鄱阳湖沿岸带螺类的物种组成及其生境
2014-03-29严家跃黄心一陈家宽
严家跃黄心一陈家宽,
(1. 复旦大学生物多样性与生态工程教育部重点实验室, 上海 200433; 2. 南昌大学生命科学研究院流域生态学研究所 南昌330031)
鄱阳湖沿岸带螺类的物种组成及其生境
严家跃1黄心一1陈家宽1,2
(1. 复旦大学生物多样性与生态工程教育部重点实验室, 上海 200433; 2. 南昌大学生命科学研究院流域生态学研究所 南昌330031)
2011年5月和6月在鄱阳湖沿岸带的62个样点采集了淡水螺类样本并调查了生境类型, 分析了淡水螺类的物种组成及其生境分布特征。结果表明: 共采集到淡水螺类19种, 分属田螺科(8种)、 觿螺科(8种)、椎实螺科(2种)和黑螺科(1种)。与前人的研究相比, 本次调查的螺类物种数有较大的下降, 但田螺科和 觿螺科的物种仍是整个螺类群落的主要组成部分。淤泥生境的螺类物种组成与其他生境的差异较大, 淤泥生境的螺类物种丰富度显著高于沙土、砂石和水草3种生境 (P<0.05) , 表明螺类对生境具有一定的栖息偏好性。最后详细讨论了淡水螺类种数变化、物种组成及其生境分布的影响因素。
鄱阳湖; 淡水螺类; 物种多样性; 生境类型
淡水螺类广泛地分布于陆地水域, 并在水生植物、底泥、砂石和腐木等不同生境中高度特化[1]。在水生生态系统中, 它们既可作为分解者分解动植物残体和有机碎屑[2], 又能作为消费者牧食高等藻类和维管植物[3—5], 还能作为次级生产者被鱼类、甲壳类和涉禽类等多种动物取食[6,7], 因而在物质循环和能量流动中发挥着重要的作用。然而, 淡水螺类却因其较小的个体和不起眼的外形长期被人们所忽视, 并在日益增强的经济开发活动中遭受着前所未有的威胁[1], 如鄱阳湖螺类正遭受湖泊养殖、采砂和污染等方面的损害。
另一方面, 水文特征和水生植被等自然条件的改变会影响淡水螺类的生长和扩散, 并最终影响整个群落的组成和分布[8]。因此, 调查螺类群落的物种组成和分布是其生态学研究的基础。目前, 国内有关淡水螺类的研究多包含在大型底栖动物研究当中[9—12], 专门针对淡水螺类群落的研究还比较少[13—16]。在这些研究当中, 仅有陈晔光[13]和吴和利[16]对鄱阳湖全境的螺类群落进行过调查。前者仅对调查所得种类及其生境进行了描述, 后者对淡水螺类的生境也仅作了描述。可见, 不同生境类型下的螺类物种组成及其分布特征尚缺乏研究。
淡水螺类具有扩散能力弱、易采集和高敏感度等特点, 可以作为环境变化的指示生物[17]。浅水沿岸带频繁的水陆交替会引起生境条件的迅速改变,对淡水螺类的组成和分布产生影响。因此, 研究螺类物种组成的变化有助于了解生境的变化过程与影响。本文作者在2011年5月和6月对鄱阳湖沿岸带4种不同生境的螺类群落进行了调查, 并初步研究了生境类型与螺类物种组成的关系。
1 材料与方法
1.1 研究区概况
鄱阳湖是中国第一大淡水湖, 位于长江中游南岸, 江西省北部, 介于115°49′—116°46′N, 28°24′—29°46′E。鄱阳湖湖面水体是 “上游五河” 来水与尾闾长江退水水量吞吐的动态平衡结果[18], 至今仍保持通江、浅水和丰草的湖泊自然特征。鄱阳湖水位的季节变化巨大, 夏秋季节, 湖口水位达到 22 m (吴淞基面) 时, 鄱阳湖水体的面积为 40.78×102km2,容积为 300.89×108m3; 冬春季节, 湖口水位下降到12 m时, 水面仅为5×102km2, 容积仅为9×108m3[19],呈现出 “洪水一片, 枯水一线”的自然景观。当水位较高时, 鄱阳湖的连片水域形成一个完整而典型的水生生态系统, 而到了枯水期, 湖面急剧萎缩, 大片洲滩显露, 形成独特的水陆交替的湖滩草洲生态系统[20]。该系统中的浅水沿岸带以其复杂的基质为淡水螺类提供了多样化的生境, 不同种螺类在淤泥、沙土、砂石及水草等生境中特化分布。
1.2 样品采集及标本鉴定
于2011年5月和6月进行采样。在鄱阳湖沿岸带选取了南矶山、赣江河口、修河河口、赣修交汇、星子县、庐山区、湖口县、都昌县西、都昌县东、鄱阳县和进贤县共 11块样地(图1), 每块样地随机设置4—6个样点, 每个样点依据螺类着生基质特征分为淤泥、沙土、砂石或水草生境 (图2) 。根据《 全国淡水生物物种资源调查技术规定 (试行) 》中的底栖动物物种资源调查方法[21], 结合实地情况确定了一个螺类半定量采集方案。即采用直径为30 cm、网眼为40目的抄网, 在每个样点随机采集3次样品,每次由相同人员采集 5min。将采集到的 3次泥水混合物样品用 40目的筛网现场淘洗, 将 3次筛洗后获取的淡水螺类标本合并为一个样本, 立即用75%的乙醇溶液保存, 带回实验室后依据《中国经济动物志·淡水软体动物》[22]的系统对标本进行分类鉴定。
1.3 数据处理及统计
螺类的物种组成以科属种分类单元描述, 物种在各样地中的分布特征以出现频率(Occurrence frequency)描述, 计算公式为:
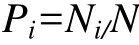
式中: Pi为第i个物种在所有调查样点中的出现频率; Ni为第i个物种在所有调查生境的出现次数; N为调查样点总数。生境间的物种组成差异以β多样性中的Cody指数(βC)描述, 计算公式为:
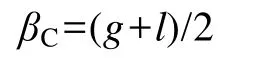
式中: g为生境A有而生境B 无的物种数; l为生境A 无而生境B 有的物种数。
螺类的生境分布特征主要以物种丰富度(S)的差异描述: 即对 4种生境和相应的螺类物种丰富度进行单因素方差分析(ANOVA), 然后用LSD法进行多重比较。数据处理均采用 Excel和 IBM SPSS Statistics 19软件进行。
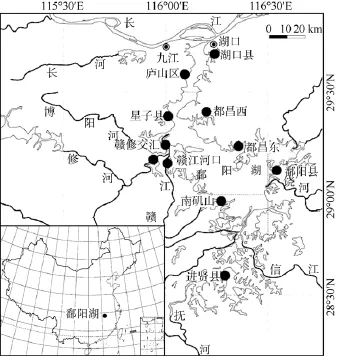
图1 样地分布图Fig. 1 Distribution of plots

图2 四种典型采样生境Fig. 2 Four representative sampling habitat types
2 结果
2.1 物种组成和分布
初步确定鄱阳湖沿岸带的螺类有4科9属19种(图3)。螺类的种类组成和地理分布(表1)显示, 各科物种数相差较大, 主要以田螺科(8种) 觿和 螺科(8种)的种类占优, 共占总物种数的 84%; 区系成分以华中区、华南区和华北区的种类优势明显, 各占总种数的80%、73.3%和60%; 不同种螺类在62个样点的出现频率差异较大, 其中以铜锈环棱螺、梨形环棱螺、大沼螺及方格短沟蜷出现频率最高, 各为96.77%、54.84%、46.77%和30.65%。
2.2 生境之间的物种组成差异
β多样性是反映生境间物种组成差异的重要指标, 生境间的共有种越少, β多样性的数值越大。Cody多样性指数(βc)是测度β多样性的一个重要方法[23]。在4种生境间的βc结果矩阵(表2)显示, 多样性指数高于5.0的占总数的一半, 低于2.5的也占一半, 2.5—5.0的区间出现断层; 高于5.0的多样性指数均为生境2、3、4分别与生境1比较的结果, 低于2.5的多样性指数则是生境2、3、4之间的比较结果。这说明生境1与生境2、3、4的物种组成差异较大, 但生境2、3、4之间的物种组成差异较小。

图3 鄱阳湖的19种淡水螺类Fig. 3 19 freshwater snail species of Poyang Lake
2.3 生境类型与物种丰富度的关系
淡水螺类在鄱阳湖沿岸带的多种栖息生境中均有分布, 但对淤泥、沙土、砂石和水草生境具有不同的栖息偏好性。淡水螺类在4种生境中的物种丰富度平均值分别为4.31、2.07、2.63和2.57, 单因素方差分析和多重比较结果表明, 淤泥生境中的螺类物种丰富度分别显著高于沙土、砂石和水草3种生境 (P<0.05) , 且沙土、砂石和水草3种生境中的螺类物种丰富度并无显著差异(P>0.05) (图4) 。这表明螺类偏好分布于淤泥生境中, 而对沙土、砂石和水草3种生境没有偏好性。
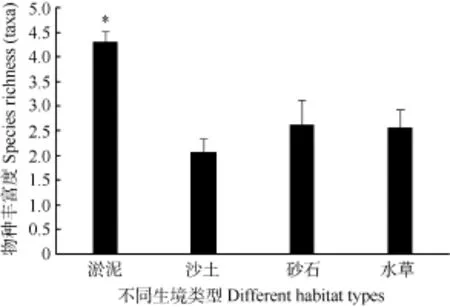
图4 不同生境类型对螺类物种丰富度的影响Fig. 4 The effect of different habitat types on snail’s species richness
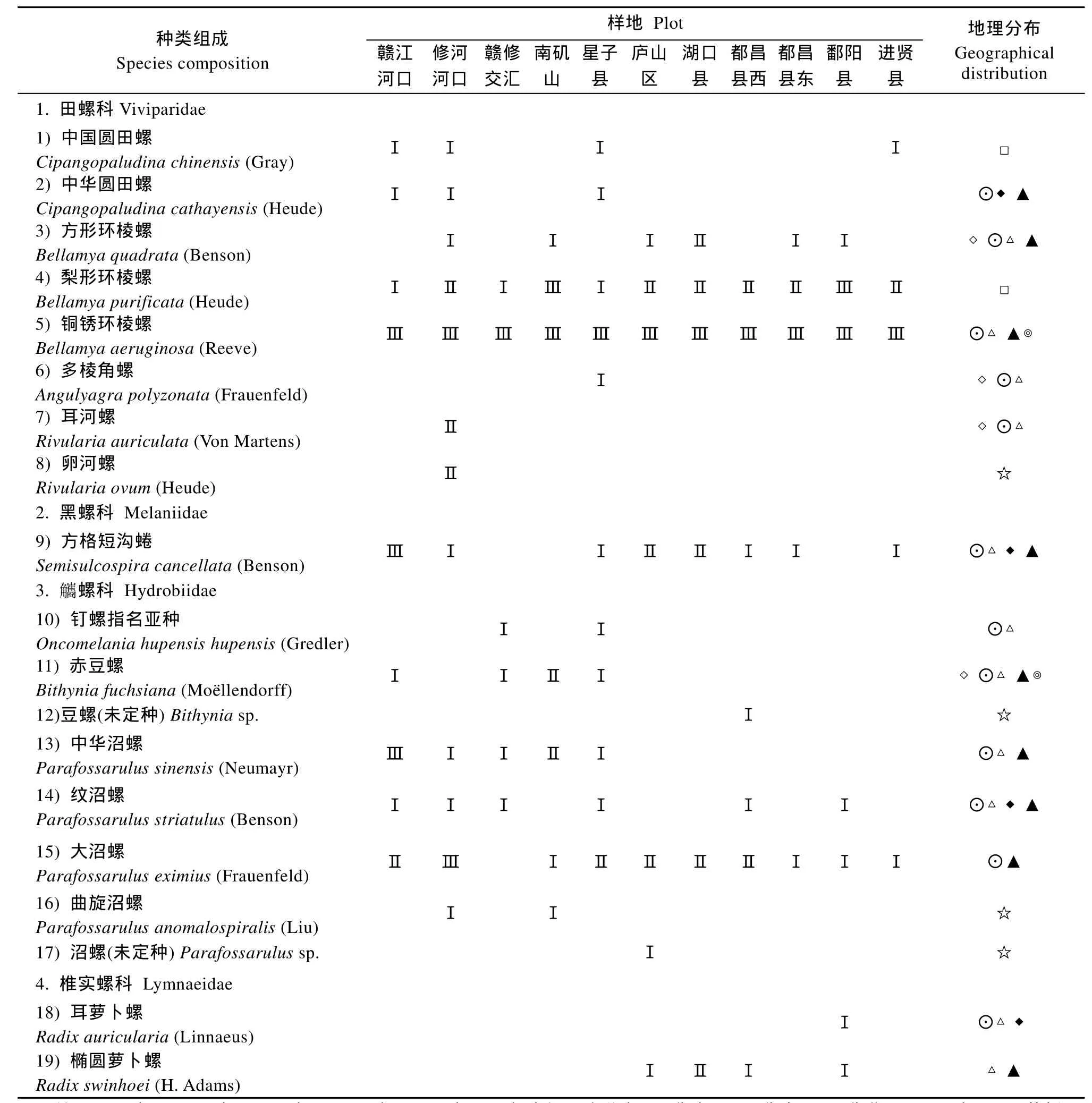
表1 鄱阳湖沿岸带螺类的种类组成和地理分布Tab. 1 Species composition and geographical distribution of freshwater snail in the littoral zone of Poyang Lake
3 讨论
3.1 物种组成变化的原因
与陈晔光[13]和吴和利[16]的调查结果相比, 本研究的物种组成有较大的变化: 前者共发现41种, 后者共发现26种, 本次调查仅发现19种螺类, 其中包含了2种未定种; 本调查发现的螺类中仅有13种为前两次研究均报道过的, 没有发现膀胱螺科和扁蜷螺科的种类, 田螺科的物种也较少; 本调查新增了多棱角螺Angulyagra polyzonata (Frauenfeld)、曲旋阵沼螺 Parafossarulus anomalospiralis (Liu)和卵河螺Rivularia ovum (Heude) 3种螺类。

表2 4种生境间的β多样性Cody指数测度结果矩Tab. 2 Matrix of β-diversity measured by cody index from four different habitats
物种组成的变化可能是采样的随机性误差和生境的确定性破坏共同影响的结果。首先采样的强度和范围不同。陈晔光[13]的数据来自 1963年、1984年和1985年的数次调查, 调查范围包括了鄱阳湖沿岸带和周围水域; 吴和利[16]则在枯水期和丰水期分别对鄱阳湖18个断面展开调查; 而本研究在枯水期对鄱阳湖沿岸带4种生境的62个样点进行了调查。其次采样的方法不同。陈晔光[13]的采样方法不详,吴和利[16]主要采用了定量采样方法, 而本研究则采用了半定量采样方法。再次, 物种鉴定可能存在一定的误差。现行的螺类鉴定方法大多是基于刘月英等[22]经典体系之上的形态学分类, 且 3次采样的最大时间跨度长达50年, 不排除部分螺类的形态学特征已发生变异, 造成鉴定误差。 最后, 鄱阳湖近年来的人为干扰日趋严重, 养殖、捕捞、采砂等经济活动或直接通过食物链影响螺类[24], 或通过破坏螺类的自然生境而威胁螺类[16], 这些确定性的变化可能会引起物种数目的下降。
3.2 物种组成的影响机制
在本次调查得到的19种螺类中, 田螺科和 觿螺科各占 8种, 为什么这两科的种类能在螺类群落中占有如此高的物种丰富度? Taylor, et al.[25]提出的种库假说(Species pool hypothesis)认为群落的物种多样性是区域过程与局域过程共同作用的结果。不少研究已表明局域过程(竞争、捕食等)是控制螺类多样性的主导过程[26,27], 资源竞争会限制螺类的生长和繁殖, 可能会造成物种之间的竞争排斥; 而捕食者对不同体型螺类的捕食偏好也能影响螺类的多度[26,28],最终可能改变螺类群落的物种组成。
有趣的是, 田螺科和 觿螺科的体型特征恰恰使其在资源竞争和逃避捕食当中显露优势。鄱阳湖4个科的体型从小到大依次是 觿螺科、椎实螺科、黑螺科和田螺科。在资源竞争方面,大体型的田螺科运动能力最强, 优先占有资源, 而小体型的 觿螺科能利用更多微小生境中的资源; 在逃避捕食方面, 鱼类和小龙虾均难以对大体型的田螺科进行碾压, 而小体型的 觿螺科更容易躲过捕食者的搜寻, 降低被捕食的概率。竞争和捕食等生态过程可能是影响鄱阳湖螺类物种组成的关键因素。
3.3 生境分布的影响机制
鄱阳湖螺类群落在沿岸带的不同生境中具有显著的分布格局: 即相比于沙土、砂石和水草 3种生境, 淡水螺类偏好分布于淤泥生境中, 淤泥生境的物种组成与其他生境的差异也最大。淤泥生境中的有机碎屑含量高于沙土和砂石生境, 因此淡水螺类可能优先选择在淤泥生境中觅食和栖息。然而淡水螺类在水草生境中的物种丰富度却显著低于淤泥生境, 与另外两种生境相比也无显著优势, 这与前人的研究结果并不一致[9,29]。
Carpenter, et al.[30]和Thomas[31]提出的假说阐明水生植物与螺类具有互利合作关系: 即螺类牧食为水草移除附生藻类的覆盖, 降低了水草的光限制及附生藻类的营养盐竞争; 反过来, 水草为栖居的螺类提供牧食、产卵以及逃避捕食的基质。苦草Vallisneria等水生植物一般仅在浅水沿岸带生长,不适应水深较大的湖盆。2011年鄱阳湖流域发生了罕见的特大干旱, 沿岸带的迅速干涸导致原有的水生植物群落迅速被湿生植物群落所取代, 尽管深水湖盆逐渐退水成为浅水带, 但往往因种子库的缺乏而不能在当年发育形成水生植物群落(实地考察);另一方面低水位胁迫和强光照条件可能增强了水草的次生代谢, 降低了水草对螺类的适口性[3], 这样就会使螺类对水草的空间和资源可利用性均降低,最终诱导螺类进行生境转换, 即其他生境的偏好种迁入到淤泥生境中, 改变了淤泥生境的物种组成,形成淤泥生境中的组成复杂度和物种丰富度均高于水草生境的局面。
4 结论
通过本文研究及与前人研究的对比可以得出以下结论: (1) 初步确定鄱阳湖沿岸带的螺类有4科9属19种, 这与前人的研究结果有较大的出入。建议应尽快建立一套研究技术规范: 恪守螺类调查规范,补充除形态学方法以外的新鉴定技术, 以获得更可靠的鉴定结果。3次调查结果存在的随机误差不能掩盖螺类物种数的明显下降趋势, 应在保护淡水螺类物种多样性的基础上, 合理开发鄱阳湖自然资源。(2) 本次调查显示田螺科和 觿螺科的种类占据了鄱阳湖群落总种数的 84%, 前人的研究结果也显示了相近的物种组成比例。田螺科和 觿螺科在资源竞争和躲避捕食等方面的潜在优势可能解释了这种物种组成格局。(3) β多样性和单因素方差分析的结果分别显示淤泥生境中具有更复杂的物种组成和更高的物种丰富度, 表明螺类可能对淤泥生境具有栖息选择性。2011年干旱引起的螺类生境转换可能解释了这种特殊的生境分布格局。
致谢:
螺类调查与鉴定得到南昌大学吴小平教授和金斌松副教授的指导以及张笑辰、曾泽国、曾泰等同学的帮助, 鄱阳湖国家级自然保护区和南矶山湿地国家级自然保护区为采样提供了便利, 在此一并表示感谢!
[1] Strong E E, Gargominy O, Ponder W F, et al. Global diversity of gastropods (Gastropoda; Mollusca) in freshwater [J]. Hydrobiologia, 2008, 595(1): 149—166
[2] Brönmark C. How do herbivorous freshwater snails affect macrophyte: A comment [J]. Ecology, 1990, 71(3), 1212—1215
[3] Li Y K. Submerged Plants and Herbivious Snails [D]. Thesis for Doctor of Science. Wuhan University, Wuhan. 2004 [李永科. 沉水植物与牧食性螺类的关系研究. 博士学位论文,武汉大学, 武汉. 2004]
[4] Elger A, Lemoine D. Determinants of macrophyte palatability to the pond snail Lymnaea stagnalis [J]. Freshwater Biology, 2005, 50(1): 86—95
[5] Li K Y, Liu Z W, Hu Y H, et al. Snail Radix swinhoei (H. Adams) herbivory on three submerged plants [J]. Acta Ecologica Sinica, 2006, 26(10): 3221—3224 [李宽意, 刘正文, 胡耀辉, 等. 椭圆萝卜螺 Radix swinhoei(H.Adams)对三种沉水植物的牧食选择. 生态学报, 2006, 26(10): 3221—3224]
[6] Gong Z J, Xie P, Yan Y J. Theories and methods of studies on the secondary production of zoobenthos [J]. Journal of Lake Sciences, 2001, 13(1): 79—88 [龚志平, 谢平, 闫云君.底栖动物次级生产力研究的理论与方法. 湖泊科学, 2001, 13(1): 79—88]
[7] Bao Y X, Ge B M, Zheng X, et al. Spatial distribution and seasonal variation of the macrobenthic community on tidal flats of Tianhe, Wenzhou Bay [J]. Acta Zoologica Sinica, 2006, 52(1): 45—52 [鲍毅新, 葛宝明, 郑祥, 等. 温州湾天河滩涂大型底栖动物群落分布与季节变化. 动物学报, 2006, 52(1): 45—52]
[8] Beche L A, McElravy E P, Resh V H. Long-term seasonal variation in the biological traits of benthic-macroinvertebrates in two Mediterranean-climate streams in California, USA [J]. Freshwater Biology, 2006, 51(1): 56—75
[9] Yan Y J, Li X Y, Liang Y L. A comparative study on community structure of macrozoobenthos between macrophtic and algal lakes [J]. Journal of Lake Sciences, 2005, 17(2):176—182 [闫云君, 李晓宇, 梁彦龄. 草型湖泊和藻型湖泊中大型底栖动物群落结构的比较. 湖泊科学, 2005, 17(2): 176—182]
[10] Xie Z C, Ma K, Ye L, et al. Structure and spacial distributional pattern of macrozoobenthos in Bao’an Lake [J]. Acta Hydrobiologia Sinica, 2007, 31(2): 174—183 [谢志才,马凯, 叶麟, 等. 保安湖大型底栖动物结构与分布格局研究. 水生生物学报, 2007, 31(2): 174—183]
[11] Wang Q, Wang H J, Cui Y D. Community characteristics of the macrozoobenthos and bioassessment of water quality in Lake Donghu District, Wuhan [J]. Acta Hydrobiologia Sinica, 2010, 34(4): 739—746 [王琴, 王海军, 崔永德. 武汉东湖水网区底栖动物群落特征及其水质的生物学评价.水生生物学报, 2010, 34(4): 739—746]
[12] Pan B Z, Wang Z Y, He X B. Studies on assemblage characteristics of macrozoobenthos in the West River [J]. Acta Hydrobiologia Sinica, 2011, 35(5): 851—856 [潘保柱,王兆印, 何雪宝. 西江大型底栖动物群落特征研究. 水生生物学报, 2011, 35(5): 851—856]
[13] Chen Y G. The freshwater snails of Poyang Lake and its surrounding waters, Jiangxi province, China [J]. Sinozoologia, 1988, (6): 69—75
[陈晔光. 鄱阳湖及其周围水域的淡水螺类. 动物学集刊, 1988, (6): 69—75]
[14] Xue J Z. The ecology of benthionic spiral shells in small reservoir basins of west Zhejiang [J]. Journal of Hangzhou Teachers College (Natural Science Edition), 2002, 1(2): 44—46 [薛俊增. 浙西山区水库小流域枯水期底栖螺类生态学初步研究. 杭州师范学院学报(自然科学版), 2002, 1(2): 44—46]
[15] Liu J, Hu Z Q. The species composition and faunal analysis of freshwater snails in the middle reaches of Xiang River [J]. Life Science Research, 2006, 10(3): 248—251
[刘俊, 胡自强. 湘江中游江段螺类的种类组成及区系分析. 生命科学研究, 2006, 10(3): 248—251]
[16] Wu H L. Studies on the Community Structure and Biodiversity of Fresh Water Snails in Poyang Lake [D]. Thesis for Master of Science. Nanchang University, Nanchang. 2007 [吴和利. 鄱阳湖淡水螺类群落结构及生物多样性研究. 南昌, 南昌大学, 2007]
[17] Shao M L, Xie Z C, Ye L, et al. Monthly change of community structure of zoobenthos in Xiangxi Bay after impoundment of Three Gorges Reservoir [J], Acta Hydrobiologia Sinica, 2006, 30(1): 64—69 [邵美玲, 谢志才,叶麟, 等. 三峡水库蓄水后香溪河库湾底栖动物群落结构的变化. 水生生物学报, 2006, 30(1): 64—69]
[18] Zhang B. Studies on Poyang Lake [M]. Shanghai: Shanghai Science and Technology Press. 1988, 18 [张本. 鄱阳湖研究.上海: 上海科学技术出版社. 1988, 18]
[19] Xiong X Q, Yang R Q. Drainage of Jiangxi Province [M]. Wuhan: Changjiang Press. 2007, 196 [熊小群, 杨荣清.江西水系. 武汉: 长江出版社. 2007, 196]
[20] Jin B S, Nie M, Li Q, et al. Basic characteristics, challenges and key scientific questions of the Poyang Lake basin [J]. Resources and Environment in the Yangtze Basin, 2012, 21(3): 268—275 [金斌松, 聂明, 李琴, 等. 鄱阳湖流域基本特征、面临挑战和关键科学问题. 长江流域资源与环境, 2012, 21(3): 268—275]
[21] Natural Ecosystem Protection Office of China Environmental Protection Bureau. China Technical Regulations for Aquatic Species Resources Survey (Trial Implementation). Beijing: Ministry of Environmental Protection of People’s Republic of China. 2010 [国家环境保护局自然生态保护司.全国淡水生物物种资源调查技术规定(试行). 北京: 国家环保部. 2010]
[22] Liu Y Y, Zhang W Z, Wang Y X, et al. Chinese Economic Zoography: Freshwater Molluscs [M]. Beijing: Science Press. 1979, 6—64 [刘月英, 张文珍, 王跃先, 等. 中国经济动物志: 淡水软体动物. 北京: 科学出版社. 1979, 6—64]
[23] Ma K P, Liu C R, Liu Y M. Measurement of biotic community diversity , Ⅱ. Measurement of β diversity [J]. Biodiversity Science, 1995, 3(1): 38—43 [马克平, 刘灿然,刘玉明. 生物群落多样性的测度方法, Ⅱ. β多样性的测度方法. 生物多样性, 1995, 3(1): 38—43]
[24] Xu Q Q, Wang H Z, Zhang S P. The impact of overstocking of mitten crab, Eriocheir sinensis, on lacustrine zoobenthic community [J]. Acta Hydrobiologia Sinica, 2003, 27(1): 42—46 [许巧情, 王洪铸, 张世萍. 河蟹过度放养对湖泊底栖动物群落的影响. 水生生物学报, 2003, 27(1): 42—46]
[25] Taylor D R, Aarssen L W, Loehle C. On the relationship between r/K selection and environmental carrying capacity: a new habitat templet for plant life history strategies [J]. Oikos, 1990, 58(2): 239—250
[26] Nyström P, Péres J R. Crayfish predation on the common pond snail (Lymnaea stagnalis): the effect of habitat complexity and snail size on foraging efficiency [J]. Hydrobiologia, 1998, 368(1): 201—208
[27] Hoverman J T, Davis C J, Werner E E, et al. Environmental gradients and the structure of freshwater snail communities [J]. Ecography, 2011, 34(6): 1049-1058
[28] Osenberg C W, Mittelbach G G. Effects of body size on the predator-prey interaction between pumpkinseed sunfish and gastropods [J]. Ecological Monographs, 1989, 59(4): 405—432
[29] Brönmark C. Freshwater snail diversity: effects of pond area, habitat heterogeneity and isolation [J]. Oecologia, 1985, 67(1): 127—131
[30] Carpenter S R, Lodge D M. Effects of submersed macrophytes on ecosystem processes [J]. Aquatic Botany, 1986, 26(3—4): 341—370
[31] Thomas J D. Mutualistic interactions in freshwater modular systems with molluscan components [J]. Advances in Ecological Research, 1990, 20(1): 125—178
STUDY ON THE SPECIES COMPOSITION AND HABITATS OF FRESHWATER SNAIL IN THE LITTORAL ZONE OF POYANG LAKE
YAN Jia-Yue1, HUANG Xin-Yi1and CHEN Jia-Kuan1,2
(1. Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, Fudan University, Shanghai 200433, China; 2. Center for Watershed Ecology, Institute of Life Science, Nanchang University, Nanchang 330031, China)
To study the species diversity and habitat distribution of the gastropods, we collected the freshwater snail samples from 62 sites in the littoral zone of Poyang Lake in May and June of 2011, and investigated the characteristics of their habitats. Nineteen freshwater snails species were identified, which were categorized into viviparidae (8 species), Hydrobiidae (8 species), Lymnaeidae (2 species) and Melaniidae (1 species). Compared to the previously reported data the number of taxa showed a sharp decline. Similar to the previous reports, viviparidae and Hydrobiidae were still the dominant species in the community. There were remarkable differences in species composition between mud and other habitats. The species richness of freshwater snails showed significant difference between mud and macrophyte. The differences were also presented between mud and sandy soil, and between mud and stone (P<0.05). These indicated that snails had a perching preference in selecting their habitats. Finally, we discussed the factors that potentially influence species diversity and habitat distribution of freshwater snails.
Poyang Lake; Freshwater snails; Species diversity; Habitat type
Q145+.1
A
1000-3207(2014)03-0407-07
10.7541/2014.58
2013-04-07;
2013-12-14
“鄱阳湖国家级自然保护区二次科考”(环保部)项目资助
严家跃(1987— ), 男, 广东连州人; 硕士研究生; 主要从事淡水螺类生态学研究。E-mail: 10210700128@fudan.edu.cn
陈家宽, E-mail: jkchen@fudan.edu.cn
