香樟人工林土壤表层细根形态特征、生物量及碳氮含量变化
2014-03-26张腾飞李贤伟刘运科杨正菊
张腾飞,李贤伟,2,范 川,魏 鹏,刘运科,苏 宇,杨正菊
(1 四川农业大学 林学院,四川 雅安 625014;2 长江上游林业生态工程四川省重点实验室,四川 雅安 625014)
根系是植物与土壤的动态界面,具有支持、吸收、合成和储存等功能,在森林生态系统的养分循环、地力维持及土壤结构改善中起着重要作用[1]。细根通常是指直径小于2 mm的根,是植物体获取土壤物质和能量的主要器官,也是植物地下最活跃、对土壤环境影响反应最敏感的部分[2]。细根生物量虽然只占地下总生物量的3%~30%[3],但由于它处于较快的周转过程之中,维持这一动态过程要消耗净初级生产力的10%~75%[2,4]。研究表明,植物根系也像地上枝条一样,在发育过程中形成了不同的分枝结构即根序,不同序级的细根具有不同的形态结构和生理功能。细根直径、比根长(SRL)和根长密度(RLD)是描述细根形态结构的重要参数,细根形态结构不同,其养分含量也有显著差异,从而影响细根的生理、生态功能[5]。以往的研究主要通过生物量的大小来评价根系吸收养分和水分的能力,认为根系生物量越高,吸收的养分和水分就越多[6-7],但具有不同根序等级的细根结构和功能不完全相同,因此以单一的生物量来判断细根对养分和水分的吸收能力并不科学。因而以不同根序的SRL、RLD和生物量来衡量细根吸收养分及水分的能力,可能更具实际意义。
人工林不仅是我国经济建设所需木材的主要来源,而且有研究表明,我国近20多年来的森林碳汇主要来自人工林的贡献[8]。随着人工林面积和蓄积量的持续增加,人工林在全球碳循环及减缓气候变化中的地位举足轻重。香樟(Cinnamomumcamphora)作为重要的园林和经济树种,长期以来,对其栽培技术、病虫害防治、生物多样性、快速繁殖、挥发油化学成分等方面的研究颇多[9-10],但是对于香樟人工林的根系研究还很少,几乎还是一个空白领域[11]。为此,本试验以香樟人工林为研究对象,探讨了香樟1~5级细根形态特征、生物量及碳、氮含量的变化,旨在为深入研究人工林细根对养分和水分的吸收规律,以及碳在地下的分配特点提供基础资料。
1 研究地概况与研究方法
1.1 研究地概况及样地设置
研究地位于四川省雅安市雨城区老板山实习林场(29°54′02″N,103°01′46″E),海拔620 m,该地属亚热带湿润季风气候,夏无酷暑,春季回暖,降水集中于夏季,多夜雨,年均温16.2 ℃,1月均温为6.1 ℃,7月均温为25.4 ℃,年降水量可达2 000 mm。林分为31年生香樟纯林,平均胸径为21 cm,平均树高14 m,南北冠幅4 m,东西冠幅5 m,郁闭度0.7,植株密度为720株/hm2,坡向为北坡,坡度5°,土壤类型为紫色土,土壤性质见表1。
在香樟人工林中随机选取3个面积为20 m×20 m的试验样地,由于人为干扰,林下植被只有少量鸢尾(Iristectorum)、接骨草(Sambucuschinensis)、土牛膝(Achyranthesaspera)和蕨类。
1.2 研究方法
1.2.1 细根样品的采集与处理 于2011年的1,4,7和10月份,分别在各样地按“S”形选取5个采样点,用15 cm×15 cm×10 cm的根系取样器分2层取根系样品,每层10 cm,每次取样30个。带回实验室后,用低温(2~4 ℃)去离子水轻轻洗去细根表面的土壤颗粒及其他残留物等杂质,挑出完整的细根根段,放入盛有低温去离子水的玻璃皿中。用Fitter等[12]的根系序列位置命名方法对细根进行分级,位于根轴最远端没有分枝的根称为1级根,1级根着生在2级根上,依次分到5级根,位于高级根上没有分枝的根也视为1级根。将分级后的各样地不同根序等级细根分别放入已标记的玻璃皿中。
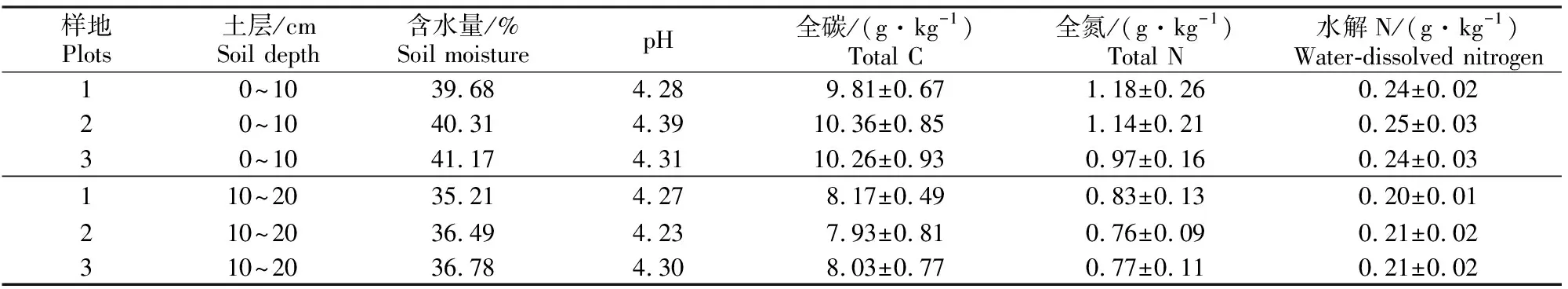
表1 四川雅安香樟人工林试验地土壤性质
1.2.2 细根形态和生物量的测定 用Epson数字化扫描仪(Expression 10000XL 1.0)和WinRHIZO 2005根系图像分析系统软件,对不同根序级细根的形态特征指标进行定量分析。扫描后将各根序级细根放入65 ℃烘箱中烘至恒质量,测定其干质量。根据WinRHIZO 2005记录的根长和细根直径等数据,结合测定的细根生物量,计算细根的SRL、RLD和细根直径。SRL和RLD的计算公式为:
SRL(m/g)=根长(m)/生物量(g)。
RLD(m/m3)=根长(m)/土样体积(m3)。
1.2.3 细根C、N含量的测定 将烘干的各序级细根样品磨碎,过孔径2 mm筛,然后取样并装袋保存。有机C用重铬酸钾外加热法测定(LY/T 1237-1999),全N用蒸馏法测定(LY/T 1269-1999)。
1.3 数据处理与统计分析
数据处理及图表生成采用Microsoft Excel 2007软件,统计分析用SPSS 17.0软件进行。用One-way ANOVA对各根系指标在不同季节、不同根序和不同土层深度的差异性做方差分析。
2 结果与分析
2.1 不同季节及土层香樟细根直径、SRL和RLD的比较
不同季节及土层香樟细根直径、SRL、RLD与生物量测定结果见表2和表3。根序、季节及二者交互作用对香樟4个指标影响的方差分析结果见表4。
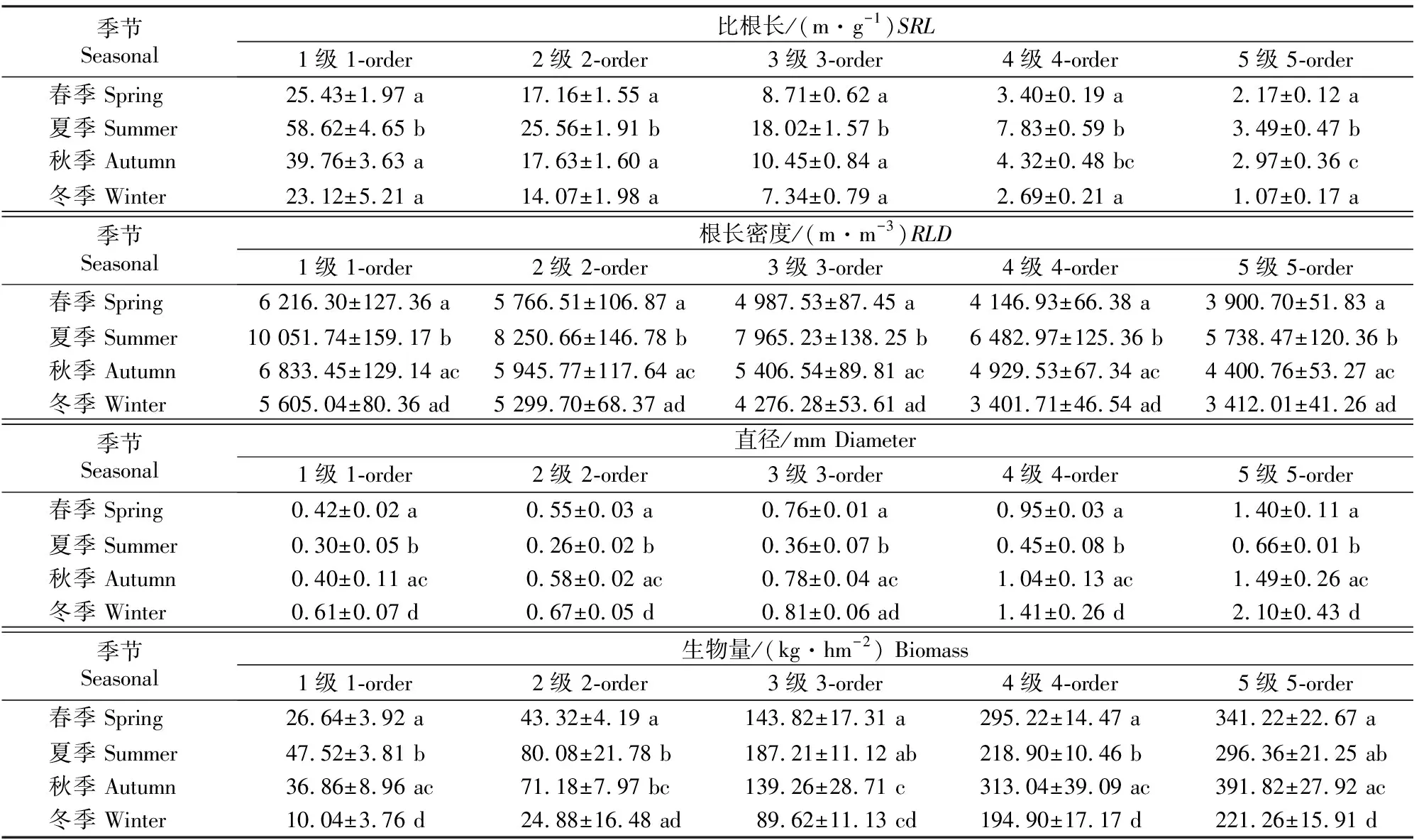
表2 四川雅安4个季节香樟细根的平均直径、比根长、根长密度和生物量
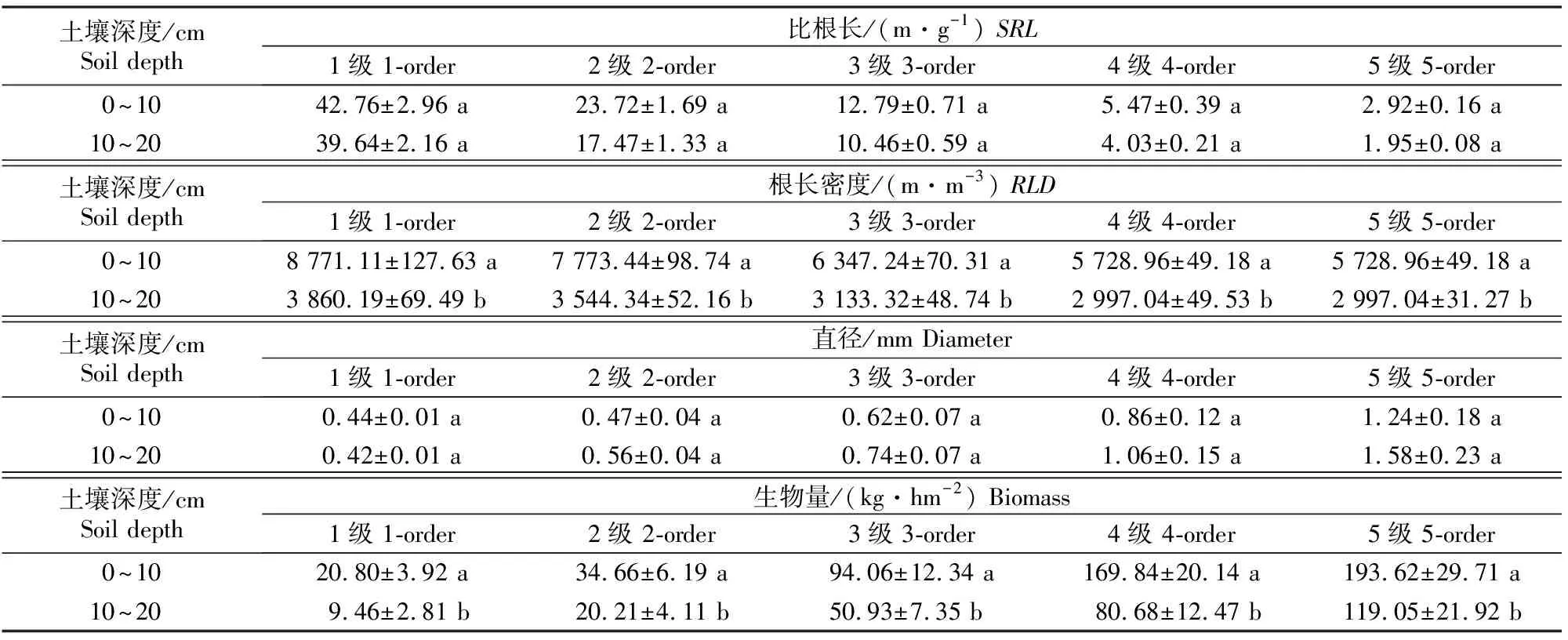
表3 四川雅安不同土层香樟细根的平均直径、比根长、根长密度和生物量
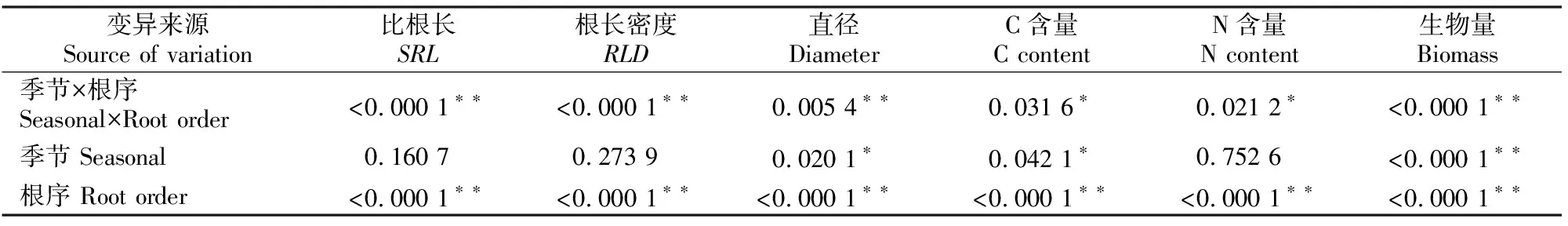
表4 根序和季节对香樟细根SRL、RLD、直径及C、N含量影响的方差分析
由表2可以看出,夏季香樟细根直径、SRL和RLD与其他季节存在显著差异(P<0.05),1~5级根直径夏季最细,显著小于其他季节(P<0.05),冬季最粗,春季和秋季差异很小;1~5级根SRL和RLD均以夏季最高,显著高于其他季节(P<0.05),冬季最低。除1级根外,其他根序级根直径均表现为冬季>秋季>春季>夏季;1~5级根的SRL均表现为夏季>秋季>春季>冬季;1~5级根的RLD均表现为夏季>秋季>春季>冬季。同一季节的细根直径随着根序的升高而增大,SRL和RLD随着根序的升高而减小(表2),且不同根序间均达到极显著差异(P<0.01)(表4),表明不同细根根序之间直径、SRL和RLD表现出的差异比季节之间的差异更为明显。季节与根序的交互作用对细根直径、SRL和RLD均有极显著影响。1~5级根的直径在0~10 cm,10~20 cm土层间均表现为随根序增加逐渐增大,而SRL和RLD则随根序的增加逐渐减小;直径和SRL在土层间未达到显著性差异(P>0.05),RLD则均以0~10 cm土层显著高于10~20 cm土层 (P<0.05)(表3)。
2.2 不同季节及土层香樟细根生物量的比较
由表3可知,不同根序香樟细根的生物量在 0~10 cm,10~20 cm土层间的变化规律相同,表现为0~10 cm土层细根生物量显著高于10~20 cm土层(P<0.05)。随着根序的升高,0~10 cm土层中1~5级细根生物量分别占细根总生物量的 3.85%,9.36%,18.14%,30.56%和38.09%;10~20 cm土层中1~5级细根生物量分别占细根总生物量的3.05%,8.26%,17.10%,31.51%和40.09%;0~20 cm土层中1~5级细根生物量分别占细根总生物量的2.28%,6.39%,15.61%,30.66%和45.06%。表2表明,1~2 级细根生物量夏季最高,冬季最低;3级细根生物量秋季最高,冬季最低;4、5级细根生物量秋季最高,冬季最低。
2.3 不同季节及土层香樟细根C、N含量的比较
如图1所示,香樟1级根的C含量最小,为124.26~447.32 g/kg,随着根序的增加,细根C含量也随之增大(冬季除外),各季节1~3级细根间C含量的变化幅度大于4、5级根,不同根序间细根C含量表现出极显著差异(P<0.01)(表3);不同季节间C含量表现出显著差异(P<0.05)(表3),冬季细根C含量最高,夏季最低;除夏季外,0~10 cm土层细根C含量均高于10~20 cm土层细根。
由图1还可以看出,香樟细根全N含量表现为随着根序的升高而降低,1级根的N含量最高,为2.76~4.87 g/kg,5级根最低,为1.73~3.61 g/kg,不同根序间N含量表现为极显著差异(P<0.01)(表3)。不同季节间细根N含量差异不显著(P>0.05)(表3),1~3级根N含量以夏季最高,春季最低,4、5级根N含量在0~10 cm土层中表现为夏季最高,冬季最低,在10~20 cm土层中表现为秋季最高,春季最低。
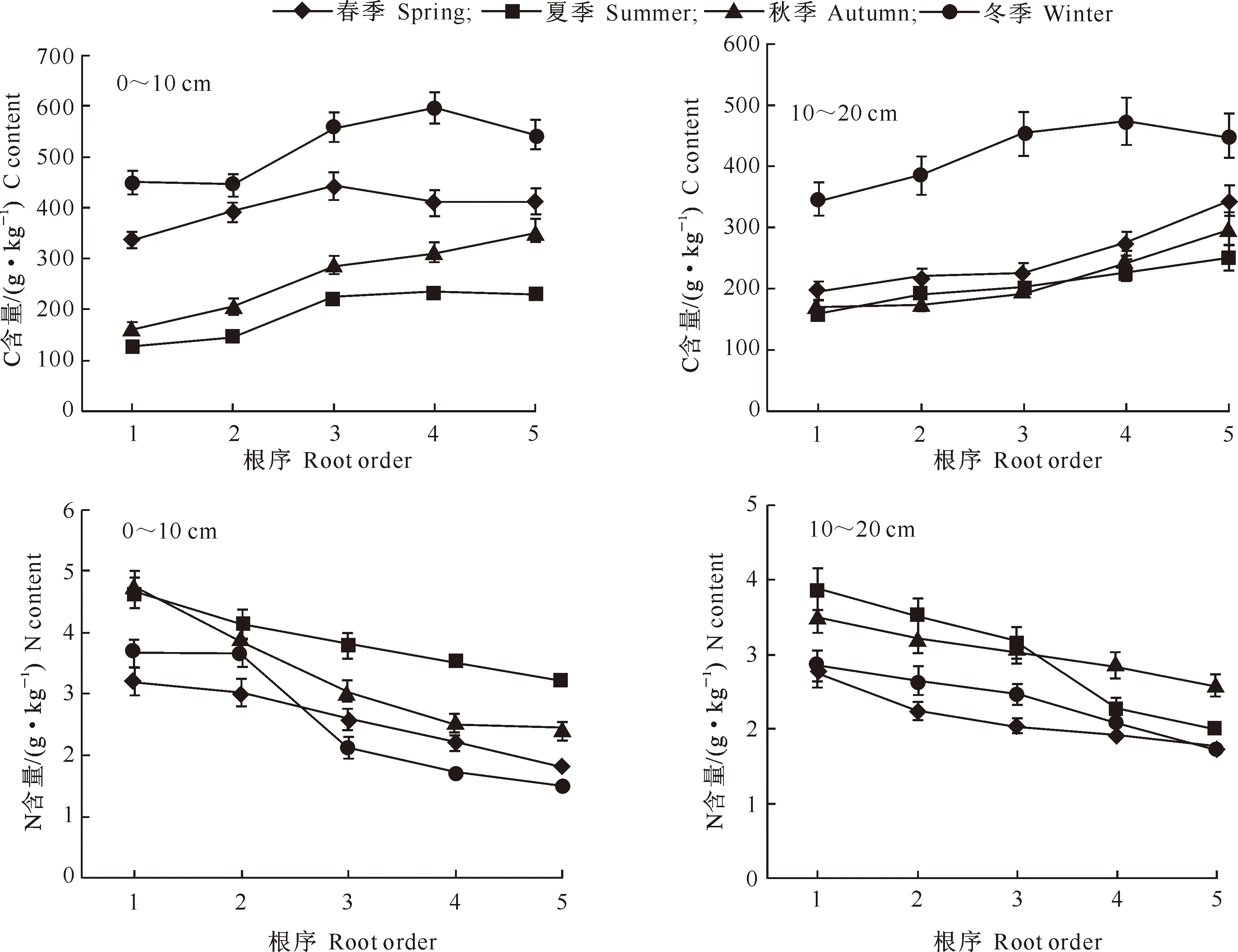
图1 四川雅安不同季节及土层1~5级香樟细根的C、N含量
3 讨 论
3.1 不同季节及土层香樟细根的直径、SRL和RLD
树木根系在长期进化过程中形成分枝(Root branching)结构,这种结构可以使根系分工明确,提高吸收养分和水分的效率,避免不良环境下个别根死亡后殃及整个根系[13]。细根直径是反映细根结构、功能的重要参数,直径大小影响细根的寿命和吸收功能[14]。香樟细根直径随根序的增大而增加,这与前人对细根直径的研究结果[13,15]一致,说明细根直径可能具有这种普遍的变化规律。大多数研究都将根直径<2 mm作为同一个单元进行分析,但本研究结果表明,1级根平均直径为0.43 mm,5级根平均直径为1.41 mm,直径<2 mm的细根包括了 1~5级根序,因而将其作为1个单元,忽略了细根形态和功能的异质性,而采用根序则能较好地预测细根结构和功能的关系[16]。不同季节香樟细根直径变化明显,夏季平均直径为0.41 mm,冬季平均直径为1.12 mm,这是因为不同季节土壤水分、温度和养分存在明显变化,细根作为对土壤变化最敏感的部分,会对土壤资源有效性的变化作出可塑性反应[17]。
SRL反映投入到细根的生物量用于吸收养分的效率和细根吸收养分及水分的能力,其值越大,表明投入相同的成本有越高的养分利用效率[18]。RLD反映细根对土壤中水分和养分的吸收效率,其值越高,效率越高,是根系对土壤空间异质性的基本反映[19]。本研究结果显示,夏季香樟1~5级根的SRL和RLD显著高于其他季节,主要是因为土壤资源有效性较高,而1~3级根木质化程度较低,对环境的变化非常敏感[20],此时细根的纵向延伸比增粗更有意义[21];由于1~3级根数量的增加,支撑 1~3级根的4、5级根就会显著增加,以满足细根对养分的吸收和运输[22]。香樟1~3级根的SRL和RLD显著高于同季节4、5级根,1~3级根的SRL占1~5级根的90.0%~95.8%,而1~3级根的RLD占1~5级根的70%,可见细根中1~3级根主要起吸收作用[23]。香樟1~3级根SRL和RLD所占比例如此之高,说明细根构建和维持单位长度消耗的碳水化合物较少,对行使细根的吸收功能有利[24],保证了林木水分、养分的供应和正常生长,反映了细根功能的异质性。
0~10,10~20 cm土层细根直径逐渐增加,SRL和RLD逐渐减少,0~10 cm、10~20 cm土层间RLD存在显著差异,是因为RLD对土壤肥力变化的敏感性显著高于SRL[25],因此对0~10 cm、10~20 cm土层土壤肥力的变化,RLD表现出显著性差异。
3.2 不同季节及土层香樟细根的生物量
细根生物量的多少是植物对土壤资源利用有效性的反映及地上光合产物在地下分配的最终体现[26]。细根生物量在1年中常出现1或2个峰值,峰值大多出现在春季展叶期前后、晚夏或秋季等[27]。本研究结果显示,香樟细根生物量峰值出现在春季和秋季,夏季和冬季较低。而1~2级根的生物量在夏季最高,冬季最低。夏季树木地上部分生长旺盛,光合产物更多用于地上部分生长和养分、水分吸收,只有少部分分配到地下根系用于细根构建,与高级根相比,低级根能有效地吸收土壤内的养分和水分,而在形成和维持生理功能方面只需投入较少的C[28],所以在地下根系C分配有限时,根据植物“投入”与“产出”效益理论[29],会分配更多的C用于低级根的构建,以满足地上部分对养分和水分的需求。秋季地上部分生长缓慢,因而地上部分累积的C主要用于地下部分生长,以便在高级根中储存更多的碳水化合物用于下一年展叶之前较长一段时间根系生长的需求[30],故4、5级根的生物量在秋季显著增加。进入冬季后,环境因子不利于根系活动,导致冬季细根死亡量增加[31],所以冬季细根生物量较低。
本研究结果表明,0~10 cm、10~20 cm土层香樟各级细根生物量变化趋势相同,均以0~10 cm土层显著高于 10~20 cm土层。Jackson等[32]认为,土壤空间异质性是导致根系分布空间异质性的主要原因,根系对土壤空间异质性的基本反应是调整生物量。根系生物量在肥沃的0~10 cm土层分布较多,而在相对贫瘠的10~20 cm土层分布较少。根系吸收机制是在比较肥沃的土层中尽可能投入较多的碳水化合物,尽可能多地吸收养分和水分[25]。
3.3 不同季节及土层香樟细根的C、N 含量
本研究发现,香樟细根C含量随着根序的增大而增加,细根N含量随着根序的增加而降低,这与以前的大部分研究结果[7,23,33]相似。香樟各级根序间细根C、N含量差异显著,表明根序级越低,对土壤有效性的敏感性越强,从根系内部解剖结构上看, 低级根(1级根)主要由初生组织构成,皮层细胞是其结构中主要的细胞类型[34]。1级根的N含量最高,C含量最低,这种养分分配格局与1级根的功能密切相关,作为细根最活跃的部分,1级根主要起到吸收养分和水分的作用。细根吸收养分和水分的同时,需要形态变化的支持,吸收养分和水分时消耗的C也需要高级根的供给,因此,细根形态和营养元素都与根序存在高度的相关性[33]。可见,划分根序对细根进行研究具有重要的意义。本研究中,夏、秋季细根C含量要明显低于春、冬季,而夏、秋季细根N含量要明显高于春、冬季,这主要是因为夏季和秋季是水分充足、温度也较高的季节,土壤中有效N的含量也较高,细根不断吸收土壤中的有效N,同时不断消耗地上部分固定且运输到细根的碳水化合物,这就使得细根中的N素积累,而C却逐渐被消耗;到了冬季,土壤中有效N含量下降,细根吸收的N含量也随之降低,进而细根呼吸消耗的C也减少,这时地上部分运输到细根的C就会积累,细根直径增粗以储存更多的C;春季香樟开始生长并消耗细根中的C,加快了细根的死亡和分解,将细根中的营养元素回馈给土壤,用于香樟又一次的生长[35]。
本研究结果表明,0~10 cm、10~20 cm土层细根C、N含量变化趋势相同,但变化很小,这种变化主要是由土壤资源有效性的差异引起的。
4 结 论
香樟细根随着序级的增加,其直径、生物量和C含量逐渐增大,1级根最小,5级根最大;而N含量、SRL和RLD逐渐减小,1级根最大,5级根最小。1~3级根与4、5级根的形态结构和养分含量有很大差别,具有吸收功能的1~3级根拥有较大的SRL和RLD以及较高的N含量,4、5级根拥有较粗的直径和较高的C含量、生物量。夏季细根趋向于吸收养分和水分,用于1~3级根生长的生物量也高于4、5级根,此时细根形态和养分也向着有利于吸收的方向变化,这可能是植物对生长环境的一种适应性变化。
研究不同根序细根形态和功能特征可以很好地了解形态与养分策略之间的关系,对认识细根形态特征和化学成分的变异格局,及其对森林生态系统功能过程的影响具有重要的意义。
[参考文献]
[1] 张小全,吴可红.森林细根生产和周转研究 [J].林业科学,2001,37(3):126-138.
Zhang X Q,Wu K H.Fine-root production and turnover for forest ecosystems [J].Scientia Silvae Sinicae,2001,37(3):126-138.(in Chinese)
[2] 梅 莉,王政权,张秀娟,等.施氮肥对水曲柳人工林细根生产和周转的影响 [J].生态学杂志,2008,27(10):1663-1668.
Mei L,Wang Z Q,Zhang X J,et al.Effects of nitrogen fertilization on fine root biomass production and turnover ofFraxinusmandshuricaplantation [J].Chinese Journal of Ecology,2008,27(10):1663-1668.(in Chinese)
[3] 裴智琴,周 勇,郑元润,等.干旱区琵琶柴群落细根周转对土壤有机碳循环的贡献 [J].植物生态学报,2011,35(11):1182-1191.
Pei Z Q,Zhou Y,Zheng Y R,et al.Contribution of fine root turnover to the soil organic carbon cycling in aReaumuriasoongoricacommunity in an arid ecosystem of Xinjiang Uygur Autonomous Region China [J].Chinese Journal of Plant Ecology,2011,35(11):1182-1191.(in Chinese)
[4] Gill R A,Jackson R B.Global patterns of root turnover for terrestrial ecosystems [J].New Phytologist,2000,147(3):13-31.
[5] 许 旸,谷加存,董雪云,等.海南岛4个热带阔叶树种前5级细根的形态、解剖结构和组织碳氮含量 [J].植物生态学报,2011,35(9):955-964.
Xu Y,Gu J C,Dong X Y,et al.Fine root morphology,anatomy and tissue nitrogen and carbon contents of the first five orders in four tropical hardwood species in Hainan Island,China [J].Chinese Journal of Plant Ecology,2011,35(9):955-964.(in Chinese)
[6] Burton A J,Pregitzer K S,Hendrick R L.Relationships betwe-en fine root dynamics and nitrogen availability in Michigan northern hardwood forests [J].Oecologia,2000,125:389-399.
[7] Pregitzer K S,De Forest J L,Burton A J,et al.Fine root architecture of nine North American trees [J].Ecological Monographs,2002,72:293-309.
[8] Fang J Y,Chen A P.Dynamic forest biomass carbon pools in China and their significance [J].Acta Botanica Sinica,2001,43(9):967-973.
[9] 刘玉红,赵鹏华,王义华.香樟栽植技术 [J].林业科学,2009,45(5):164-167.
Liu Y H,Zhao P H,Wang Y H.Cinnamomumcamrphoacultivation techniques [J].Scientia Silvae Sinicae,2009,45(5):164- 167.(in Chinese)
[10] 高金根.香樟病虫害综合防治技术 [J].安徽林业,2009,5(2):58-59.
Gao J G.Comprehensive control techniques of diseases and insect pests ofCinnamomumcamphora[J].Anhui Forestry,2009,5(2):58-59.(in Chinese)
[11] 闫德祺,禹明甫,刘德成,等.香樟耐寒性试验研究 [J].安徽农业科学,2007,35(33):78-80.
Yan D Q,Yu M F,Liu D C,et al.Research of cold endurance ofCinnamomumcamphora[J].Anhui Agricultural Sciences,2007,35(33):78-80.(in Chinese)
[12] Fitter A H,Aiinson D,Read D J,et al.Ecological interaction in soil [M].Oxofrd:Black Wells,1985:87-106.
[13] 熊德成,黄锦学,杨智杰,等.亚热带6种树种细根序级结构和形态特征 [J].生态学报,2012,32(6):1888-1897.
Xiong D C,Huang J X,Yang Z J,et al.Fine root architecture and morphology among different branch orders of six subtropical tree species [J].Acta Ecologica Sinica,2012,32(6):1888-1897.(in Chinese)
[14] Robinson D.The responses of plants to nonuniform supplies of nutrients [J].New Phytologist,1994,127(4):635-674.
[15] 常文静,郭大立.中国温带、亚热带和热带森林45个常见树种细根直径变异 [J].植物生态学报,2008,32(6):1248-1257.
Chang W J,Guo D L.Variation in root diameter among 45 common tree species in temperate,subtropical and tropical forests in China [J].Chinese Journal of Plant Ecology,2008,32(6):1248-1257.(in Chinese)
[16] Fitter A H,Stickland T R.Architectural analysis of plant root systems:Studies on plants under field conditions [J].New Phytologist,1992,121(2):243-248.
[17] Hodge A.The plastic plant:Root responses to heterogeneous supplies of nutrients [J].New Phytologist,2004,162(1):9-24.
[18] Bauhus J,Khanna P K,Menden N.Aboveground and belowground interactions in mixed plantations ofEucalyptusglobulusandAcaciamearnsii[J].Canadian Journal of Forest Research,2000,30:1886-1894.
[19] 梅 莉,王政权,韩有志,等.水曲柳根系生物量、比根长和根长密度的分布格局 [J].应用生态学报,2006,17(1):1-4.
Mei L,Wang Z Q,Han Y Z,et al.Distribution patterns ofFraxinusmandshuricaroot biomass,specific root length and root length density [J].Chinese Journal of Applied Ecology,2006,17(1):1-4.(in Chinese)
[20] 于立忠,丁国泉,史建伟,等.施肥对日本落叶松人工林细根直径、根长和比根长的影响 [J].应用生态学报,2007,18(5):957-962.
Yu L Z,Ding G Q,Shi J W,et al.Effects of fertilization on fine root diameter,root length and specific root length inLarixkaempferiplantation [J].Chinese Journal of Applied Ecology,2007,18(5):957-962.(in Chinese)
[21] 马晓东,朱成刚,李卫红.多枝柽柳幼苗根系形态及生物量对不同灌溉处理的响应 [J].植物生态学报,2012,36(10):1024-1032.
Ma X D,Zhu C G,Li W H.Response of root morphology and biomass ofTamarixramosissimaseedlings to different water irrigations [J].Chinese Journal of Plant Ecology,2012,36(10):1024-1032.(in Chinese)
[22] 王向荣,谷加存,梅 莉,等.水曲柳和落叶松细根形态及母根与子根比例关系 [J].生态学报,2006,26(6):1686-1692.
Wang X R,Gu J C,Mei L,et al.Fine root order morphology and proportion between mother roots and daughter roots inFraxinusmandshuricaandLarixgmeliniiplantations [J].Acta Ecologica Sinica,2006,26(6):1686-1692.(in Chinese)
[23] Guo D L,Mitchell R J,Hendricks J J.Fine root branch orders respond differentially to carbon source-sink manipulations in a longleaf pine forest [J].Oecologia,2004,140:450-457.
[24] 王向荣,王政权,韩有志,等.水曲柳和落叶松不同根序之间细根直径的变异研究 [J].植物生态学报,2005,29(6):871-877.
Wang X R,Wang Z Q,Han Y Z,et al.Variations of fine root diameter with root order in Manchurian ash and Dahurian larch plantations [J].Chinese Journal of Plant Ecology,2005,29(6):871-877.(in Chinese)
[25] Fransen B,Kroon H D,Berendse F.Root morphological plasticity and nutrient acquisition of perennial grass species from habitats of different nutrient availability [J].Oecologia,1998,115:351-358.
[26] Eissenstat D M,Yanai R D.The ecology of root lifespan [J].Advances in Ecological Research,1997,27:1-60.
[27] 杜有新,潘根兴,李恋卿,等.贵州中部喀斯特山地不同植被生态系统细根生态特征及养分储量 [J].应用生态学报,2010,21(8):1926-1932.
Du Y X,Pan G X,Li L Q,et al.Fine root biomass and its nutrient storage in karst ecosystems under different vegetations in Central Guizhou,China [J].Chinese Journal of Applied Ecology,2010,21(8):1926-1932.(in Chinese)
[28] Ryan M G,Hubbard R M,Pongracic S,et al.Foliage,fine-root,woody-tissue and stand respiration inPinusradiatain relation to nitrogen statu [J].Tree Physiol,1996,16:333-343.
[29] 刘金梁,梅 莉,谷加存,等.内生长法研究施氮肥对水曲柳和落叶松细根生物量和形态的影响 [J].生态学杂志,2009,28(1):1-6.
Liu J L,Mei L,Gu J C,et al.Effects of nitrogen fertilization on fine root biomass and morphology ofFraxinusmandshuricaandLarixgmelinii:A study with in-growth core approach [J].Chinese Journal of Ecology,2009,28(1):1-6.(in Chinese)
[30] Pregitzer K S.Woody plants,carbon allocation and fine roots [J].New Phytologist,2003,158(3):421-424.
[31] 杨丽韫,罗天祥,吴松涛.长白山原始阔叶红松(Pinuskoraiensis)林及其次生林细根生物量与垂直分布特征 [J].生态学报,2007,27(9):3609-3617.
Yang L Y,Luo T X,Wu S T.Fine root biomass and its depth distribution across the primitive Korean pine and broad-leaved forest and its secondary forests in Changbai Mountain,northeast China [J].Acta Ecologica Sinica,2007,27(9):3609-3617.(in Chinese)
[32] Jackson R B,Canadell J R,Mooney H A,et al.A global analysis of root distribution for terrestrial biomass [J].Oecologia,1996,108:389-411.
[33] 刘 佳,项文化,徐 晓,等.湖南会同5个亚热带树种的细根构型及功能特征分析 [J].植物生态学报,2010,34(8):938-945.
Liu J,Xiang W H,Xu X,et al.Analysis of architecture and functions of fine roots of five subtropical tree species in Huitong,Hunan Province,China [J].Chinese Journal of Plant Ecology,2010,34(8):938-945.(in Chinese)
[34] Guo D L,Xia M X,Wei X,et al.Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species [J].New Phytologist,2008,180(12):673-683.
[35] Pregitzer K S,Kubiske M E,Yu C K,et al.Relationships among root branch order,carbon,and nitrogen in four temperate species [J].Oecologia,1997,111:302-308.
