为此,作者对在透光率不同的荫棚下三七幼苗的生长特性及干物质积累和分配特征进行研究,以期为三七良种育苗过程中适宜光照条件的设置提供实验依据。
1 研究地自然概况和研究方法
1.1 研究地自然概况
实验在云南省文山州砚山县苗乡三七实业有限公司三七科技示范园内进行。砚山县位于云南东南部,地理坐标为东经103°35′~104°45′、北纬23°18′~23°59′,海拔1 539 m。属亚热带大陆性季风气候[16],气候温和,夏无酷暑、冬无严寒、春暖夏凉;年降雨量1 200 mm,年均温16 ℃,最冷月均温9 ℃,最热月均温21 ℃,全年无霜期300 d以上,日均温≥10 ℃的积温为5 000 ℃。砚山县是三七的主要传统产区,也是三七最适宜的产区之一。
1.2 研究方法
1.2.1 实验设计及处理 在三七产区,用玉米秸秆搭建的荫棚透光率为1.0%~2.0%,用杉树枝叶搭建的荫棚透光率为3.0%~5.0%。根据作者前期的调查研究结果,将本实验所用荫棚的透光率分别设置为自然全光照的22%、12%、8%、3%和1%。其中,1%透光率为极端低光照水平,相当于部分农户用玉米秸秆搭建的荫棚透光率;3%透光率相当于部分农户用杉树枝叶搭建的荫棚透光率;8%和12%透光率分别代表当前专业科技工作者认定的三七种植的最适透光率;而22%透光率则代表三七幼苗能忍受的高光照水平。
采用三七专用遮阳网搭建上述5个不同透光率的荫棚,每个荫棚长4 m、宽2 m、高2 m。荫棚内光照强度用LI-1400型光量子数据采集器(美国LI-COR公司)测量,在晴天从08:00到18:00 1 min记录1次数据;同一时刻荫棚内光照强度与荫棚外光照强度的比值即为透光率,全天所有时刻透光率的平均值即为荫棚的透光率。
于2013年1月31日将三七种子用多抗霉素浸种15 min,然后播种于育苗盘中,播种基质为沸石、甘蔗渣和壤土按体积比5∶ 3∶ 2的比例混合而成的混合基质。播种后将育苗盘移至不同透光率的荫棚下,实时监管,使三七植株免受病害、虫害和草害;荫棚内安装自动喷灌系统,实施正常水分管理。
1.2.2 不同指标的测定 于2013年10月12日,在三七幼苗膨大后期随机从各处理组中挖取15株单株,洗净。用电子数显游标卡尺(精度0.01 mm)分别测量块根直径(根膨大处直径)和茎基径;用直尺分别测量株高、冠幅、主根长(块根和须根的垂直自然根长)、块根长(从芽眼至筋条变小处的长度)、单株须根长(每一单株须根长度的总和)、每片小叶的长度和宽度(小叶最长处和最宽处的长度),并计算三七单株叶面积(将叶片按椭圆形进行计算),统计叶片数和根数等形态指标。
将根、茎和叶片分开并置于烘箱中,于105 ℃杀青1 h后于60 ℃烘干至恒质量;用电子天平(精度0.001 g)称取块根干质量、须根干质量、根总干质量(块根和须根干质量之和)、茎干质量和叶片干质量,并计算植株总干质量、根冠比(根干质量与茎和叶片干质量之和的比值)、根质比(根干质量与植株总干质量的比值)、茎质比(茎干质量与植株总干质量的比值)、叶质比(叶片干质量与植株总干质量的比值)及比叶面积(叶面积与干质量的比值)等。
从每个处理组中选取长势较好的植株30株,用SPAD 502 Plus叶绿素仪(日本KONICA MINOLTA公司)测定叶绿素相对含量(SPAD)。
1.3 数据处理
采用SPSS 14.0统计分析软件进行单因素方差分析(P≤0.05),用Sigma Plot 10.0 软件绘图。
2 结果和分析
2.1 对三七植株形态指标的影响
不同透光率对三七幼苗植株形态指标的影响见表1。由表1可以看出:在透光率22%的条件下三七幼苗株高最大,且显著(P≤0.05)高于透光率8%和1%处理组,但与透光率12%和3%处理组差异不显著(P>0.05)。在透光率3%条件下三七幼苗的冠幅最大,且与透光率22%和1%处理组有显著差异。在透光率22%条件下三七幼苗块根长度最大,与透光率8%处理组差异不显著,但与其他处理组有显著差异,且随透光率降低呈现逐渐减小的趋势。在透光率8%条件下三七幼苗的主根长及茎基径均最大,但与其他处理组差异不显著。在透光率22%和12%条件下三七幼苗块根直径显著大于透光率8%、3%和1%处理组,且随透光率降低块根直径逐渐减小。在透光率22%条件下三七幼苗的单株须根数和单株须根长均最大,并与其他处理组有显著差异,且随透光率降低呈逐渐减小的趋势。另外,在透光率22%条件下叶片明显皱缩(数据未显示)。

表1 不同透光率对三七幼苗植株形态指标的影响
2.2 对三七植株不同器官干质量及分配的影响
2.2.1 对不同器官干质量的影响 不同透光率对三七幼苗单株不同器官干质量的影响见表2。由表2可见:透光率对各器官的干质量和植株总干质量有显著影响,且随透光率增加各器官干质量和植株总干质量依次增大。在透光率22%条件下,各器官干质量和植株总干质量均最大,且除须根干质量外,各器官干质量及植株总干质量均显著高于其他处理组;而在透光率1%条件下,各器官干质量及植株总干质量均显著小于其他处理组。
2.2.2 对不同器官干质量分配的影响 不同透光率对三七幼苗不同器官干质量分配的影响见表3。结果显示:在透光率8%条件下,根冠比和根质比均最大,且与透光率22%和12%处理组无显著差异,但均显著大于透光率3%和1%处理组。在透光率1%条件下,茎质比最大且与其他处理组差异显著;而透光率3%处理组的茎质比也较大,显著大于透光率22%、12%和8%处理组。在透光率1%条件下叶质比最大,显著大于其他处理组;在透光率3%条件下叶质比也较大,并显著大于透光率22%和8%处理组。
表2 不同透光率对三七幼苗单株不同器官干质量的影响
表3 不同透光率对三七幼苗不同器官干质量分配的影响
2.3 对三七幼苗单株叶面积和比叶面积的影响
不同透光率对三七幼苗单株叶面积和比叶面积的影响见表4。结果显示:在透光率1%条件下单株叶面积最小,且与其他处理组差异显著;而其他4个处理组的单株叶面积无显著差异,但在透光率3%条件下单株叶面积最大。随透光率增大比叶面积依次减小;其中,在透光率1%条件下比叶面积最大,其次为透光率3%处理组,均显著高于其他3个处理组;但在透光率22%、12%和8%条件下单株比叶面积无显著差异。
2.4 对三七幼苗叶片叶绿素相对含量的影响
不同透光率对三七幼苗叶片叶绿素相对含量(SPAD)的影响见图1。结果表明:在透光率不同的条件下三七叶片的SPAD值有明显差异。在透光率1%、3%和8%条件下三七叶片的SPAD值均显著大于透光率22%和12%处理组,其中,在透光率8%条件下SPAD值最大,而在透光率22%条件下三七叶片的SPAD值最小。
表4 不同透光率对三七幼苗单株叶面积和比叶面积的影响
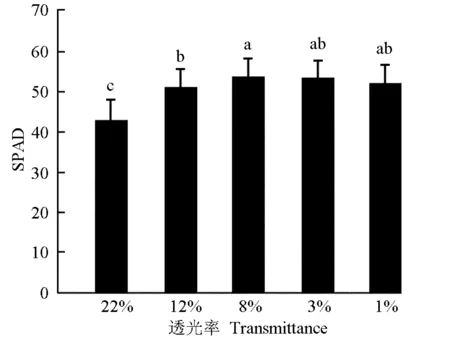
图中不同的小写字母表示各处理间差异显著(P≤0.05) Different small letters in the figure indicate the significant difference (P≤0.05) among different treatments.
3 讨论和结论
光照对三七生长发育有极大影响,三七的生长、产量及病虫害发生状况与荫棚的透光率有密切关系[5]。在光照强度受限时,植物通过增加株高向高空伸展,增加叶片光合面积以截获更多光能。在透光率3%条件下,三叶鬼针草(BidenspilosaLinn.)的株高显著高于全光照条件下[17];对于生长在不同光照环境下的麻栎(QuercusacutissimaCarr.)和刺槐(RobiniapseudoacaciaLinn.)幼苗,极度弱光环境限制了它们的株高、基径、总叶面积、叶面积指数和总叶数等形态指标的增长,但与全光照环境相比,适度遮阳有利于植株的形态生长[18]。本研究结果也证实:光对三七幼苗的形态指标有明显影响;随透光率降低,块根直径减小、须根数和须根长也减少且差异显著;在透光率8%条件下主根长及茎基径最大;在透光率22%条件下株高最大;而冠幅则在透光率3%条件下最大。总体上看,三七作为一种典型的喜阴植物,适当遮阳有利于其生长;透光率降低使其植株的外观形态朝着有利于捕获更多光能的方向变化。
光是植物合成有机物的能源,光照强度直接影响植物的光合特性及干物质形成[19]。孟金柳等[20]认为:高光照水平下樟树〔Cinnamomumcamphora(Linn.) Presl〕植株具有较大的根干质量分配比例,低光照水平下则具有较大的叶干质量分配比例,而中度光照条件下根和叶干质量分配比例则位于两者之间。因此,在荫蔽条件下植株的叶生物量相应增加、根生物量相应减少,导致叶质比较大、根质比较小。徐飞等[18]的研究结果表明:随光照强度减弱麻栎各器官的干质量积累降低,而刺槐各器官干质量积累和分配则在适度荫蔽条件下达到最优;最优的干质量分配方式是保证捕获的光能恰好满足光合作用需要。在本实验中,随透光率降低三七根、茎和叶的干质量积累呈逐渐减小的趋势,且在透光率3%和1%条件下明显受到抑制;以透光率8%为临界点,随透光率降低根冠比及根质比减小、茎质比和叶质比增加。说明随透光率降低三七幼苗的生长策略是将干质量优先分配到茎和叶,以捕获更多的光能克服光资源限制。
比叶面积(SLA)是表征植物生长过程中碳收获策略的关键叶性状指标[21-22], SLA的大小是衡量叶片获取光能的指标[23]。一般来说,环境中光照越弱、叶片越薄、单位面积干质量越小,SLA越大,所以SLA与荫蔽程度有关。SLA随光照环境的变化而改变,在强光下减少光抑制、降低叶温、增加水分利用效率;在弱光下增加光能捕获及利用效率等[18]。李明财等[24]对东北次生林主要树种的研究结果表明:随环境中光照强度的减弱,红松(PinuskoraiensisSieb.et Zucc.)、黄檗(PhellodendronamurenseRupr.)、胡桃楸(JuglansmandshuricaMaxim.)和水曲柳(FraxinusmandschuricaRupr.)等树种的SLA明显增加。黎蕾等[25]的研究结果表明:随光照强度增强,催吐萝芙木(RauvolfiavomitoriaAfzel. ex Spreng.)的SLA减小;许中旗等[26]认为:在遮阳条件下蒙古栎(QuercusmongolicaFisch. ex Ledeb.)幼苗具有更高的SLA。在本研究中,透光率1%条件下三七单株叶面积显著小于其他处理组,而SLA则随透光率降低不断增大且差异显著,表明低光照环境下三七通过增加SLA提高叶片光能捕获能力, 进而提高叶片的光合生产能力和碳收获量。
叶绿素是衡量叶片光合作用性能的重要指标,单位叶面积叶绿素含量与叶片光合能力密切相关[27];叶绿素含量增加有利于增强光合作用能力、提高作物产量和品质;叶片叶绿素含量与SPAD值有极显著相关性[28]。吴飞燕等[29]的研究结果表明:随光照强度减弱,石栎〔Lithocarpusglaber(Thunb.) Nakai〕叶片的叶绿素含量增加。本研究中,在透光率相对较低的环境中三七幼苗叶片的SPAD值较大,说明在低光照条件下三七叶片的光合色素含量增加,有利于三七植株在低光照环境中捕获更多的光能进行光合作用,从而增加碳收获量和干物质的积累。
三七是一种典型的喜阴植物,适当遮阳有利于其生长和干物质积累。当环境中的光照水平低于其最适值时,三七植株的外部形态朝着有利于捕获更多光能的方向变化;其生长策略是将生物量优先分配到茎部和叶部,以捕获更多的光能克服光资源限制。同时,在低光照环境中三七通过增加比叶面积和叶绿素含量提高叶片光能捕获能力,进而提高光合生产能力和碳收获能力。总的来看,在低光照环境下三七幼苗的形态特征、生长特性和色素含量等变化都是以捕获更多光能、提高光合能力和碳收获量为优先策略,且在透光率8%条件下三七幼苗生长较佳。
参考文献:
[1] 国家药典委员会. 中华人民共和国药典: 2005年版(一部)[M]. 北京: 化学工业出版社, 2005: 10-11.
[2] 黄社霄. 人参、西洋参、三七药用探讨[J]. 中国中医药现代远程教育, 2011, 9(15): 73-74.
[3] 陈中坚. 以“三七”名称命名的药用植物概述[J]. 特产研究, 2009(2): 23-25.
[4] 郑光植, 杨崇仁. 三七生物学及其应用[M]. 北京: 科学出版社, 1994: 5.
[5] 杨红玉. 三七生长生理及栽培研究的现状与方向[J]. 中医药学报, 2000(4): 37-39.
[6] 陈中坚, 曾 江, 王 勇, 等. 三七种植业现状调查[J]. 中药材, 2002, 25(6): 387-389.
[7] DISSANAYAKE P, GEORGE D L, GUPTA M L. Effect of light, gibberellic acid and abscisic acid on germination of guayule (PartheniumargentatumGray) seed [J]. Industrial Crops and Products, 2012, 32(2): 111-117.
[8] POORTER H, NAGEL O W. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review[J]. Australian Journal of Plant Physiology, 2000, 27(6): 595-607.
[9] DELAGRANGE S, MONTPIED P, DREYER E, et al. Does shade improve light interception efficiency? A comparison among seedlings from shade-tolerant and -intolerant temperate deciduous tree species[J]. New Phytologist, 2006, 172(2): 293-304.
[10] FENG Y L, WANG J F, SANG W G. Biomass allocation, mor-phology and photosynthesis of invasive and noninvasive exotic species grown at four irradiance levels[J]. Acta Oecologica, 2007, 31(1): 40-47.
[11] De SIMONE S, OKA Y, INOUE Y. Effects of light on root hair formation inArabidopsisthalianaphytochrome-deficient mutants[J]. Journal of Plant Research, 2000, 113(1): 63-69.
[12] WAHL S, RYSEY P, EDWARDS P J. Phenotypic plasticity of grass root anatomy in response to light intensity and nutrient supply[J]. Annals of Botany, 2001, 88(6): 1071-1078.
[13] NAGEL K A, SCHURR U, WALTER A. Dynamics of root growth stimulation inNicotianatabacumin increasing light intensity [J]. Plant, Cell and Environment, 2006, 29(10): 1936-1945.
[14] 郭巧生. 药用植物栽培学[M]. 北京: 高等教育出版社, 2004: 199.
[15] 罗 群, 游春梅, 官会林. 环境因素对三七生长影响的分析[J]. 中国西部科技, 2010, 9(9): 7-8, 12.
[16] 陈中坚, 崔秀明, 王朝梁, 等. 文山优质三七基地生态区划和布局研究[J]. 人参研究, 1999(3): 29-31.
[17] 潘玉梅, 刘明超, 唐赛春, 等. 光强对三叶鬼针草生长特征的影响[J]. 广西植物, 2012, 32(1): 77-82.
[18] 徐 飞, 郭卫华, 徐伟红, 等. 不同光环境对麻栎和刺槐幼苗生长和光合特征的影响[J]. 生态学报, 2010, 30(12): 3098-3107.
[19] 华劲松, 戴红燕, 夏明忠. 不同光照强度对芸豆光合特性及产量性状的影响[J]. 西北农业学报, 2009, 18(2): 136-140.
[20] 孟金柳, 曾 波, 叶小齐, 等. 不同光照水平下叶损失对樟(Cinnamomumcamphora)生物量分配的影响[J]. 西南师范大学学报: 自然科学版, 2004, 29(3): 439-444.
[21] 张 林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展[J]. 植物生态学报, 2004, 28(6): 844-852.
[22] WRIGHT I J, WESTOBY M, REICH P B. Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span[J]. Journal of Ecology, 2002, 90(3): 534-543.
[23] 潘复静, 孙英杰, 张德楠, 等. 桂北岩溶区青冈栎-青檀群落植物叶片比叶面积研究[J]. 广西植物, 2013, 33(3): 295-299.
[24] 李明财, 朱教君, 孙一荣. 东北次生林主要树种比叶面积对光照强度的响应[J]. 生态学杂志, 2009, 28(8): 1437-1442.
[25] 黎 蕾, 蔡传涛, 刘贵周. 光强和施氮量对催吐萝芙木生长及生物量的影响[J]. 中国生态农业学报, 2010, 18(1): 57-61.
[26] 许中旗, 黄选瑞, 徐成立, 等. 光照条件对蒙古栎幼苗生长及形态特征的影响[J]. 生态学报, 2009, 29(3): 1121-1128.
[27] MURCHIE E H, HORTON P. Acclimation of photosynthesis to irradiance and spectral quality in British plant species: chlorophyll content, photosynthetic capacity and habitat preference[J]. Plant, Cell and Environment, 1997, 20(4): 438-448.
[28] 雷泽湘, 艾天成, 李方敏, 等. 草莓叶片叶绿素含量、含氮量与SPAD值间的关系[J]. 湖北农学院学报, 2001, 21(2): 138-140.
[29] 吴飞燕, 伊力塔, 李修鹏, 等. 不同光照强度对石栎幼苗叶绿素含量及叶绿素荧光参数的影响[J]. 东北农业大学学报, 2012, 43(4): 88-92.
