不同寄主植物上麦长管蚜生活史的比较
2014-03-26高素霞黄贤亮刘德广
高素霞,黄贤亮,刘德广 ,陈 辉
(西北农林科技大学 a 旱区作物逆境生物学国家重点实验室,b 林学院,陕西 杨凌 712100)
麦长管蚜Sitobionavenae(Fabrieius)是一种世界范围内广泛分布的害虫,在我国各麦区均有发生,是大多数麦区的优势种[1]。麦长管蚜除了为害小麦外,还为害大麦、燕麦和黑麦,另外还取食禾本科和莎草科的杂草[2]。尽管麦长管蚜能取食田边的很多种杂草,但更喜食禾本科的麦类作物[3]。麦长管蚜先为害麦叶, 然后转到穗部为害, 造成小麦严重减产和品质下降。同时,该虫在取食过程中能传播大麦黄矮病毒(BYDV),并随着蚜虫的迁飞引起小麦黄矮病的流行,造成严重的经济损失。农民长期使用高毒化学杀虫剂防治该虫,使环境日益恶化,蚜虫的抗药性也日益增强,因此应该发展以生态调控为基础的蚜虫绿色防治技术。
不同寄主植物所含次生物质有别,使蚜虫的取食受到影响[4],其生命表参数会发生变化,种群也可能因此发生分化。小麦品系间次生物质如生物碱、单宁、氧肟酸的含量有明显差异[5-6],其中氧肟酸的含量与蚜虫的感染率呈明显负相关[7]。可见,这些次生物质使得抗蚜小麦品种对麦长管蚜有明显的抗生作用,而且其抗生性在蚜虫世代间可能存在累加效应[8]。小麦收割后,麦长管蚜为了繁衍下去,需要寻找替代寄主或野生寄主如田边杂草。国内外有关麦双尾蚜(DiuraphisnoxiaMord.)各种寄主的研究很多[9-10],但对麦长管蚜与其寄主植物之间关系的研究还非常有限,特别是关于它在不同寄主植物上的生命表方面的研究几乎还是空白。本研究通过对不同寄主上麦长管蚜的生活史和生命表参数进行分析,探讨蚜虫在田间经历长期自然选择后,取食经验对其生长发育及繁殖的影响,以及寄主转换后,蚜虫对各种寄主植物的适合度是否发生改变,以探明寄主植物在麦长管蚜种群分化中的作用,从而为发展麦长管蚜生态控制技术奠定基础,为抗蚜育种提供新的材料。
1 材料与方法
1.1 供试虫源
2012-04开始在陕西杨凌周边的小麦、大麦、燕麦上采集各麦长管蚜种群。为了尽可能地采集到不同蚜虫克隆系样品,每头蚜虫均间隔10 m以上采集,将无翅孤雌成蚜带回实验室,在(20±1) ℃及光周期16 h∶8 h(L∶D)条件下分别饲养于小麦、大麦和燕麦上,在每种寄主上饲养2~3代后用于试验。
1.2 供试寄主植物及寄主转换方法
试验所用3种寄主植物分别是小麦(矮抗58)、燕麦(sandle)和大麦(西安91-2)。
3种植物分别栽种于直径10 cm的塑料花盆,以维持不同的种群。试验前,每种植物分别转移至直径6 cm的塑料杯中,每杯1株苗,所有植物都在养虫室同一条件下生长,每周浇水2次。寄主转换试验中,小麦上麦长管蚜种群分别被转接到小麦、大麦和燕麦上,以小麦作为对照,共计3个处理,每处理设30个重复;燕麦和大麦上麦长管蚜种群也同样地在上述3种寄主上转换并进行生测。
1.3 生命表参数调查方法
试验前,从各种群取刚蜕皮的无翅成蚜,单头接于1~2叶期的幼苗上(大约7 d),产蚜4 h后取走成蚜,单株只留1头刚出生的若蚜,然后罩于用透明塑料胶片制成的圆柱形(直径6 cm,高15 cm)笼罩中,顶部用尼龙纱网封口。每天早晚2次调查若蚜蜕皮和存活情况,并移去蚜蜕。蜕皮4次后进入成蚜阶段,每天1次定时调查并记录产蚜数量,然后用毛笔刷掉所有初产若蚜,保留成蚜,直至所有成蚜死亡时结束观察。试验均在温度(20±1) ℃、相对湿度(65±5)%、光周期为16 h∶8 h(L∶D)的恒温气候箱中进行,所用植物每7 d左右更换1次。
1.4 数据分析
用以下公式计算麦长管蚜世代平均周期(T)、净增殖率(R0)、内禀增长率(rm)和周限增长率(λ) 及种群倍增时间(DT)[11]:T=∑(lxmxx)/∑(lxmx),R0=∑lxmx,rm=ln(R0)/T,λ=erm,DT=ln(2)/rm。式中:x为处理时间,lx为第x天时雌虫存活率,mx为第x天时平均每头雌虫产下的后代数,即繁殖力。
数据用“平均值±标准差”表示。处理均应用 SAS 9.1 统计分析软件进行,用PROC GLM步骤进行方差分析,然后用Tukey检验进行处理间差异显著性分析。生命表参数参照Maia等[12]的方法进行分析和比较。
2 结果与分析
2.1 不同寄主植物麦长管蚜种群若虫发育时间和成虫繁殖参数的比较
由表1可以看出,不同寄主植物上麦长管蚜若虫发育历期存在明显差异。当原始寄主和替代寄主相同时,小麦和燕麦上麦长管蚜种群若虫发育历期较短(约8.5 d),大麦上麦长管蚜种群若虫发育历期较长(10.1 d)。当小麦上麦长管蚜种群转到大麦后,发育历期显著延长;大麦上麦长管蚜种群转接到小麦后,若虫发育历期则显著缩短;而燕麦上麦长管蚜种群在转移到大麦和小麦后,若虫发育历期没有显著差异。不同寄主上麦长管蚜种群成虫寿命也存在明显差异。当原始寄主和替代寄主相同时,大麦上麦长管蚜种群成虫寿命最长(35.6 d),小麦和燕麦上麦长管蚜种群成虫寿命明显缩短。大麦上麦长管蚜种群转到小麦和燕麦之后,其成虫寿命缩短11 d以上;小麦上麦长管蚜种群转到燕麦上之后,其成虫寿命也明显缩短,而转移到大麦之后则无显著变化;燕麦上麦长管蚜转到大麦和小麦上之后,其成虫寿命显著延长。总体来看,原始寄主为燕麦时,其上麦长管蚜种群成虫的繁殖力较强,小麦上麦长管蚜种群居中,大麦上麦长管蚜种群繁殖力较弱。小麦上麦长管蚜种群转到燕麦上之后,其繁殖力显著下降;燕麦上麦长管蚜种群转到大麦上之后,其繁殖力也显著下降;而大麦上麦长管蚜种群转换寄主后其繁殖力无显著变化。除燕麦上麦长管蚜种群在转换寄主之后其成虫繁殖期没有显著变化外,大麦和小麦上麦长管蚜种群成虫繁殖期在转换寄主前后变化显著。
由表1还可以看出,不同寄主植物上麦长管蚜种群的日均产蚜量也存在明显差异。当原始寄主和替代寄主相同时,小麦和燕麦上麦长管蚜种群的日均产蚜量高于大麦。转换寄主后,大麦上麦长管蚜种群日均产蚜量增加,而小麦上麦长管蚜种群基本不变;燕麦上麦长管蚜种群转到大麦上之后日均产蚜量显著下降,转到小麦上之后无显著变化。
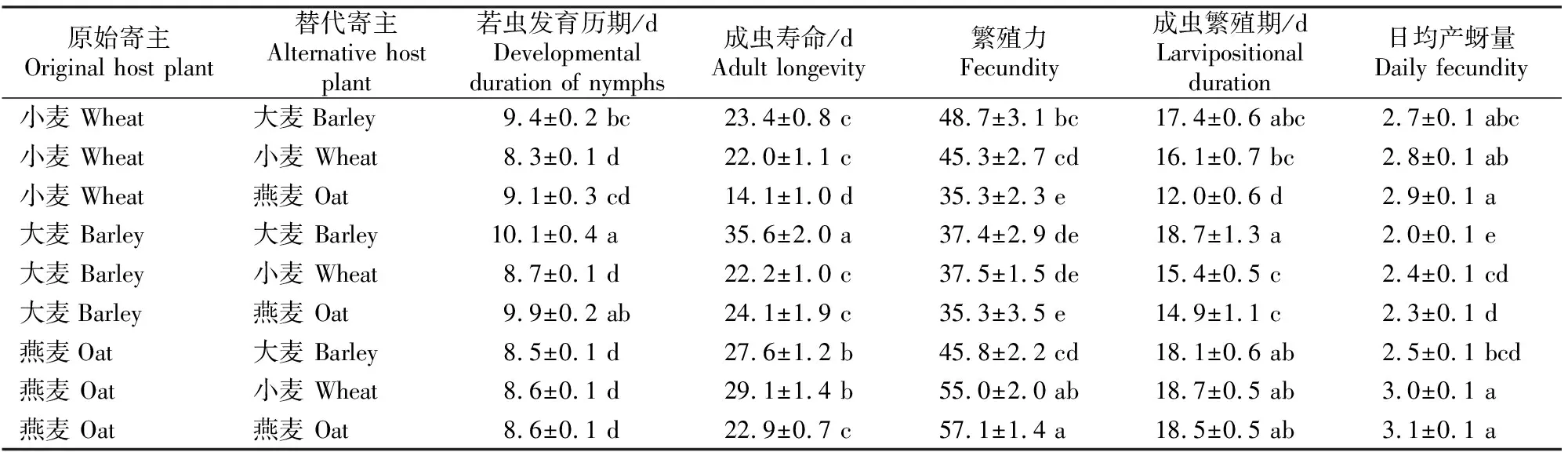
表1 不同寄主植物上麦长管蚜发育历期和繁殖参数的比较
2.2 不同寄主植物上麦长管蚜种群生命表参数的比较
从表2可以看出,当原始寄主和替代寄主相同时,燕麦上麦长管蚜种群的内禀增长率、净增殖率和周限增长率均最大,小麦上麦长管蚜种群居中,大麦上麦长管蚜种群最小。小麦和燕麦上麦长管蚜种群转换寄主之后,其内禀增长率(rm)趋于下降;大麦上麦长管蚜种群转移寄主后,其rm显著上升。麦长管蚜各种群的种群倍增时间(DT)与内禀增长率(rm)的变化趋势刚好相反。小麦上麦长管蚜种群转移到燕麦上之后,其净增殖率(R0)显著下降;转移到大麦上之后其R0无显著变化。燕麦上麦长管蚜种群转到大麦上之后,其R0也显著下降;而转移到小麦上之后则无显著变化。大麦上麦长管蚜种群转到小麦、燕麦上后,其R0有所上升,但差异不显著。当原始寄主和替代寄主相同时,大麦上麦长管蚜种群的世代周期(T)最长,小麦和燕麦上麦长管蚜种群的T较短。小麦上麦长管蚜种群转到大麦上后,其T延长;大麦上麦长管蚜种群转换寄主后,其T显著缩短;而燕麦上麦长管蚜种群转换寄主后,其T无显著变化。

表2 不同寄主植物上麦长管蚜种群的生命表参数
不同寄主植物上麦长管蚜种群成虫存活率和繁殖力的变化见图1。
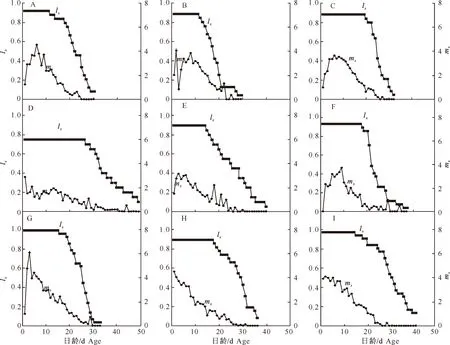
图1 不同寄主植物上麦长管蚜各日龄成虫存活率和繁殖力的比较
图1显示,当原始寄主和替代寄主相同时,燕麦上的麦长管蚜种群前期成虫存活率最高(1,图1G),小麦上麦长管蚜种群居中(图1A),大麦上麦长管蚜种群的存活率最低(0.75,图1D)。各个寄主植物上麦长管蚜种群成虫开始出现死亡的时间也不同,其中小麦上麦长管蚜种群12 d时存活率已经开始下降,并且下降迅速;燕麦上麦长管蚜种群次之,16 d时存活率开始下降;大麦上麦长管蚜种群27 d时才有成虫死亡,存活率下降幅度较小。当原始寄主和替代寄主相同时,麦长管蚜特定时间的繁殖力也存在很大差异,小麦和燕麦种群成虫的繁殖力总体呈先上升后下降的趋势,繁殖高峰分别出现在羽化后第6天和第3天(图1A,图1G);而大麦上麦长管蚜种群成虫的繁殖力较低(图1D),且总体呈降低趋势,繁殖期较长。小麦上麦长管蚜种群转到大麦上之后,前期存活率变化不大(图1C),但在燕麦上成虫存活率下降趋势比较明显(图1B)。转换寄主后,大麦上麦长管蚜种群在燕麦(图1E)和小麦(图1F)上存活率明显升高,成虫出现死亡的时间也有所提前。燕麦上麦长管蚜种群转到小麦上后,前期存活率没有变化(图1I);但转到大麦上之后,前期存活率明显下降(图1H)。小麦上麦长管蚜种群转移到燕麦上后繁殖力高峰期提前,繁殖力略有提高;而转到大麦上之后繁殖力峰值有所降低。大麦上麦长管蚜种群转换寄主后,在替代寄主上繁殖力呈先升后降的趋势,而燕麦上麦长管蚜种群转换寄主后其繁殖力呈下降趋势。
3 结论与讨论
本研究结果表明,麦长管蚜种群在不同寄主上的生长发育和繁殖参数存在明显差异。当原始寄主和替代寄主相同时,与大麦上麦长管蚜种群相比,小麦和燕麦上麦长管蚜种群若虫发育历期较短,繁殖力和日均产蚜量较高,成虫寿命较短;小麦上麦长管蚜种群的成虫繁殖期缩短,但燕麦上麦长管蚜种群变化不大。当原始寄主和替代寄主相同时,燕麦上麦长管蚜种群的内禀增长率、净增殖率和周限增长率均最大,小麦上麦长管蚜种群居中,大麦上麦长管蚜种群最小;燕麦上麦长管蚜种群前期存活率最高,小麦上麦长管蚜种群居中,大麦上麦长管蚜种群最低;燕麦和小麦上麦长管蚜种群存活率均下降较早且快,大麦上麦长管蚜种群下降较晚且慢;小麦和燕麦上麦长管蚜种群成虫在前10 d的繁殖力均较高,而大麦上麦长管蚜种群则一直维持在较低的水平。总的来看,麦长管蚜对小麦和燕麦的适应性要高于大麦。笔者在多年的田间调查中也发现,在麦长管蚜发生高峰期,小麦上蚜虫的虫口密度明显高于大麦。本研究结果显示,转换寄主后,麦长管蚜种群成虫存活率和繁殖力随日龄的增加而产生了明显的波动性变化。这些现象说明麦长管蚜在不同寄主植物上出现了一定的分化,其种群分化的一个关键原因在于寄主植物之间的次生代谢物质有明显区别,对蚜虫产生的自然选择压力也会不同[13-15]。不同寄主上麦长管蚜的分化程度还有待于进一步验证。
蚜虫在不同寄主植物上发生分化的现象并不少见[14]。例如,豌豆蚜已分化出适应不同寄主植物的专化型[16-17]。也有研究表明,来自同一寄主不同克隆系的麦长管蚜 (S.avenae)对小麦和鸭茅草(Dactylisglomerata)的选择性存在显著差异,这些蚜虫克隆系往往在原始寄主上比转换寄主上生活得要好,且寄主植物转换顺序对小麦上蚜虫克隆系的影响较其对鸭茅草上蚜虫克隆系的影响大,因此小麦上蚜虫克隆系比鸭茅草上蚜虫克隆系更具有专化性[3]。本研究中,小麦上麦长管蚜种群转到大麦上后,其若虫发育历期和世代周期延长,但种群周限增长率下降;转到燕麦上后,其若虫发育历期延长,而且成虫寿命和繁殖期都缩短。燕麦上麦长管蚜种群转到大麦上后,成虫寿命延长,但繁殖力和种群周限增长率下降;转到小麦上后,若虫发育历期、成虫繁殖期及种群生命表参数均无显著变化。大麦上麦长管蚜种群转到小麦或燕麦后,其成虫寿命和成虫繁殖期及若虫发育历期缩短,种群周限增长率提高。可见,相对于燕麦或大麦种群,小麦上麦长管蚜种群似乎具有较高的寄主专化性。这与Dean[18]的研究结果“麦长管蚜对大麦的喜好性强于燕麦和小麦,在大麦上有较高的存活率和较短的世代周期”不同。原因可能在于Dean仅仅采用来自大麦的惟一麦长管蚜克隆系,并将它转接到大麦、小麦和燕麦上进行测试所致。另外也有研究表明,采自同一寄主植物和同一位置的麦长管蚜在对小麦和鸭茅草选择时,则更偏向选择小麦[19]。由于本试验中所用的寄主植物种类有限,还不能对麦长管蚜是否具有寄主专化性下定论或对其普食和专化程度作出确切评估。因此,还需要利用更多的寄主特别是野生寄主来进行测试。
[参考文献]
[1] 胡想顺,赵惠燕,Heimbach Udo,等.3个新引进小麦品种对麦长管蚜抗性的初步研究 [J].西北植物学报,2004,24(7):1221-1226.
Hu X S,Zhao H Y,Heimbach Udo,et al.Study on cereal aphid resistance on three winter wheat cultivars introduced into China [J].Acta Bot Boreal-Occident Sin,2004,24(7):1221-1226.(in Chinese)
[2] 仵均祥.农业昆虫学 [M].北京:中国农业出版社,2002.
Wu J X.Agricultural entomology [M].Beijing:China Agriculture Press,2002.(in Chinese)
[3] De Barro P J,Sherratt T N,David O,et al.An investigation of the differential performance of clones of the aphidS.avenaeon two host species [J].Oecologia,1995,104:379-385.
[4] Tsai J H,Wang J J.Effects of host plants on biology and life table parameters ofAphisspiraecola(Homoptera:Aphididae) [J].Environ Entomol,2001,30(1):45-50.
[5] 陈巨莲,倪汉祥,孙京瑞.主要次生物质对麦蚜的抗性阈值及交互作用 [J].植物保护学报,2002,29(1):7-12.
Chen J L,Ni H X,Sun J R.The resistance threshold and interactions of several plant secondary metabolites to wheat aphids [J].Acta Phytophylacica Sinica,2002,29(1):7-12.(in Chinese)
[6] Eleftherianos I,Vamvatsikos P,Ward P D,et al.Changes in the levels of plant total phenols and free amino acids induced by two cereal aphids and effects on aphid fecundity [J].Journal of Applied Entomology,2006,130:15-19.
[7] Gianoli E,Papp M,Niemeyer H M.Costs and benefits of hydr-oxamic acids-related resistance in winter wheat against the bird cherry-oat aphid,RhopalosiphumpadiL. [J].Annals of Applied Biology,1996,129(1):83-90.
[8] 郭 萧,李克斌,尹 姣,等.不同小麦品种(系)对麦长管蚜生命参数的影响 [J].中国农业科学,2010,43(10):2056-2063.
Guo X,Li K B,Yin J,et al.Effects of wheat varieties on population parameters ofMacrosiphumavenae(Fabricius) [J].Scientia Agricultura Sinica,2010,43(10):2056-2063.(in Chinese)
[9] Clement S L,Lester D G,Wilson A,et al.Behavior performance ofDiuraphisnoxia(Homoptera:Aphididae) on fungal endophyte-infected and uninfected perennial ryegrass [J].Econ Entomol,1992,85(2):583-588.
[10] Schotzko D J,Smith C M.Effects of preconditioning host plants on population development of Russian wheat aphids(Homoptera:Aphididae) [J].Journal of Economic Entomology,1991,84(3):1083-1087.
[11] 徐汝梅.昆虫种群生态学 [M].北京:北京师范大学出版社,1987:152-160.
Xu R M.Insect population ecology [M].Beijing: Beijing Normal University Press,1987:152-160.(in Chinese)
[12] Maia A H N,Luiz A J B,Campanhola C. Statistical inference on associated fertility life parameters using jackknife technique:Computational aspects [J].Journal of Economic Entomology,2000,93(2):511-518.
[13] Futuyma D J,Philippi T E.Genetics of the aphid Crypomyzus,with a preliminary analysis of the inheritance of host preference,reproductive performance and host-alternation [J].Entomol Exp Appl,1987,57:65-76.
[14] Via S.Ecological genetics and host adaptation in herbivorous insects:The experimental study of evolution in natural and agricultural systems [J].Annu Rev Entomol,1990,35:421-446.
[15] Bernays E A.Effects of experience on host-plant selection [M]//Chemical ecology of insects 2.Amherst,US:Springer,1995:47-64.
[16] Frantz A,Plantegenest M,Simon J C.Host races of the pea aphidAcyrthosiphonpisumdiffer in male wing phenotypes [J].Bulletin of Entomological Research,2009,100:59-66.
[17] McLean A H C,van Asch M,Ferrari J,et al.Effects of bacterial secondary symbionts on host plant use in pea aphids [J].Proceedings of the Royal Society B:Biological Sciences,2010,278:760-766.
[18] Dean G J W.Bionomics of aphids reared on cereals and some Gramineae [J].Annals of Applied Biology,1973,73:127-135.
[19] Lushai G,Sherratt T N,David O,et al.Host selection by winged summer females of the aphidSitobionavenae[J].Entomologia Experimentalis et Applicata,1997,85:199-209.
