昌邑海洋生态特别保护区潮间带大型底栖动物时空分布特征
2014-03-24张守本刘一霆刘旭东
冷 宇,张守本,刘一霆,赵 升,刘旭东
1.山东省海洋生态环境与防灾减灾重点实验室,山东 青岛 266033
2.国家海洋局北海环境监测中心,山东 青岛 266033
3.山东省潍坊市海洋环境监测中心站,山东 潍坊 261041
昌邑海洋生态特别保护区于2007年10月底获得国家海洋局批准建立,该保护区坐落于山东省昌邑市北部堤河以东、海岸线以下的滩涂上,总面积为2 929.28 hm2,主要保护以柽柳为主的多种滨海湿地生态系统和各种海洋生物, 是中国惟一以柽柳为主要保护对象的国家级海洋特别保护区,区内有柽柳林、滩涂湿地、浅海等多种生态类型。
潮间带是世界滩涂湿地生态系统的一个重要类型,处于陆海过渡地带,由于人类的频繁活动以及海陆理化因子的交替作用,潮间带成为海洋中最敏感的生态系统之一,同时又是人类活动和干扰最为严重的区域。大型底栖动物是潮间带生态系统的重要组成部分,在海洋生态系统的物质循环和能量利用中均具有重要的作用[1]。研究大型底栖动物对了解环境状况,保护生态资源等具有重要意义,因此潮间带大型底栖动物生态学的研究一直备受国内外学者的关注[2-8]。
近年来,人为活动对生态环境影响的日益加剧,直接导致了潮间带大型底栖动物群落结构的变化。因此,及时查清潮间带不同岸相水域的大型底栖动物现状,为保护、开发和利用潮间带海洋生物资源提供科学依据就显得极为重要[9-10]。
对于昌邑海洋生态特别保护区的生态研究,迄今只有植物群落生态分布规律与土壤理化性质的研究[11],该区尚未见系统的底栖动物群落时空变化情况的研究报道。因此笔者在保护区内进行潮间带大型底栖动物调查、分析和研究,旨在掌握此区域潮间带动物种类组成、分布现状、季节变化等基础资料,揭示大型底栖动物群落时空分布的特点,同时为该地区生态系统保护研究提供基础资料,对海岸环境资源的可持续利用以及海洋环境保护和管理提供相关理论依据。
1 实验部分
1.1研究地点及采样方法
调查时间为2011年的3、6、8、10、12月,选择保护区内有代表性的1个断面(LC02),按高、中、低3个潮区设立7个取样站,分别为高潮带1个(LC02-H),中潮带4个(LC02-M1、LC02-M2、LC02-M3、LC02-M4),低潮带2个(LC02-L1、LC02-L2)。定量采样用25 cm×25 cm×30 cm的取样框在每个站位随机取样2次,所采泥样用孔径为0.5 mm的分样筛淘洗,再将获取到的大型底栖动物标本用5%福尔马林溶液固定后带回实验室鉴定分析。室内分析鉴定按《海洋调查规范》(GB 12763.6—2007)中规定的方法进行。研究地点如图1所示。

图1 调查站位设置示意图
1.2数据处理及分析
1.2.1生物多样性指数分析
采用描述生物群落结构的3个参数(种类多样性H′、种类均匀度指数J和种类丰富度指数M),对大型底栖动物群落结构进行分析[12],计算公式分别为
Shannon-Wiener多样性指数
H′=-∑Pilog2Pi
Pielou均匀度指数
J=H′/log2S
Margalef丰富度指数
M=(S-1)/lnN
式中:S为总物种数,Pi为种i的个体数占总个体数的比例,N为全部物种的个体数。
物种优势度计算公式为
Y=(ni/N)×fi
式中:ni为第i种的个体数,fi为该种在各站位出现的频率,当Y>0.02时,认为该种是调查海域的优势种。
1.2.2聚类分析
采用PRIMER 5软件根据物种生物密度平方根转换计算Bray-Curtis相似性系数矩阵,用软件包中等级聚类分析法(CLUSRER)分析群落结构。
2 结果与分析
2.1物种组成及分布
共采集到潮间带动物55种,隶属于8个动物门,9纲,22目,41科,50属。其中,以多毛类最多,为21种,占潮间带动物种类组成的38.18%;软体动物次之,共17种,占潮间带动物种类组成的30.91%;甲壳类出现11种,占种类组成的20.00%;腔肠动物出现2种,占3.64%;纽形、螠虫、腕足和鱼类各1种,分别占1.82%。潮间带动物密度占优势的种类为四角蛤蜊、彩虹明樱蛤、长竹蛏、托氏虫昌螺、秀丽织纹螺、泥螺、鸭嘴海豆芽、日本美人虾、宽身大眼蟹、长趾股窗蟹。各月潮间带动物种类组成详见表1。
从时间分布来看,3月份出现的种类最多,为36种;12月份出现的种类最少,仅10种。动物种类垂直分布以高潮带出现的种类最少,仅3种,中潮带和低潮带种类较丰富,均为43种。5个月份各潮带出现的种类数见图2。
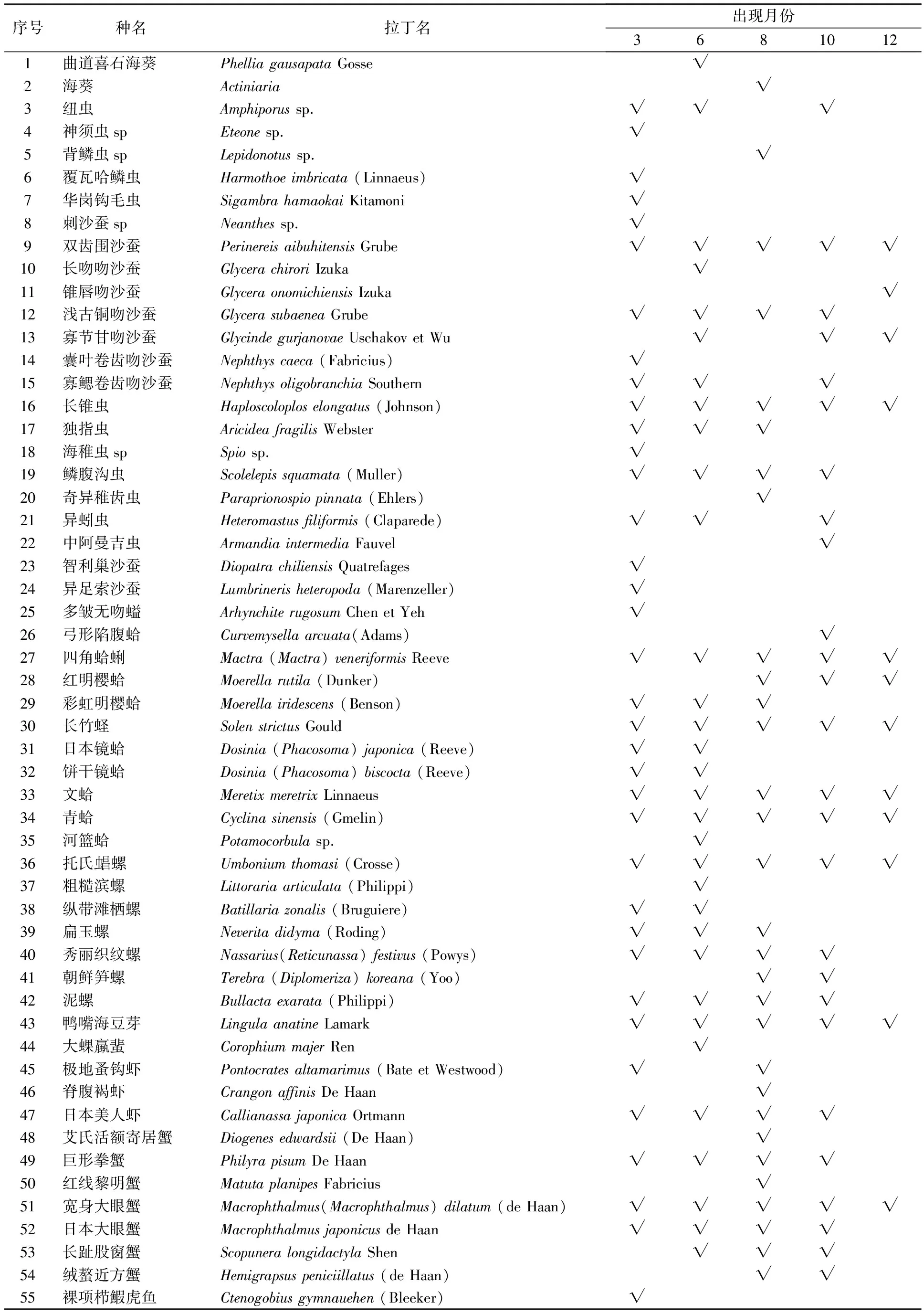
表1 调查发现的大型底栖动物

图2 5次调查中不同潮带动物种类分布情况
2.2生物量和生物密度
2.2.1生物量分布
潮间带动物年均生物量为113.1 g/m2,以6、10月较高,分别为183.9、169.0 g/m2,8、12月较低,分别为61.7、40.8 g/m2。生物量垂直分布呈现自高潮带向低潮带逐渐增加的趋势。生物量不同月份间(F4,8=2.99,P=0.08)差异不显著,在不同潮带间(F2,8=12.75,P=0.003)差异极显著。高潮带年均生物量为8.6 g/m2,其生物量构成为蟹类,3、12月未出现生物个体,6、10月的生物量高于8月。中潮带年均生物量为87.1 g/m2,其生物量构成主要为泥螺、托氏虫昌螺等腹足类,12月未出现生物个体,以6、10月的生物量较高。低潮带年均生物量为95.8 g/m2,其生物量构成主要为托氏虫昌螺、四角蛤蜊、彩虹明樱蛤、长竹蛏、鸭嘴海豆芽、日本美人虾和宽身大眼蟹等,以6、10月的生物量较高,8、12月较低。各潮带不同月份潮间带动物量分布状况见图3。

图3 5次调查中大型底栖动物生物量的季节变化
2.2.2生物密度分布
潮间带动物年均生物密度为366.5 ind./m2,以6、10月较高,分别为528.0、522.4 ind./m2,3、12月较低,分别为260.7、70.7ind./m2。生物密度垂直分布呈现自高潮带向低潮带逐渐增加的趋势。生物密度不同月份间(F4,8=3.54,P=0.06)差异不显著,在不同潮带间(F2,8=13.20,P=0.003)差异极显著。高潮带年均生物密度为76.8 ind./m2,其生物密度构成主要为长趾股窗蟹,偶见双齿围沙蚕,3、12月未出现生物个体,6、8月的生物密度较高。中潮带年均生物密度为322.8 ind./m2,其生物密度构成主要为双齿围沙蚕、浅古铜吻沙蚕、托氏虫昌螺、彩虹明樱蛤、长竹蛏等,12月未出现生物个体,以10月的生物密度较高。低潮带年均生物密度为399.6 ind./m2,其生物密度构成主要为浅古铜吻沙蚕、托氏虫昌螺、秀丽织纹螺、彩虹明樱蛤、四角蛤蜊、长竹蛏、鸭嘴海豆芽和日本美人虾等,以6、8、10月的生物密度较高,3、12月较低。各潮区不同月份潮间带生物密度分布状况见图4。

图4 5次调查中大型底栖动物生物密度的变化
2.3生物多样性
各潮带间不同月份的M指数、J指数以及H′指数值见表2。
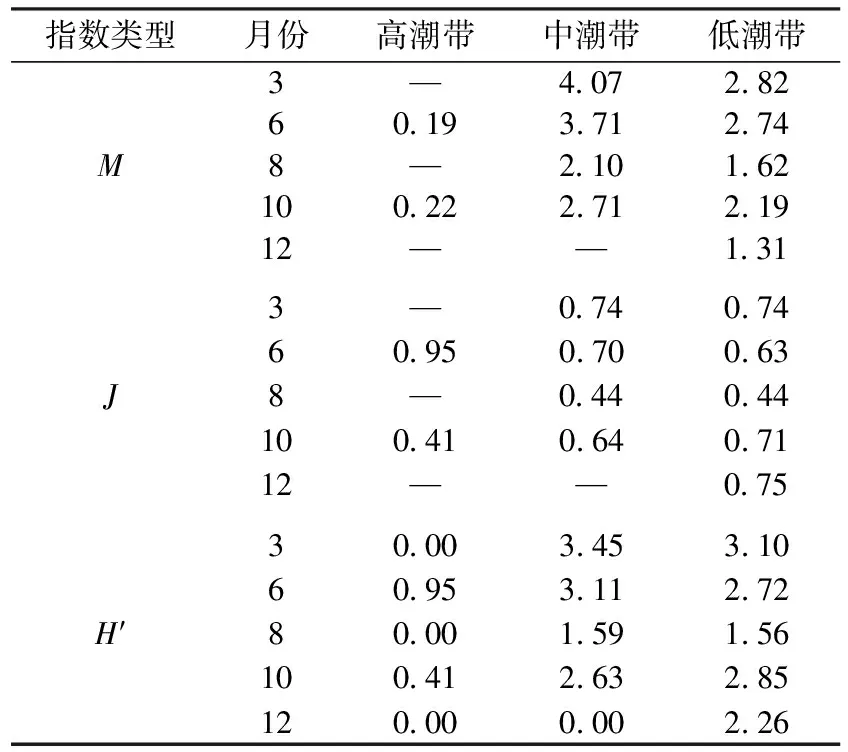
表2 各站位大型底栖动物群落不同月份的生物多样性
对这3个多样性指数进行潮带间与月份间的无重复双因素方差分析,结果为M指数在不同月份间(F4,8=9.05,P=0.03)差异显著,在不同潮带间(F2,8=49.93,P=0.001)差异极显著;J在不同月份间(F4,4=1.51,P=0.35)以及不同潮带间(F2,4=0.003,P=0.997)差异均不显著;H′在不同月份间(F4,8=2.74,P=0.11)差异不显著,在不同潮带间(F2,8=13.70,P=0.003)差异极显著。
2.4群落聚类分析
由群落聚类分析结果(图5)可见,以40%的相似性来看,高潮带动物单独聚为一支,12月低潮带动物(高、中潮带均未采集到动物)单独聚为一支,3、6、8、10月的中、低潮带相似性较高,聚为一支。One-way ANOSIM检验表明各群落间差异极为显著(R=0.93,P=0.001)。

图5 群落结构聚类分析
3 讨论
3.1潮间带大型底栖动物的空间分布
潮位对大型底栖动物群落的物种数、密度和生物量有重要的影响,有研究认为从高潮带到低潮带,物种数、密度和生物量会不断增高[13],较低潮带的物种数和密度较大[14]。也有研究认为,中潮带物种数和密度最高,而低潮带的生物量最大[15-16]。
笔者对潮间带调查研究发现,生物种类的垂直分布为高潮带出现的种类最少,中潮带和低潮带种类较丰富,生物量以及生物密度的垂直分布呈现自高潮带向低潮带逐渐增加的趋势。生物密度及生物量在潮带间差异均极显著,分析发现:四角蛤蜊自中潮带中区开始呈零星分布,高密度区出现在中潮带下区和低潮带上区,是生物量垂直变化的主要原因;栖息于中、低潮带的经济性滩面动物托氏虫昌螺是生物密度垂直变化的主要原因,该种自中潮带中区开始呈零星分布,高密度区出现在中潮带下区和低潮带上区。大个体的托氏虫昌螺多分布于潮位较高的区域,而个体较小的则出现于低潮带,随潮带的降低,其个体逐渐变小,而密度逐渐增高。
无重复双因素方差分析显示,M指数和H′指数在不同潮带间差异均显著,J指数差异不显著。这与调查区域的优势种在高潮带仅出现长趾股窗蟹有关,也因此导致了中、低潮带的H′指数值基本一致,而高潮带出现生物较少,多样性的差异比较明显。
3.2潮间带大型底栖动物的时间分布
无重复双因素方差分析显示,调查区域的生物密度、生物量在不同月份间的差异均不显著,这与以往对潮间带大型底栖动物的季节变化的研究结果相同[17-19]。说明季节的温度变化对各样点的上述指标影响不大。调查区12月开始进入冰冻期,冬季调查时高潮带和中潮带上区的底温已达-4℃,此区域内极少有生物分布,而因其他季节在此区域内栖息的生物种类较少,密度较低,因此冬季海冰的影响未造成生物资源量的明显变化。此调查断面中潮带下区和低潮带,冬季被海冰所覆盖,冰层下方的底泥温度多为0℃,其中较大个体的虾、蟹类和多毛类仍然能够生存(见表1),只是活动迟缓,资源量降低。M指数差异显著,主要与12月气温较低,仅低潮带出现生物,生物种类较少有关。
3.3群落聚类分析
聚类分析方法在群落生态学的研究中被广泛采用,在评价群落结构的相似性上具有重要作用[20]。该研究中, 调查潮间带大型底栖动物群落的聚类分析表明,其群落结构主要与潮汐及季节有关,高潮带首先聚为一支,与中、低潮带的相似性不足10%,12月的群落单独聚为一支,3、6、8、10月的中、低潮带物种组成相似性程度较高(Bra-Curtis 相似性系数超过40%)。聚类结果充分说明了潮汐和季节是影响大型底栖动物群落架构组成的2个重要因素,其中潮汐是影响最大的因素。
3.4保护建议
合理开采滩涂经济生物,对减少生物空间竞争压力,保护生物栖息环境具有促进作用。泥螺和托氏虫昌螺是调查区内2种主要的经济性腹足类动物,它们生活习性相似,空间分布上有重叠现象。泥螺为当地引入养殖种类,生活在中潮带,小个体泥螺栖息于中潮带上区,大个体泥螺分布于中潮带中区,中潮带下区偶然出现。每年春夏季由人工采集收获,苗种资源依靠自然繁殖和人工撒播补苗2种方式。托氏虫昌螺也分布于中潮带,自中潮带上区至下区,生物个体逐渐变小,数量逐渐增加,密集区位于中潮带下区,最高密度达 150 ind./m2。每年夏季由机械拖动耙网采集,作为池塘虾、蟹养殖的饵料,苗种来源于自然繁殖。
2种生物在中潮带中区出现混合栖息现象,且同为滩面生物的优势种,生物密度以托氏虫昌螺较高,生物量则以泥螺较高。该研究发现,尽管存在人为干扰及生物引入现象,调查区域托氏虫昌螺仍是中、下潮带的优势种,泥螺尚未形成大规模入侵现象;而与其相距约30 km的小清河口滩涂,经调查发现,泥螺已经成为中潮带滩面生物的绝对优势种,托氏虫昌螺的资源量明显衰退。相似的栖息环境造成了不同的生态结果,可能是保护区内的调查滩涂不适宜泥螺的幼体生长,同时人为补苗量相对较少,限制了泥螺的过度增殖。这一现象需要今后进一步的连续跟踪监测和深入研究,这项研究对合理开发调查区域的生物资源、保护生态环境具有重要意义。
参考文献:
[1]赵永强,曾江宁,高爱根,等.椒江口滩涂大型底栖动物群落格局与多样性[J].生物多样性,2009,17(3):103-112.
[2]Taft O W, Haig S M. The value of agricultural wetlands as invertebrate resources for wintering shorebirds[J]. Agriculture, Ecosystems and Environment,2005,110(3):249-256.
[3]王宝强,薛俊增,庄骅,等.洋山港潮间带大型底栖动物群落结构及多样性[J].生态学报,2011,31(20):5 865-5 874.
[4]侯森林,余晓韵,鲁长虎.盐城自然保护区射阳河口潮间带大型底栖动物空间分布与季节变化[J].生态学杂志,2011,30(2):297-303.
[5]Fujii T. Spatial patterns of benthic macrofauna in relation to environmental variables in an intertidal habitat in the Humber estuary, UK: Developing a tool for estuarine shoreline management[J]. Estuarine, Coastal and Shelf Science, 2007,75(1):101-119.
[6]Feder H M, Jewett S C, Blanchard A L. Southeastern Chukchi Sea (Alaska) macrobenthos[J]. Polar Biology,2007,30(3):261-275.
[7]胡知渊,李欢欢,鲍毅新,等.灵昆岛围垦区内外滩涂大型底栖动物生物多样性[J].生态学报,2008,28(4):1 498-1 507.
[8]周晓,葛振鸣,施文彧,等.长江口新生湿地大型底
栖动物群落时空变化格局[J].生态学杂志,2007,26(3):372-377.
[9]张宝琳,王洪法,李宝泉,等.胶州湾辛岛潮间带大型底栖动物生态学调查[J].海洋科学,2007,31(1):60-64.
[10]龙华,余骏,周燕.大型底栖动物污染指数在乐清湾潮间带环境质量评价中的应用[J].海洋学研究,2008,26(4):97-104.
[11]汤爱坤,刘汝海,许廖奇,等.昌邑海洋生态特别保护区土壤养分的空间异质性与植物群落的分布[J].水土保持通报,2011,31(3):88-93.
[12]马克平,刘玉明.生物群落多样性的测度方法I:α多样性的测定方法(下)[J].生物多样性,1994,2(4):231-239.
[13]Edgar G J, Barrett N S. Benthic macrofauna in Tasmanian estuaries: Scales of distribution and relationships with environmental variables[J]. Journal of Experimental Marine Biology and Ecology, 2002,270(1): 1-24.
[14]Honkoop P J C, Pearson G B, Lavaleye M S S, et al. Spatialvariation of the intertidal sediments and macrozoo-benthic assemblages along Eighty-mile Beach, Northwestern Aus tralia[J]. Journal of Sea Research,2006,55(4):278-291.
[15]Spruzen F L, Richardson A M M, Woehler E J. Spatial variation of intertidal macroinvertebrates and environmental variables in Robbins Passage wetland, NW Tasmania[J].Hydrobiologia, 2008, 598(1): 325-342.
[16]吕永林,张永普,李摇凯,等.浙江洞头大竹屿岛潮间带大型底栖动物多样性[J].生态学杂志,2011,30(4):707-716.
[17]鲍毅新,胡知渊,李欢欢,等.灵昆东滩围垦区内外大型底栖动物季节变化和功能群的比较[J].动物学报,2008,54(3):416-427.
[18]葛宝明,鲍毅新,郑祥,等.灵昆岛潮间带大型底栖动物群落结构与生态位分析[J].生态学报,2005,25(11):3 037-3 043.
[19]寿鹿,廖一波,徐晓群,等. 福清核电站邻近潮间带大型底栖动物数量分布与群落结构[J].海洋学研究,2009,27(2):42-50.
[20]马藏允,刘摇海,王惠卿,等.底栖动物群落结构变化多元变量统计分析[J].中国环境科学,1997,17(4):297-300.
