不同营养方式对小球藻FACHB 484生长的影响及其非自养生长机理研究
2014-03-29牛海亚马玉龙石勋祥牛
牛海亚马玉龙,石勋祥牛 芮
(1. 宁夏大学能源化工国家重点实验室培育基地, 宁夏 750021; 2. 宁夏大学化学化工学院, 宁夏 750021)
不同营养方式对小球藻FACHB 484生长的影响及其非自养生长机理研究
牛海亚1马玉龙1,2石勋祥1牛 芮1
(1. 宁夏大学能源化工国家重点实验室培育基地, 宁夏 750021; 2. 宁夏大学化学化工学院, 宁夏 750021)
系统研究了小球藻FACHB 484在含有葡萄糖的不同营养方式下的生长情况, 并通过抑制试验探讨葡萄糖在小球藻FACHB 484光异养和兼养生长条件下所起的作用以及小球藻FACHB 484是否存在氧化呼吸系统的关键酶类。结果表明: 小球藻FACHB 484可利用葡萄糖进行化能异养、光激活异养、光异养及兼养生长, 其生长速率大小为: 兼养>光异养>光激活异养>化能异养>光合自养。兼养培养的最大生物量和比生长速率分别是自养培养的8.6和3.4倍, 其比生长速率接近于光合自养和光异养培养下的比生长速率之和。葡萄糖主要作为小球藻FACHB 484兼养和光异养培养的碳源, 而能量主要源自光。小球藻FACHB 484存在氧化呼吸链代谢途径, 其细胞中有琥珀酸脱氢酶和细胞色素氧化酶。
小球藻; 异养生长; 兼养生长; 代谢机理
小球藻(Chlorella)为绿藻门、小球藻属的单细胞绿藻[1]。与高等植物相比, 小球藻不仅可利用光能和CO2进行光合自养(Photoautotrophy)生长, 而且可以利用有机碳进行异养(Heterotrophy)和兼养生长(Mixotrophy)[2]。光合自养的光抑制性是限制微藻生物量增加的主要原因之一。异养培养以外加有机碳为碳源和能源[3], 能够降低或消除光合自养培养的不足。异养培养和兼养培养不仅能提高微藻的生物量[4], 而且微藻的细胞结构和生化组分也发生了变化。缪晓玲等[5]通过异养转化细胞工程技术获得了油脂含量高达 57.9%的异养小球藻, 使油脂含量在化合自养的基础上提高了4倍。
目前, 关于微藻异养、兼养的生长机理还不清楚。一些微藻尽管有代谢有机碳所必须的酶系, 但对有机碳却不能同化吸收, 如小定鞭藻(Prymnesium parvum)和杜氏盐藻(Dunaliella tertiolecta)[6]。Gladue[7]对一些微藻没有异养生长能力的原因提出了三种假说: 通透性障碍、相关酶的缺乏和限制性的呼吸能力。但不同微藻或同一种微藻不同品系间的异养、兼养生长机理可能存在差异。吴庆余等[8]对小球藻(Chlorella protothecoides)的两种品系(中国品系C与美国品系A)在高浓度葡萄糖、低浓度有机氮的条件下进行异养培养, 发现两株小球藻在生长速率、叶绿素含量及细胞内腺苷酸能荷值(AEC)方面存在很大的差异性。近年来, 关于小球藻培养的研究报道主要集中在自养培养和异养培养方面[9], 但对小球藻的自养、化能异养 (Chemoheterotrophy)、光激活异养 (Light-activated heterotrophic growth, LAHG)、光异养 (Photoheterotrophy)和兼养培养的系统比较研究报道较少, 尤其在利用葡萄糖代谢机理方面的研究报道甚少。本研究针对以上两个方面进行试验, 为小球藻的基础研究及利用小球藻进行生产研究的其他领域提供相关数据。
1 材料与方法
1.1 藻种
小球藻FACHB 484(Chlorella sp. 484)购自中国科学院典型培养物保藏委员会淡水藻种库(FACHB)。
1.2 培养基与试剂
SE基础培养液(1L), 其配方如下: NaNO3250 mg, K2HPO475 mg, MgSO4·7H2O 75 mg, CaCl2·2H2O 25 mg, KH2PO4175 mg, NaCl 25 mg, FeCl3·6H2O 5 mg, EDTA-Fe 1 mL, Trace mental solution 1 mL,土壤提取液40 mL, 蒸馏水958 mL。
试剂: 葡萄糖, 3-(3, 4-二氯苯基)-1, 1-二甲脲(DCMU, 购于Sigma公司), 2, 4-二硝基苯酚, 叠氮化钠, 丙二酸钠, 头孢噻肟钠。
1.3 培养条件
将配置分装好的培养基在高压灭菌锅中 121℃灭菌 20min, 灭菌后冷却至室温, 添加头孢噻肟钠至0.5 mg/L防止染菌。在超净台中将生长至对数期的微藻接种于含150 mL培养基的250 mL锥形瓶中(接种量20, v/v), 温度30℃、平均光强5000 lx的培养箱中培养, 光暗比12h 12h∶ 。
1.4 试验设计
小球藻 FACHB 484不同营养方式比较 光合自养培养: 将微藻接种于SE基础培养基中, 在光照条件下培养。设置三个平行, 每隔 24h测定其生物量及pH变化。兼养培养: 在SE培养基中添加碳浓度200 mg/L的葡萄糖, 于光照条件下培养。设置三个平行, 每隔24h测定其生物量及pH变化。化能异养: 在SE培养基中添加碳浓度200 mg/L的葡萄糖, 将锥形瓶用黑色布袋包裹, 于完全黑暗条件下培养。设置三个平行, 每隔 24h测定其生物量及pH变化。光激活异养: 在SE培养基中添加碳浓度200 mg/L的葡萄糖, 将锥形瓶用黑色布袋包裹, 于完全黑暗条件下培养, 每24h取下布袋, 以5000 lx脉冲式短波照射 10min。设置三个平行, 每隔 24h测定其生物量及pH变化。光异养: 在SE培养基中添加碳浓度200 mg/L的葡萄糖, 并添加10–6mol/L 3-(3, 4-二氯苯基)-1, 1-二甲脲(DCMU), 于光照条件下培养。设置三个平行, 每隔 24h测定其生物量及pH变化。
兼养与光异养碳源试验 在自养、兼养、和光异养、培养基中分别添加5.0×10–7和5×10–5mmol/L的2, 4-二硝基苯酚(DNP), 设置三个平行, 每隔24h测定其生物量。
呼吸链抑制剂对小球藻FACHB 484生长的影响在自养和兼养培养基中分别添加0.1和1 mmol/L的丙二酸钠, 光照培养, 设置三个平行, 每隔24h测定其生物量。
在自养和兼养培养基中分别添加1和10 μmol/L的叠氮化钠, 光照培养, 设置三个平行, 每隔24h测定其生物量。
1.5 样品测定
藻细胞生物量测定。采用紫外可见分光光度仪测定微藻溶液在最大吸收波长λ690处的吸光度值(A690), 并采用血球计数板计数微藻细胞个数, 建立微藻细胞个数与吸光度值 A690之间的拟合曲线, 曲线方程如下:

式中y为微藻吸光度值A690, x为藻细胞个数。
微藻比生长速率(Specific growth rate): 微藻比生长速率是依据细胞密度值通过最小二乘法拟合得到, 用来评价微藻的日生长情况及平均比生长速率,即其在一定时期内的总体生长情况。公式如下[10]:
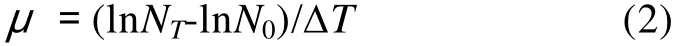
式中μ为微藻比生长速率(/d), N0为接种时微藻细胞密度, NT为培养T天后微藻细胞密度, ΔT为微藻接种到T天时的时间间隔。
2 结果
2.1 小球藻FACHB 484营养方式比较
由图1可知: 小球藻FACHB 484能利用葡萄糖进行化能异养、光激活异养、光异养和兼养生长。在化能异养和光激活异养条件下, 小球藻 FACHB 484颜色依然为绿色, 但生长缓慢。在添加了DCMU的光异养条件下小球藻 FACHB 484生长状况优于化能异养和光激活异养, 但在培养 12d以后因葡萄糖耗尽而死亡。小球藻FACHB 484在兼养条件下的生长状况最好。并且所试微藻兼养比生长速率(0.33)与光合自养比生长速率(0.13)和光异养比生长速率(0.19)之和(0.32)非常接近, 说明小球藻FACHB 484兼养培养比生长速率等于光异养培养与自养培养比生长速率之和。
2.2 在不同营养方式下小球藻FACHB 484体系pH变化

图1 小球藻FACHB 484在不同营养方式下的生长情况Fig. 1 The growth performance of Chlorella sp. FACHB 484 under different trophic conditions

图2 小球藻FACHB 484在不同营养方式下体系pH变化Fig. 2 The change of medium pH of Chlorella sp. 484 under different trophic conditions
由图2可知: 小球藻FACHB 484在加入葡萄糖进行化能异养、光激活异养、光异养和兼养生长时,体系pH都首先有一个小幅的下降, 之后又升高, 且升高的幅度与小球藻 FACHB 484生物量增加的幅度相一致。而在自养生长条件下, 小球藻 FACHB 484体系pH保持了一个小幅的波动上升趋势。推测小球藻FACHB 484在利用葡萄糖生长的初期, 代谢产生了一些小分子的酸类物质[11]。之后由于葡萄糖浓度下降, SE培养基中NaNO3为一种生物性碱,随着微藻细胞对3NO−的吸收, 培养基中 Na+过剩,微藻细胞泵出的OH–占优势, 使溶液pH不断升高,而在光合自养生长过程中微藻生长缓慢, 且由于藻液中昼夜CO2含量的不同, 使pH保持了一个小幅波动上升的趋势。小球藻FACHB 484能够利用葡萄糖进行化能异养、光激活异养、光异养和兼养生长, 说明小球藻FACHB 484中存在一些正常代谢的酶类。
2.3 兼养与光异养碳源试验
图3 2, 4-二硝基苯酚对不同培养方式下小球藻FACHB 484生长的影响Fig. 3 Effects of 2, 4-dinitrophenol on the growth of Chlorella sp. FACHB 484 under different conditions
上述研究表明: 小球藻FACHB 484能利用葡萄糖进行化能异养、光激活异养、光异养和兼养生长,为了进一步探明葡萄糖在小球藻 FACHB 484光异养和兼养生长中的作用, 本实验以DNP为解偶联剂,对葡萄糖光异养培养和兼养培养条件下的小球藻FACHB 484生长状况进行研究。从图3中可知, 在光异养培养和兼养培养条件下, 添加5×10–7和5× 10–5mmol/L DNP对小球藻FACHB 484生长未产生显著影响。
2.4 呼吸链抑制剂对小球藻FACHB 484生长的影响
大多数微藻为光合自养生物, 但许多微藻也能利用有机碳进行兼养和异养生长。只有少数微藻可以在完全黑暗的条件下利用有机碳进行异养生长。本试验已证明小球藻FACHB 484能够利用葡萄糖进行生长。为了进一步验证小球藻FACHB 484是否存在呼吸代谢途径和末端氧化系统的酶, 采用丙二酸钠和叠氮化钠对小球藻FACHB 484进行抑制试验。由图4、图5可知, 在自养培养和兼养培养条件下, 小球藻FACHB 484对丙二酸钠和叠氮化钠的抑制都敏感, 随抑制剂浓度的增加, 抑制效果明显增强。

图4 丙二酸钠对小球藻FACHB 484生长的影响Fig. 4 Effects of sodium malonate on the growth of Chlorella sp. FACHB 484
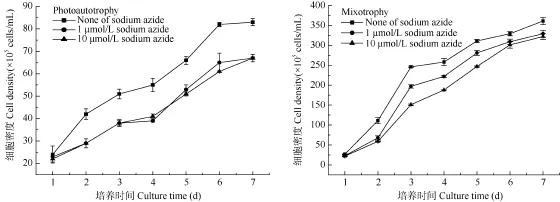
图5 叠氮化钠对小球藻FACHB 484生长的影响Fig. 5 Effects of sodium azide on the growth of Chlorella sp. FACHB 484
3 讨论
许多微藻具有利用有机碳进行兼养和异养生长的能力, 如鱼腥藻HB1017能利用果糖、葡萄糖和蔗糖为底物进行化能异养生长[12,13]。集胞藻 (Synechocystis sp.) PCC6803可以在每天一个短时光照(5min)条件下利用葡萄糖进行光激活异养[14,15], 三角褐指藻具有兼养生长的能力[16]。本试验对小球藻FACHB 484进行了自养、兼养和异养的系统试验。结果表明, 在含葡萄糖的培养基中, 小球藻 FACHB 484在不同营养方式下的生长情况为: 兼养生长>光异养生长>光激活异养生长>化能异养生长>光合自养(图1)。在光异养条件下培养小球藻FACHB 484, 12d以后死亡, 是由于DCMU是光合作用抑制剂, 在培养基中加入 DCMU以后,微藻细胞光系统Ⅱ的活性被抑制, 阻断了非环式电子传递, NADPH不再产生, CO2不能被微藻同化吸收[17—19]。所以当葡萄糖被耗尽时, 如不再添加碳源,微藻细胞由于缺少生长所必须的碳源而死亡。文献报道, 大多数微藻兼养的比生长速率等于异养培养与自养培养比生长速率之和(μ兼养=μ异养+μ自养), 兼养培养兼有光合自养与异养代谢的性质, 在光合自养条件下, CO2作为唯一碳源维持微藻生长, 在光异养条件下, 非环式电子传递被阻断, CO2不能被同化吸收,微藻只能利用有机碳进行生长。本试验研究结果表明, 小球藻FACHB 484兼养的比生长速率等于光合自养培养与光异养培养比生长速率之和, 兼养培养是获得小球藻FACHB 484高密度、高产量、短周期培养的最佳营养方式。
本实验以DNP为解偶联剂, 进一步探明葡萄糖在小球藻FACHB 484光异养和兼养生长中的作用。生物体的磷酸化有代谢连接的磷酸化和呼吸链连接的磷酸化两种类型, 即底物水平磷酸化和氧化磷酸化。氧化磷酸化的作用是有机物在分解过程中释放的能量用于ATP的合成。DNP是一种疏水性质子载体, 作为氧化磷酸化的解偶联剂, DNP能轻易扩散穿过线粒体内膜, 以质子化的形式将膜间隙的H+带回线粒体并释放到基质中, 从而消除了线粒体内膜两侧的H+浓度梯度, 破坏了激活ATP合成酶的质子驱动力[20], ATP不能被合成, 使氧化和磷酸化脱偶联, 氧化释放的能量全部以热的形式散发。DNP对小球藻FACHB 484生长未产生显著影响(图3), 因此可以排除小球藻 FACHB 484在光异养和兼养培养条件下主要通过葡萄糖氧化磷酸化提供能量的途径。这说明小球藻FACHB 484在光异养和兼养培养条件下的能量来源是光, 而葡萄糖主要为小球藻FACHB 484生长起提供碳源的作用。
为了进一步验证小球藻 FACHB 484是否存在呼吸代谢途径和末端氧化系统的酶, 采用丙二酸钠和叠氮化钠对小球藻FACHB 484进行抑制试验。琥珀酸脱氢酶是连接氧化磷酸化与电子传递的枢纽之一, 具有严格的立体专一性。丙二酸是琥珀酸的类似物, 是琥珀酸脱氢酶的强有力的竞争性抑制物,可以阻断三羧酸循环。细胞色素氧化酶是电子传递链末端的酶。细胞色素aa3和P450辅基中的铁原子可以与叠氮化合物形成一个配位键, 阻断呼吸链的电子传递。小球藻FACHB 484对丙二酸钠和叠氮化钠的抑制都敏感, 随抑制剂浓度的增加, 抑制效果明显增强(图4、图5)。这说明小球藻FACHB 484线粒体内膜存在琥珀酸脱氢酶和细胞色素氧化酶系统。因此可见, 小球藻FACHB 484中存在氧化呼吸系统的一些关键酶系。
[1] Li J S, Wang P, Dong L M. Chlorella high density training and lipid extraction condition optimizing [J]. Microbiology China, 2012, 39(4): 486—494 [李金穗, 汪苹, 董黎明. 小球藻高密度培养及油脂提取条件的优化. 微生物学通报, 2012, 39(4): 486—494]
[2] Liu X J. Characteristics of photoautotrophy, mixotrophy and heterotrophy of Phaeodactylum tricornutum [D]. Thesis for Doctor of Science. Jinan University, Guangzhou. 2008 [刘晓娟. 三角褐指藻的自养、兼养和异养特性研究. 博士学位论文, 暨南大学, 广州. 2008]
[3] Miao X L, Wu Q Y. Biodiesel production from heterotrophic microalgal oil [J]. Bioresource Technology, 2006, 97(6): 841—846
[4] Li C, Feng Y J, Zhao D W, et al. Microalgae lipid accumulation and carbon flux analysis based on municipal wastewater used as medium [J]. Renewable Energy Resources, 2012, 30(6): 93—96 [李超, 冯玉杰, 张大伟,等. 以市政污水为底物的微藻油脂积累和碳流分析. 可再生能源, 2012, 30(6): 93—96]
[5] Miao X L, Wu Q Y. Bio-oil fuel production from microalgae after heterotrophic growth [J]. Renewable Energy Resources, 2004, (4): 41—43 [缪晓玲, 吴庆余. 藻类异养转化制备生物油燃料技术. 可再生能源, 2004, (4): 41—43]
[6] Octavio P G, Froylan M.E. Escalante, Luz E. de-Bashan, et al. Heterotrophic cultures of microalgae: Metabolism and potential products [J]. Water Research, 2011, (45): 11—36
[7] Gladue R M. Heterotrophic microalgae prodution: potential for application to aquaculture feeds [C]. Fulks w. Main K L. Rotifer and Microalgae culture systems. Honolulu: Proceedings of a US-Asia Workshop. 1991: 275—286
[8] Wu Q Y, Kuang M. Growth AEC and pigments of Chlorella protothecoides in two strains under the autotrophic and heterophic conditions [J]. Plant Physiology Journal, 1992, 18(3): 293—299 [吴庆余, 匡梅. 小球藻两个品系在自养与异养条件下的生长、能荷与色素差异. 植物生理学报, 1992, 18(3): 293—299]
[9] Cao Y T, Kong W B, Xi Y Q, et al. The effects of different nutrition modes on growth characteristics and cell components of Chlorella vulgaris [J]. Food and Fermentation Industries, 2011, 37(10): 45—50 [曹云涛, 孔维宝, 葸玉琴, 等. 不同营养方式对普通小球藻生长特性和细胞组成的影响. 食品与发酵工业, 2011, 37(10): 45—50]
[10] Hu Z X, Xu N, Duan Y S. Effects of nitrogen sources on the growth of Heterosigma akashiw, Karenia sp., Phaeocystis globosa and Chaetoceros sp. [J]. Ecology and Environmental Sciences, 2010, 19(10): 2452—2457 [胡章喜, 徐宁, 段舜山. 不同氮源对4种海洋微藻生长的影响. 生态环境学报, 2010, 19(10): 2452—2457]
[11] Yu G C, Xin X F, Cai Z L, et al. Mixotrophic cultures of Anabaena sp. PCC7120 [J]. Engineering Chemistry& Metallurgy, 2000, 21(1): 52—57 [喻国策, 辛晓峰, 蔡昭铃,等. 鱼腥藻Anabaena sp. PCC7120的混合营养生长. 化工冶金, 2000, 21(1): 52—57]
[12] Jin C Y, Song L R, Li S H. Study on chemoheterotrophic growth of Anabaena HB1017 [J]. Acta Hydrobiologica Sinica, 1995, 19(1): 13—19 [金传荫, 宋立荣, 黎尚豪. 鱼腥藻HB1017株化能异养研究. 水生生物学报, 1995, 19(1): 13—19]
[13] Jin C Y, Song L R, Li S H. The mixotrophic growth of Anabaena sp. HB1017 [J]. Acta Hydrobiologica Sinica, 1996, 20(2): 134—137 [金传荫, 宋立荣, 黎尚豪. 鱼腥藻 1017株的混合混合营养型生长. 水生生物学报, 1996, 20(2): 134—137]
[14] Anderson S L, Mclntosh L. Light-activated heterotrophic growth of the cyanobacterium Synechocystis sp. strain PCC 6803: a blue-light-requiring process [J]. Journal of Bacteriology, 1991, (173): 2761—2767
[15] Liu Z Y, Xu X D, Kong R Q. Studies on s110886, a gene essential to light-activated heterotrophic growth in Synechocystis sp. PCC6803 [J]. Acta Hydrobiologica Sinica, 2006, 30(4): 375—379 [刘朝莹, 徐旭东, 孔任秋. 集胞藻PCC6803光激活异养生长必需基因s110886的研究. 水生生物学报, 2006, 30(4): 375—379]
[16] Liu X J, Duan Y S, Li A F. Effects of organic carbon source on growth, biochemical components and fatty acid composition of Phaeodactylum tricornutum [J]. Chinese Journal of Biotechnology, 2008, 24(1): 147—152 [刘晓娟, 段舜山, 李爱芬. 有机碳源对三角褐指藻生长胞内物质和脂肪酸组分的影响. 生物工程学报, 2008, 24(1): 147—152]
[17] Chen L Z, Xie M, Bi Y H, et al. The combined effects of UV-B radiation and herbicides on photosynthesis, antioxidant enzymes and DNA damage in two bloom-forming cyanobacteria [J]. Ecotoxicology and Environmental Safety, 2012, (80): 224—230
[18] Xia J, Li Y J, Zou D. Effects of salinity stress on PSII in Ulva lactuca as probed by chlorophyll fuorescence measurements [J]. Aquatic Botany, 2004, (80): 129—137
[19] Yu L H, Guo H L, Xu X D, et al. Effect of the photosynthesis inhibitor DCMU on chlorophylls synthesis in heterotrophic cyanobacteria [J]. Acta Hydrobiologica Sinica, 2002, 26(1): 102—104 [余利红, 郭厚良, 徐旭东, 等. 光合抑制剂DCMU对异养生长蓝藻叶绿素合成的作用. 水生生物学报, 2002, 26(1): 102—104]
[20] Lin Q S, Wu Q R. Studies on oxidative phosphorylation Ⅱ. The release of inhibition of 2, 4-dinitrophenol on succinate oxidation in rat-liver mitochondria [J]. Acta Biochimica et Biophysica Sinica, 1963, 3(3): 293—300 [林其谁, 伍钦荣.氧化磷酸化作用的研究Ⅱ2, 4-二硝基苯酚对琥珀酸氧化抑制效应的解除. 生物化学与生物物理学报, 1963, 3(3): 293—300]
STUDY ON THE EFFECTS OF DIFFERENT NUTRITION MODES ON THE GROWTH OF CHLORELLA SP. FACHB 484 AND THE MECHANISM OF HETEROTROPHIC GROWTH
NIU Hai-Ya1, MA Yu-Long1,2, SHI Xun-Xiang1and NIU Rui1
(1. State Key Laboratory Cultivation Base of Energy Resources & Chemical Engineering, Ningxia University, Yinchuan 750021, China; 2. College of Chemistry and Chemical Engineering, Ningxia University, Yinchuan 750021, China)
Here we systematically studied the growth of Chlorella sp. FACHB 484 in glucose-containing medium in different nutrition modes. We also conducted the inhibition tests to investigate the effects of glucose on the growth of Chlorella sp. FACHB 484 in heterotrophic and mixotrophic conditions, as well as to identify the key enzyme in oxidation respiratory system. We observed that Chlorella sp. FACHB 484 could utilize glucose as the source of nutrition in all the test nutritional modes and showed different growth rates in the order mixotrophy > photoheterotrophy > light-activated heterotrophy > chemoheterotrophy > photoautotrophy. The maximum biomass and specific growth rate (SGR) of the tested algae in mixotrophy were 8.6 and 3.4 times higher than those in photoautotrophy. The value of SGR of Chlorella sp. FACHB 484 in mixotrophy was close to the sum of the values of SGRs in photoautotrophy and photoheterotrophy. In mixotrophic and photoheterotrophic modes, Chlorella sp. FACHB 484 used glucose and light as the sources of carbon and energy respectively. The cells of Chlorella sp. FACHB 484 expresses succinic dehydrogenase and cytochrome oxidase which are most likely the key enzymes of the oxidation respiratory chain.
Chlorella sp.; Heterotrophic growth; Mixotrophic growth; Metabolic mechanism
S968; Q949.2
A
1000-3207(2014)03-0474-06
10.7541/2014.67
2013-01-28;
2013-10-17
国家973计划前期研究专项(No. 2012CB723106); 宁夏环境科学研究项目(No. 20100803)资助
牛海亚(1987—), 女, 宁夏固原人; 硕士研究生; 主要研究方向为生物质能源。E-mail: bixinyuhu@qq.com
马玉龙(1966—), 男, 回族, 博士后, 教授; 主要研究方向为生物质冶炼与高值转化。E-mail: nxylma@163.com
