长白山阔叶红松林典型森林群落功能多样性及其与地形因子的关系
2014-03-26么旭阳胡耀升刘艳红
么旭阳,胡耀升,刘艳红
(北京林业大学 林学院,森林培育与保护省部共建教育部重点实验室,北京 100083)
植物功能多样性(Functional diversity)与生态系统功能的关系一直是近年来研究的热点。研究内容主要包括植物功能多样性对资源动态的短期效应与生态系统或群落生产力的关系,以及植物功能多样性对生态系统稳定性的长期效应等。而在实际研究中,关于植物功能特征的选择、试验设计、试验因素的控制、结果的统计分析以及尺度选择等,均对研究结果有重要影响,因此产生了不同的理论和机制[1]。目前,研究者较为广泛接受的一个功能多样性的定义是:特定生态系统中物种功能特征的数值和范围,也叫功能特性多样性(Functional trait diversity,FTD),主要强调群落中物种功能的差异[2-3]。据此可见,功能多样性是生物多样性的最重要的组成部分,也能够反映生态系统功能的重要特征[1,4-5]。目前已有几种功能多样性指标测量方法,如Rao’s Q指数[6]、CWM指数[7]、功能丰富度指数[4]、功能均匀度指数[8]等,且各种功能多样性指数的计算都需要在物种特征和物种丰富度数据的基础上进行。
生态系统的功能多样性可用体现群落内不同特征值分布状况的指数来定量描述,因此,简单的物种多样性指标难以真实地体现物种特征对生态系统过程的重要作用。生态系统功能不仅依赖于物种的数目, 而且依赖于物种所具有的功能特征[9-10]。近年来,基于植物功能特征的物种功能多样性研究方法在生态学中受到重视,有不少研究开始这一方面的尝试[11-12]。对于植物功能特征的研究,多是在探讨植物功能性状特征及其与主要环境因子的相关性[13-14],而关于群落功能多样性指数与主要地形因子关系的研究鲜见报道。同时,以典型群落为基础,探讨群落功能与生态系统功能间的关系,也是森林合理经营的基础[15]。
长白山阔叶红松林为北温带地区特有的森林类型,目前,世界范围内的生物多样性日益减少,森林资源利用效益备受关注,天然红松林面积逐步缩小,大面积森林逐步为次生林和人工林所代替[16]。基于此,本试验通过标准固定样地调查,对长白山典型森林群落的功能多样性和物种多样性进行研究,探讨影响群落功能多样性指数的主要环境因子,初步揭示植物群落功能多样性与群落重要生态过程之间的关系,以期为该区植被恢复和生态系统保护提供理论与技术支持。
1 研究地概况与研究方法
1.1 研究地概况
研究区位于长白山自然保护区北坡的阔叶红松林内,该地区受海洋性气候影响较大,气候温和湿润,年均降水量700 mm, 多集中在6-8月(480~500 mm)。温度由北向南呈逐渐升高的趋势,冬、夏季温差大,年平均气温3.6 ℃,最冷月(1月份)平均气温-15.6 ℃,最热月(7月份)平均气温19.7 ℃。长白山北坡的阔叶红松林地坡度平缓,土壤以深厚的暗棕色森林土为主。由于该地区林分结构复杂,多形成复层混交异龄林。森林主要树种有紫椴(Tiliaamurensis)、红松(Pinuskoraiensis)、蒙古栎(Quercusmongolica)、色木槭(Acerpictumsubsp.mono)、水曲柳(Fraxinusmandshurica)、白桦(Betulaplatyphylla)和山杨(Populusdavidiana)等。
1.2 调查方法
1.2.1 样地设置 于2012-07-08,对杨桦林、白桦林、臭冷杉红松林以及紫椴红松林等4种典型森林群落进行样方调查。样地海拔700~1 100 m,绝对高差280.3 m,立地条件较好,每种群落随机选择4个20 m×20 m的样地,在样地中进行群落学调查。对乔木进行每木检尺,记录树高、胸径、冠幅,并将样地分成16个5 m×5 m的单元格,在坐标纸上标记每棵树的位置。在样地的4个角距边缘1 m处各设1个5 m×5 m的灌木样方,调查灌木的种类、株数、株高和盖度;在每个灌木大样方的4个角再设置4个1 m×1 m的草本样方,调查草本的种类、株高和盖度。记录每个样地的海拔高度、坡度、坡位和坡向(表1)。坡位数据1表示上坡位,2表示中坡位,3表示下坡位。坡向数据是以朝东为起点(即为0°)顺时针旋转的角度表示,采取每45°为一个区间的划分方法,以数字表示各等级:1表示北坡(247.5°~292.5°),2表示东北坡(292.5°~337.5°),3表示西北坡(202.5°~247.5°),4表示东坡(337.5°~22.5°),5表示西坡(157.5°~202.5°),6表示东南坡(22.5°~67.5°),7表示西南坡(112.5°~157.5°),8表示南坡(67.5°~112.5°),即各等级数字越大,表示越向阳,越干热[17]。

表1 长白山阔叶红松林内4种典型森林群落样地的立地状况
1.2.2 物种重要值的计算 计算每个样方的物种重要值,采用综合指标物种重要值来反映物种特征。
乔木树种的重要值=(相对多度+相对显著度+相对频度)/3;
灌木(草本)植物的重要值=(相对密度+相对盖度+相对高度)/3。
1.3 数据分析与处理
1.3.1 群落内物种多样性指数 (1)物种丰富度(S)。S=样地内的物种数。
(2)Shannon-Winner多样性指数。
式中:S为群落内总物种数,Pi为第i物种个体数与所有物种总数的比值。
计算各样点内不同样地的物种丰富度、Shannon-Winner指数的平均值,得到各样点不同阔叶红松林群落的上述指数的样地平均值[18]。
1.3.2 群落功能多样性指数 植物群落功能多样性指数可对植物群落内物种的功能特征进行量化描述,是直观分析植物群落功能多样性的基础。本研究通过外业调查选取植物功能性状,主要包括比叶面积(SLA)、叶厚度、茎组织密度、植物叶片和枝的N、P含量[5]。
本研究选取应用广泛、计算较为简便的Rao’s Q指数对群落的功能多样性进行量化分析。
Rao’s Q指数即利用物种特性计算种间差异性,如果第i个物种在群落中的比例是Pi,物种i与物种j的相异性是d,则Rao’s Q(FD)指数的表达式为:

式中:dij为物种i与j的差异,n为性状数,Xik为物种i的k性状值,Xjk为物种j的k性状值,dij变化于0 (两物种具有完全相同的特征)与1(两物种具备完全不同的特征)之间[6];S为群落内总物种数,Pi和Pj分别为物种i和物种j的个体数占群落内总物种数的比例。
1.3.3 灰色关联度分析 用DPS软件对16块样地的地形因子与功能多样性指数和物种多样性指数进行灰色关联度分析。数据输入过程中,各项地形因子如海拔、坡度数据均以实际观测记录值表示。
2 结果与分析
2.1 长白山阔叶红松林内4种典型森林群落的多样性指数
2.1.1 不同群落内的物种多样性指数 阔叶红松林内4种典型森林群落的Shannon-Winner指数平均值见图1。由图1可知,就群落整体而言,阔叶红松林各群落的物种多样性一般为草本层>灌木层>乔木层。而就不同植物的生活型而言,乔木层和草本层的Shannon-Winner指数排序均为:白桦林>臭冷杉红松林>杨桦林>紫椴红松林;灌木层的Shannon-Winner指数排序为白桦林>杨桦林>臭冷杉红松林>紫椴红松林。
2.1.2 不同群落的功能多样性指数 由图2可以看出,乔木层和灌木层各群落Rao’s Q指数排序均为紫椴红松林>臭冷杉红松林>杨桦林>白桦林;草本层Rao’s Q指数排序为白桦林>紫椴红松林>臭冷杉红松林>杨桦林。

图1 长白山阔叶红松林内4种典型森林群落的Shannon-Winner指数
2.2 长白山阔叶红松林内4种典型森林群落整体的物种多样性和功能多样性
根据群落内物种丰富度,计算并对比分析4种典型森林群落整体的物种多样性和功能多样性,结果(表2)显示,其物种多样性Shannon-Winner指数排序为臭冷杉红松林>白桦林>紫椴红松林>杨桦林,其功能多样性Rao’s Q指数排序为紫椴红松林>臭冷杉红松林>杨桦林>白桦林。

表2 长白山阔叶红松林内4种典型森林群落整体的物种多样性和功能多样性
2.3 长白山阔叶红松林内典型森林群落的地形因子与其多样性指数的关系
选取功能多样性指数和物种多样性指数作为因变量,以海拔、坡度、坡位、坡向4个地形因子为自变量,用16块样地的实测数据进行灰色关联度分析。灰色关联度值越大,表明自变量数列对因变量的影响越大[19]。由表3可以看出,影响S和Rao’s Q指数的各地形因子排序均为坡度>海拔>坡向>坡位;影响H′的各地形因子排序为海拔>坡向>坡度>坡位。
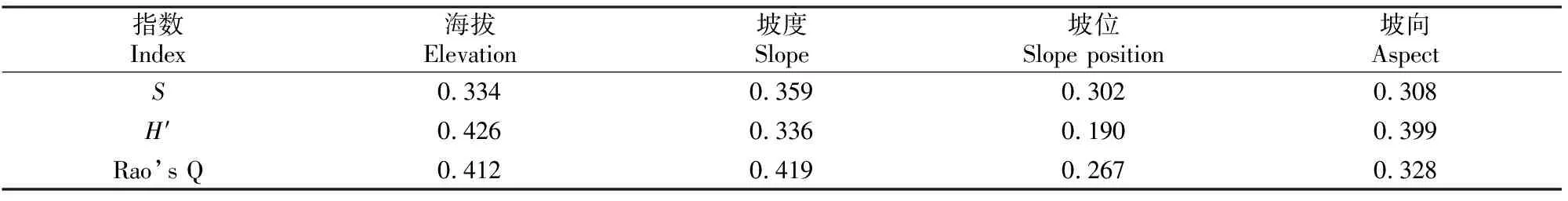
表3 长白山阔叶红松林内典型森林群落的多样性指数与地形因子的灰色关联度分析
3 讨 论
3.1 群落内物种多样性和功能多样性的影响因素
杨桦林、白桦林、臭冷杉红松林和紫椴红松林是最适应长白山阔叶红松林地区气候条件的群落类型,其群落生长发育最完善,最能代表该地区植被的功能,对该地区植物群落功能的影响也最大。杨桦林处于演替早期,阳性先锋树种的多度及重要值较高,在林分中占据着重要地位,生境具有开放性和光照充足等特点,故灌木层和草本层植物能够得到充分的光照,生长良好;白桦林处于演替中期,阳性树种和耐阴树种都能够生长,组成群落乔木层的物种类丰富,因而乔木层物种多样性指数较高;紫椴红松林中由于红松占绝对优势,抑制了乔木层其他植物种的繁殖与生长,因而乔木层植物的种类与数量都比较少,群落结构简单,林下光线较好,故林下植物多样性相对较高,但是由于紫椴红松林样地有野生动物频繁出没,使其草本层植物生物多样性受到的干扰程度较大。
植物不同生活型在群落中的地位对评价群落的动态及生态系统过程重要性有较大影响,本研究结果显示,除白桦林外,其余3种群落不同层次间的Rao’s Q指数均表现为乔木层>灌木层>草本层,白桦林Rao’s Q指数为草本层>乔木层>灌木层。乔木层作为植物群落内的优势层,除受下层灌木和草本的影响外,受到阳光直接照射、温度和降水的影响也比较大,其占据了最优越的水热条件,同时也是群落功能的主要承担者,决定着群落的多样性,而灌木层和草本层则处于次要地位。相比于其他森林群落,紫椴红松林在群落功能方面处于较高的地位,有研究表明,植物演替后期,其功能特性受自身生理学特性的影响更大[20]。对于白桦林而言,由于其处于演替中期,草本层物种较为丰富,种间差异性较大,功能性状数值差异显著,因此草本层Rao’s Q指数较大。
3.2 群落整体物种多样性和功能多样性的影响因素
陆地植物群落物种多样性的影响因素是多种多样的,各种因子所起的作用也有大有小,但某一地域的物种多样性是各种因子综合影响的结果,因此试图用某一种模式概括物种多样性的演替规律是不可能的。本研究结果表明,物种多样性随着演替的进行而呈现先上升后下降的趋势,这与贺金生等[21]总结出的演替梯度上物种多样性的变化规律是一致的。
在本研究的4种典型森林群落中,紫椴红松林(200年)作为阔叶红松林演替阶段的顶级群落,其功能多样性指数最大,其次是演替中期的臭冷杉红松林(130年),最后为演替中期的白桦林(70年)和演替初期的杨桦林(30年),根据各群落功能多样性指数变化趋势,说明功能多样性可能与群落演替阶段有关。在长白山阔叶红松林中,优势植被类型为森林群落,群落内乔木种类的组成及其数量直接影响群落环境,并与群落的功能高低显著相关,而灌木层和草本层处于次要地位。由本研究结果可以初步推测,长白山阔叶红松林群落的 Rao’s Q指数随演替的进行先下降后上升。这与Shannon-Winner指数随着演替的变化趋势恰恰相反。原因在于,Rao’s Q指数会随着物种丰富度的增加而降低,因为群落物种数量增加会导致随机抽取的2个物种的特征差异平均距离减小[13];而Shannon-Winner指数一般情况下会随着物种丰富度的增加而增大[14]。
3.3 地形因子对群落物种多样性的影响
地形因子的作用在于影响物质和能量的再分配,地形是多种环境因子的复合,水分、温度和土壤养分的变化集中体现在地形梯度的变化上,进而反映在植物功能性状的变化上。尤其在小尺度研究领域,这些非生物因子的影响尤其明显[22]。海拔对植物功能性状的影响机理主要是,随着海拔的升高,植物生长发育逐渐受到温度和水分的限制,因而适应性会发生显著变化,尤以叶片对环境变化的反应最为敏感。通常来讲,生活在相对贫瘠的环境中的植物具有较小的比叶面积(SLA)[23],有许多研究发现,叶面积随着海拔的升高而减小,随着年均气温及年均降雨量的降低而减小[24-25]。这可能是由于在高海拔和低温环境下,叶片接受光能、同化二氧化碳的时间缩短,植物因此可通过叶片增加氮含量。同时许多研究表明,无论是单位面积的叶氮含量还是单位质量的叶氮含量,均随海拔的升高而增加[26-28]。
在山地环境中,不同坡位形成的小生境差异较大,上坡位具有太阳辐射和紫外线照射强、土层薄、土壤贫瘠、土壤含水量低、温度较高等特点[29],与中下坡位形成差异,因而表现出坡位对植物叶片氮含量影响较大的结果。坡向的变化也会影响到光照、温度和水分条件,特别是在北半球地区,东坡与西坡、东北坡与东南坡、西北坡与西南坡以及各种不同坡向之间的光照时间和强度不同,其所在地段的温度和水分条件必然也会有一定的差异,这种环境条件的差异必然会影响植物的叶片形态[24]。植物的生长发育不仅取决于自身的生理特性,也受环境条件的影响。对于功能多样性指数而言,以物种性状特征为基础,其综合作用反映群落的整体功能特征。但也有研究结果表明,环境因子对树种不同生长阶段个体的作用有一定的差别,环境因子主要作用于幼树和小树,对大树的解释能力较差[20],演替后期的生境由于植被的缓冲作用,在环境和植被变化方面较为封闭和稳定,各环境因子在空间尺度上的一致性较强,演替早期和演替后期的群落不仅植物组成不同,而且在演替的不同阶段,不同物种的生理和生态特性及其对环境的适应性也有一定的差异。植物群落与其所处环境有着不可分割的联系,生境特征决定着森林群落的种类;同时,森林群落也不断影响着生境的许多特征[30]。紫椴红松林处于演替顶级阶段,在树种与环境因子相互作用的过程中形成了较好的适应关系,且在紫椴红松林中幼树较少,树木种群主要由大径级植株构成,因此植物群落整体的功能特征受环境状况和树种本身功能性状的影响更大[31]。本研究结果表明,海拔对植物群落物种多样性的影响最大,而坡度对群落功能多样性影响最大。
植物群落结构多样性研究的主要是群落中有机体性状值的范围和分布,是植物群落研究的基础。而植物群落功能多样性的研究是在这一基础上发展而来的,由于两者相辅相成的关系,使得其在生态学研究过程中常常同时出现,并逐渐成为生态学研究的重点和热点。因此,功能多样性是解释物种多样性-环境-生态系统功能关系的关键因子。尽管研究人员就“功能多样性比物种数能够更强烈地影响系统功能”这一观点已达成了共识,但事实上,还没有明确的机制来阐释功能多样性与环境和物种多样性的关系,以及解释环境条件是如何影响功能多样性的[32]。多样性对生态系统过程的影响应归因于单个物种及其互相作用的功能特征,而不是物种数。物种多样性是表征群落学的重要指标,在反映植物群落的生境差异、结构类型、演替阶段和稳定性程度等方面均有一定的意义。也有研究发现,与物种多样性相比,功能特征和功能特征多样性与生态系统过程的关系更加紧密,其中尤以功能特征与生态系统过程的关联更密切[1]。
总体而言,以群落为单位对功能多样性指数与环境的关联性进行分析,对认识不同群落对生境的适应性具有重要的理论和实际意义。但实际上,植物群落都是由单一物种形成,并在群落中达成物种共存,包含着复杂的种内和种间关系,而这又会对群落水平的环境关联性起到促进或抑制作用。因此,较物种水平而言,将包括了物种组成和种内、种间关系等综合因素的群落实际物种组成作为一个有机整体, 来理解群落功能多样性-环境的关系具有更为重要的理论意义。
[参考文献]
[1] Díaz S,Cabido M.Vivela diffrence:Plant functional diversity matters to ecosystem processes [J].Trends in Ecology & Evolution,2001,16(11):646-655.
[2] Tilman D,Reich P,Knops J,et al.Diversity and productivity in a long-term grassland experiment [J].Science,2001,294(5543):843-845.
[3] Petchey,Gaston K.Functional diversity (FD),species richness and community composition [J].Ecology Letters,2002,5(3):402-411.
[4] Mason N,Mouillot D,Lee W,et al.Functional richness,functional evenness and functional divergence:The primary components of functional diversity [J].Oikos,2005,111(1):112-118.
[5] 江小雷,张卫国.功能多样性及其研究方法 [J].生态学报,2010,30(10):2766-2773.
Jiang X L,Zhang W G.Functional diversity and its research method [J].Acta Ecologica Sinica,2010,30(10): 2766-2773.(in Chinese)
[6] Rao C R.Diversity and dissimilarity coefficients:An unified approach [J].Theor Popul Biol,1982,21:24-43.
[7] Lavorel S,Grigulis K,McIntyre S,et al.Assessing functional diversity in the field-methodology matters [J].Functional Ecology,2008,22(1):134-147.
[8] Mouillot D,Mason W H N,Dumay O,et al. Functional regularity:A neglected aspect of functional diversity [J].Oecologia,2005,142(3):353-359.
[9] Hooper D U,Vitousek P M.The effects of plant composition and diversity on ecosystem processes [J].Science,1997,277(5330):1302-1305.
[10] Díaz S,Hodgson J G,Thompson K,et al.The plant traits that drive ecosystems:Evidence from three continents [J].Journal of Vegetation Science,2004,15(3):295-304.
[11] Ackerly D,Cornwell W.A trait-based approach to community assembly:Partitioning of species trait values into within-and among community components [J].Ecology Letters,2007,10(2):135-145.
[12] Suding K,Lavorel S,Chapin F,et al.Scaling environmental change through the community-level: A trait-based response-and effect framework for plants [J].Global Change Biology,2008,14(5):1125-1140.
[13] 尧婷婷,孟婷婷,倪 健,等.新疆准噶尔荒漠植物叶片功能性状的进化和环境驱动机制初探 [J].生物多样性,2010,18(2):201-211.
Yao T T,Meng T T,Ni J,et al.Leaf functional trait variation and its relationship with plant phylogenic background and the climate in Xinjiang Junggar Basin,China [J].Biodiversity Science,2010,18(2):201-211.(in Chinese)
[14] 马克平,刘玉明.生物群落多样性的测度方法(Ⅰ) [J].生物多样性,1994,2(4):231-239.
Ma K P,Liu Y M.Measurement of biological community diversity (Ⅰ) [J].Chinese Biodiversity,1994,2(4):231-239.(in Chinese)
[15] 于振良,董立荣.阔叶红松林的研究综述 [J].吉林林学院学报,1996,12(4):235-238.
Yu Z L,Dong L R.Summary of broadleaved Korean pine forest [J].Journal of Jilin Forestry University,1996,12(4):235-238.(in Chinese)
[16] 陈大珂,周晓峰, 祝 宁.天然次生林:结构·功能·动态与经营 [M].哈尔滨:东北林业大学出版社,1994.
Chen D K,Zhou X F,Zhu N.Natural secondary forest: Structure,function,dynamic and management [M].Harbin:Northeast Forestry University Press,1994.(in Chinese)
[17] 张 峰,张金屯.历山自然保护区猪尾沟森林群落植被格局及环境解释 [J].生态学报,2003,23(3):421-427.
Zhang F,Zhang J T.Pattern of forest vegetation and its environmental interpretation in Zhuweigou,Lishan Mountain Nature Reserve [J].Acta Ecologica Sinica,2003,23(3):421-427.(in Chinese)
[18] Magurran A E.Ecological diversity and its measurement [M].Princeton,New Jersey:Princeton University Press,1998.
[19] 宝 乐,刘艳红.东灵山地区不同森林群落叶功能性状比较 [J].生态学报,2009,29(7):3692-3703.
Bao L,Liu Y H.Comparison of leaf functional traits in different forest communities in Mt.Dongling of Beijing [J].Acta Ecologica Sinica,2009,29(7):3692-3702.(in Chinese)
[20] 张春雨,赵秀海,赵亚洲,等.长白山温带森林不同演替阶段群落结构特征 [J].植物生态学报,2009,33(6):1090-1100.
Zhang C Y,Zhao X H,Zhao Y Z,et al.Community structure in different successional stages in north temperate forests of Changbai mountains,China [J].Chinese Journal of Plant Ecology,2009,33(6):1090-1100.(in Chinese)
[21] 贺金生,陈伟烈,江明喜,等.长江三峡地区退化生态系统植物群落物种多样性特征 [J].生态学报,1998,18(4):399-407.
He J S,Chen W L,Jiang M X,et al.Plant species diversity of the degraded ecosystems in the Three Gorges Region [J].Acta Ecologica Sinica,1998,18(4):399-407.(in Chinese)
[22] Loreau M,Naeem P.Biodiversity and ecosystem functioning:Current knowledge and future challenges [J].Science,2001,294(5543):804-808.
[23] Westoby M,Falster D S,Moles A T,et al.Plant ecological strategies:Some leading dimensions of variation between species [J].Annual Review of Ecology and Systematics,2002,33:125-159.
[24] Ackerly D,Knight C,Weiss S,et al.Leaf size,specific leaf area and microhabitat distribution of chaparral woody plants:Contrasting patterns in species level and community level analyses [J].Oecologia,2002,130(3):449-457.
[25] Bragg J G,Westoby M.Leaf size and foraging for light in a sclerophyll woodland [J].Functional Ecology,2002,16(5):633-639.
[26] Friend A D,Woodward F I,Switsur V R.Field measurements of photosynthesis,stomatal conductance,leaf nitrogen and 13 C along altitudinal gradients in Scotland [J].Functional Ecology,1989,3(1):117-122.
[27] Korner C.The nutritional status of plants from high altitudes [J].Oecologia,1989,81(3):379-391.
[28] Craine J M,Lee W G.Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand [J].Oecologia,2003,134(4):471-478.
[29] 祁 建,马克明,张育新.北京东灵山不同坡位辽东栎(Quercusliaotungensis)叶属性的比较 [J].生态学报,2008,28(1):122-128.
Qi J,Ma K M,Zhang Y X.Comparisons on leaf traits ofQuercusliaotungensisKoidz on different slope positions in Dongling Mountain of Beijing [J].Acta Ecologica Sinica,2008,28(1):122-128.(in Chinese)
[30] 戈 峰.现代生态学 [M].北京:科学出版社,2002:210-211.
Ge F.Modern ecology [M].Beijing:Science Press,2002:210-211.(in Chinese)
[31] 张春雨,赵秀海,夏富才.长白山次生林树种空间分布及环境解释 [J].林业科学,2008,44(8):1-8.
Zhang C Y,Zhao X H,Xia F C.Spatial distribution of tree species and environmental interpretations of secondary forest in Changbai Mountains [J].Scientia Silvae Sinicae,2008,44(8):1-8.(in Chinese)
[32] 李伟绮.生境空间对植物功能多样性与生产力关系的影响 [D].兰州:兰州大学,2010.
Li W Q.Effects of available soil space on functional traits-productivity relationship [D].Lanzhou:Lanzhou University,2010.(in Chinese)
