辽东栎冠层光合生理特性的空间异质性
2014-03-22胡耀升么旭阳刘艳红
胡耀升, 么旭阳, 刘艳红
(北京林业大学 森林培育与保护省部共建教育部重点实验室,北京 100083)
植物光合作用是其有机物质产生并积累的根本,直接影响着生态系统物质循环和水热平衡,一直是植物生理学、生理生态学中的热点问题,也是全球其他物质循环的最重要枢纽[1-3]。而冠层作为林木与外界环境因子相互作用最直接的部分,同时也是光合作用的主要载体,由于林冠结构和几何特征对光照和水分的影响,使能量运输和分配在冠层有着较大的差异[4],植物在长期适应环境条件的过程中,导致光合特性在冠层空间上呈现出空间异质性[5, 6]。辽东栎(Quercuswutaishanica) 是中国亚热带和暖温带常绿落叶阔叶混交林的主要优势种之一,对其分布区生态平衡、水土保持等有着重要的影响[7]。前人对辽东栎的光合生理生态特性、光响应曲线以及与外界环境因子之间的关系进行了报道[8-11],但对辽东栎冠层空间上光合生理特性的空间异质性研究较少。本文通过野外实际测量树冠内不同层次和不同方位的光合作用参数,定量分析了辽东栎冠层光合生理特性在冠层空间上的差异,旨在为在冠层水平上精确估算植物生产力,以及进一步估算落叶阔叶林生产力提供参考。
1 研究地区与研究方法
1.1 自然概况
研究地位于北京东灵山地区(40°00′~40°03′N,115°26′~115°30′E),地处北京市西郊的门头沟区,与河北省毗邻,距市区约100 km。东灵山属太行山脉,海拔高度800~2303 m,为典型暖温带大陆性季风气候。年均气温约为27℃,年平均降水量约为500 mm,且多集中在5~8月份,为总降水量的75%左右。冬季受西伯利亚和蒙古高压的影响,多西北风,寒冷干燥;夏季受东南海洋气流向北移动的影响,雨水充足。土壤主要为棕壤。东灵山地带性植被多为落叶阔叶林,分布广泛,构成了基本的山地形态。其他植被类型有落叶阔叶灌丛、温性针叶林、寒温性针叶林和草甸[12]。
1.2 研究方法
以北京东灵山研究地辽东栎林为研究对象,随即设置5个样地(20 m ×20 m),进行样地调查,确定标准木。每个样地选择一株标准木,共5棵。将标准木的树冠等分为上、下2个冠层,在每个冠层分东南西北4个方向。
于2013年7月—8月植物生长旺期,利用LI-6400便携式红外气体分析仪(美国LI-COR公司生产),采用红蓝光源6400-02B进行控制环境的光响应测定,光响应测定的光强梯度由强至弱依次设置为1800、1600、1400、1200、1000、800、600、400、200、150、100、50、20和0 μmolm-2s-1。测试时间选在晴朗天气的上午9:30—11:30。在辽东栎不同冠层不同方向选取3片无损伤的叶片测定,取其平均值作为实测数据。输出参数包括光合有效辐射(PAR,μmol m-2·s-1)、气温(Ta,℃)、气孔导度(Cond,mol H2O m-2·s-1)、水气压亏缺(VpdL,KPa)、大气相对湿度(RH,%)、蒸腾速率(Tr,mmol H2O m-2·s-1)等。利用模型拟合计算光合作用的光补偿点(LCP)、光饱和点(LSP)、饱和光合速率(Amax)、表观光合量子效率(AQY)和暗呼吸速率(RD)等指标[13]。
1.3 数据处理
光合作用的光响应曲线采用 Prado等[14]的非线拟合方程:
Pn =Amax (1-e-k (PAR-P))
Amax为饱和光合速率,k是常数,Pn为实际测定的净光合速率,PAR为光合有效辐射,LCP、LSP按最大净光合速率为90%时的PAR求得。
应用Excel软件进行响应数据的整理,应用Origin绘图软件进行曲线的绘制,应用SPSS16.0软件对试验数据进行方差分析。
通径分析可以把各生理生态因子对光合速率的相关系数细分为直接和间接的影响, 然后确定每个因子对总体相关性的相对贡献大小[15]。在本文中,设定光合速率为因变量,Cond、PAR、Tr、VpdL、RH、Ta为自变量,各因子的直接通径系数和间接通径系数的计算参照文献[16]。
2 结果与分析
2.1 光响应曲线在辽东栎冠层空间上的变化
从图1可以看出:从冠层底部到上部,光响应曲线的曲率逐渐减小,从光饱和到光抑制的变化幅度降低,表明冠层底部叶片对强光的适应能力较差;不同于冠层垂直方向的变化,叶片的光响应曲线在水平方向上的变化差异相对较小,东南方向的叶片略大于西北方向的。同时,通过对光响应曲线的数学拟合可以得到Amax、AQY和RD等参数,从而定量地呈现冠层在不同方位上叶片对光变化的响应特征。

图1 辽东栎冠层不同空间位置的光响应曲线
2.2 光合生理特性在辽东栎冠层空间上的变化
由图2可以得出:在垂直方向上,辽东栎冠层光合生理特性在冠层上部出现最高值的为Amax、RD、LCP和LSP,分别为9.22 μmol CO2m-2·s-1、1.593 μmol CO2m-2·s-1、755 μmol m-2·s-1和10.3 μmol m-2·s-1,最高值出现在冠层底部的只有AQY,为0.082 μmol CO2m-2·s-1。表明在辽东栎冠层垂直空间上,上部叶片的光合活性和生产力较高,但AQY较低,适应强光的能力高;相比较下冠层底部的光合活性较低,但具有较高的光能利用效率,可以充分利用散射光进行物质的积累。
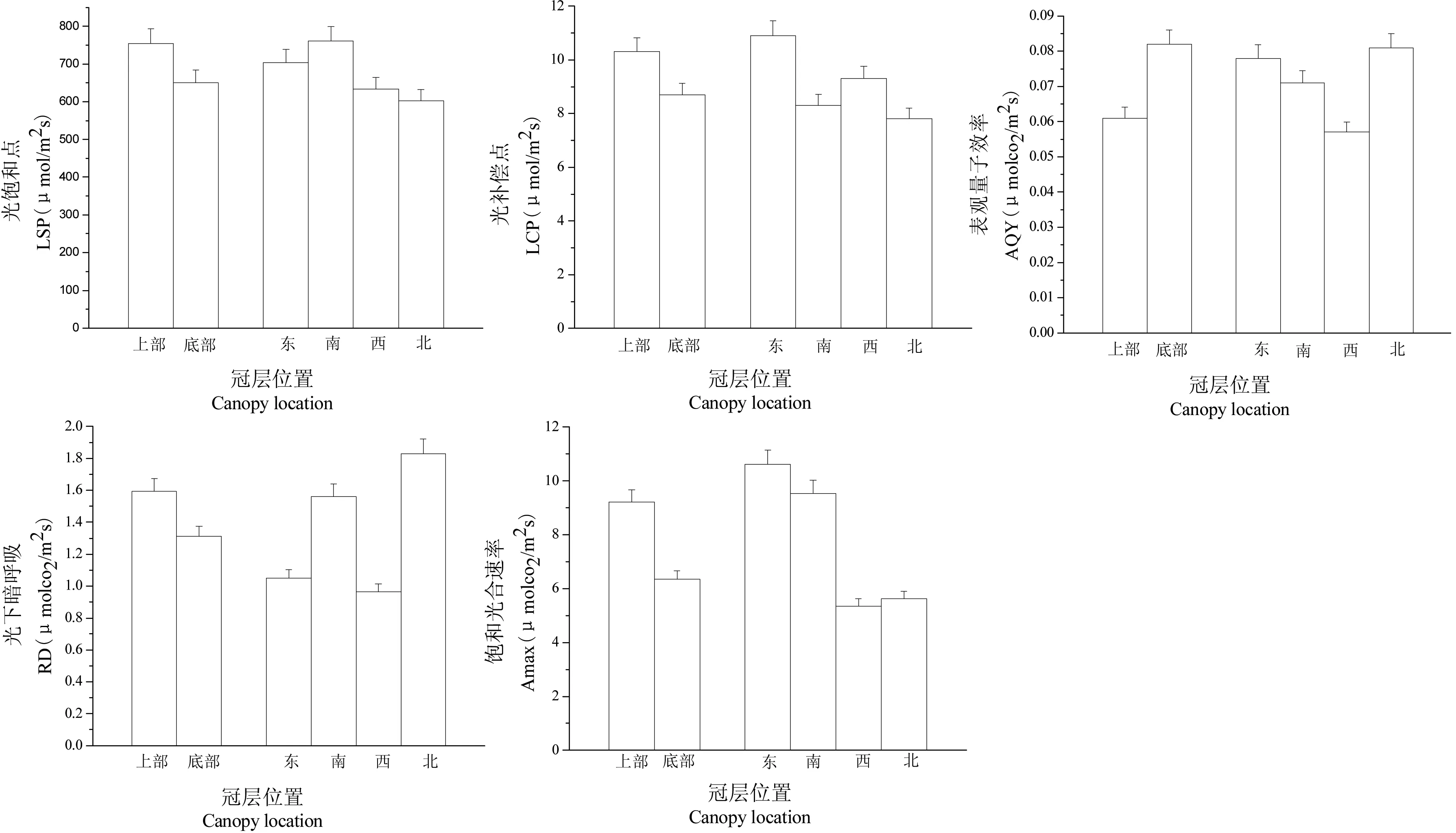
图2辽东栎冠层光合生理特性的空间异质性
Fig 2 Spatial differences of photosynthetic characteristics in the canopy ofQuercuswutaishanica
辽东栎冠层光合生理特性水平方向上的分布规律为:冠层东南向叶的Amax、LCP和LSP较高,分别比最低值高38.6%、16.9%和32.3%,而RH和AQY则为西北向叶较高。除LCP之外,各方向叶片的其他光合生理特性均有显著差异(P<0.05)。
2.3 主要生理生态因子对辽东栎不同层次、不同方向光合速率的影响
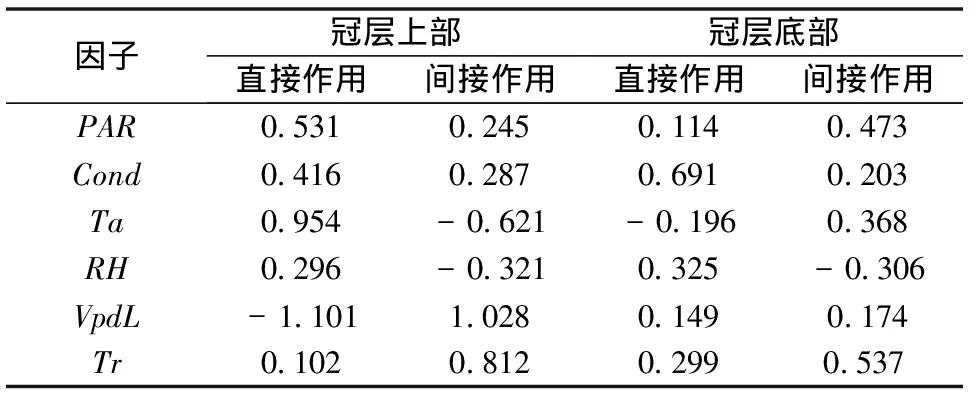
表 1 辽东栎冠层不同层次生理生态因子对光合速率的通径结果
PAR代表光合有效辐射、Ta代表温度、Cond代表气孔导度、VpdL代表大气水气压亏缺、RH代表大气相对湿度、Tr代表蒸腾速率。
光合速率可以最直接地表现植物光合作用的强弱,它的发生过程也受到许多外在和内在因素的共同影响[17]。为了更好地分析这些影响因素对光合速率的直接和间接作用在冠层空间上的异同,利用通径分析分别分析了主要生理生态因子对各层次各方向光合速率的影响大小。由表1可知,在冠层上部水气压亏缺、温度、光合有效辐射对光合速率的直接作用较大,而水气压亏缺、蒸腾速率和温度的间接作用较大;在底部气孔导度和蒸腾速率分别有着最大的直接和间接作用。
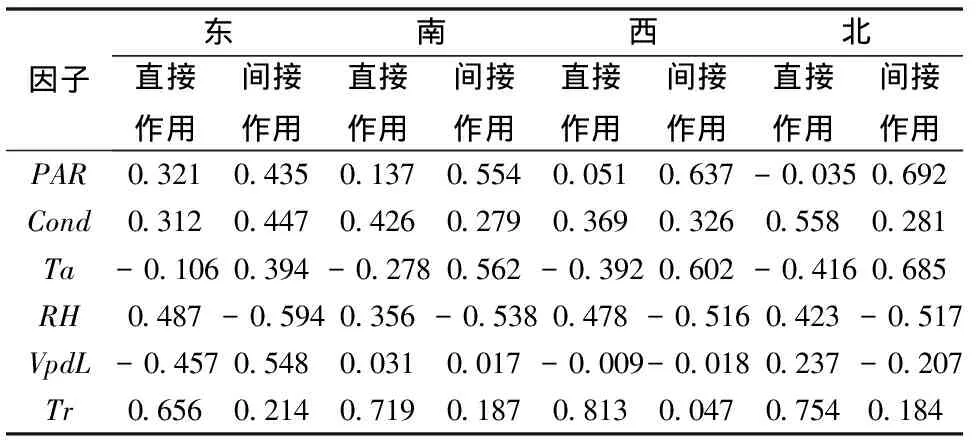
表2 辽东栎冠层不同方位生理生态因子对光合速率的通径结果
PAR代表光合有效辐射、Ta代表温度、Cond代表气孔导度、VpdL代表大气水气压亏缺、RH代表大气相对湿度、Tr代表蒸腾速率。
表2显示,蒸腾速率在冠层东、南、西3个方位都表现出最大的直接作用,而湿度、水气压亏缺在东方向的间接作用较大,光合有效辐射、温度在西、北方向的间接作用较大。
3 结论与讨论
饱和光合速率(Amax)是衡量植物光合能力的重要指标,不同树种不同叶片生长方式都会对其产生影响[13]。本研究中,在辽东栎冠层垂直空间上,叶片的Amax随着冠层部位的下降而减小。Harley[18]等曾通过研究得出白橡(Quercusalba)和红枫(Acerrubrum)冠层上部叶的饱和光合速率是底部叶的1.4倍。可见,在垂直空间上冠层的光合生理特性具有显著差异。引起辽东栎叶片饱和光合速率冠层差异的可能原因是由于叶片长期适应冠层内不同的光环境,使其内在结构和生理代谢等产生变化[19]。通常,辽东栎上部叶片光照条件要较为优越,而下部叶片长期处于遮荫或部分遮荫状态,因而只能获得较低的光照强度和较少的光照时间[20]。
光饱和点、光补偿点、表观量子效率、暗呼吸是植物对光利用能力的重要指示指标。一般认为,光饱和点越高,植物利用强光的能力越强;光补偿点越低,植物利用弱光的能力越强;表观量子效率则反映叶片对光能的利用效率[21]。通过四者在辽东栎冠层空间上的变化,可以看出:辽东栎冠层上部叶片有着较高的光合活性,利用强光的能力强;而冠层底部虽然光合活性低但利用弱光的能力强,从而使冠层利用光能的效率最高。辽东栎在冠层不同空间部位上光合特征产生的变化,可能与叶片自身的内在结构有关,如酶活性、叶绿素含量等[22],还可能与外界环境因子相关,如光照、水分、温湿度等。
植物长期生长在自然环境中,由于其冠层的几何结构特征,不同空间上叶片接受的光照和空气温湿度差异较大,使得冠层内部的微气候环境呈现出不同水平,导致树冠不同层次叶片的光合能力显著差异[23]。研究得出,水气压亏缺与光合速率呈现显著负相关关系,温度则与光合速率呈现显著正相关关系[24], 且水气压亏缺和温度经常共同对光合速率产生综合影响[25]。本文运用通径分析得出,水气压亏缺和温度在辽东栎冠层上部对光合速率有最大的直接和间接作用,与上述结论相统一。一般认为,气孔是植物进行CO2和水汽交换的重要通道,气孔导度是反映这种交换能力的关键指标[26]。而且蒸腾速率与气孔导度有着十分紧密的联系,二者都是影响光合速率的重要生理指标[27], 在本研究结果中也体现了这一结论。
导致冠层光合生理特性产生空间异致性的影响因素有很多,其中最重要的是光环境的变化[28]。由于冠层结构的几何特征,使得光在传递过程中呈现一定的规律性,从冠层底部到上部光通量呈指数函数增加;同时由于上层叶片对光合有效辐射的选择性吸收和反射,冠层下部的光强明显减弱,不足冠层上部的百分之十[13]。此外,提供植物光合作用的物质能量和水分大部分来自蒸腾作用,光环境不仅直接影响光照条件,还间接影响水分、温湿度等, 进而影响到水分利用效率和蒸腾速率。同时,还应考虑到叶片在生长凋落过程中,新叶着生位置的光环境会对叶片的形态生成造成直接影响,再加上叶龄、季节变化[29]等对叶片光合生理特性的影响,使得冠层光合作用的研究具有一定挑战性。
参考文献:
[1]李海梅,何兴元,陈 玮,等.沈阳城区五种乔木树种的光合特性[J].应用生态学报,2007,18(8):1709-1714.
[2]李海梅,何兴元,陈 玮.沈阳城市森林主要绿化树种——丁香的光合特性研究[J].应用生态学报,2004,15(12):2245-2249.
[3]宋庆安,童方平,易霭琴,等.虎杖光合生理生态特性日变化研究[J].西北植物学报,2007,27(9):1909-1914.
[4]张小全,徐德应,赵茂盛.林冠结构、辐射传输与冠层光合作用研究综述[J].林业科学研究,1999,12(4):411-421.
[5]徐炳成,山 仑,黄 瑾.黄土丘陵区不同立地条件下沙棘光合生理日变化特征比较[J].西北植物学报,2003,23 (6): 949-954.
[6]王满莲,蒋水元,李 锋, 等.不同立地条件罗汉果组培苗的光合特性[J].广西植物,2010, 30(4):507-512.
[7]康永祥,岳军伟,张巧明.黄龙山林区辽东栎群落类型划分及其生物多样性研究[J].西北林学院学报,2007,22 (3):7-10.
[8]蔺 琛,马钦彦,韩海荣,等.山西太岳山辽东栎的光合特性[J].生态学报,2002,22(9): 1399-1406.
[9]白宏兵,吴发启,程 伟.辽东栎幼苗光合生理生态特性对施氮响应的初步研究[J].草地学报,2013 (001):100-108.
[10]秦 娟,上官周平.子午岭林区山杨-辽东栎混交林的生理生态效应[J].应用生态学报,2006,17 (6):972-976.
[11]蒙乾财,张晓飞,刘 晓,等.陕北黄土高原辽东栎林分布区北部边界形成的光合作用机制[J].应用生态学报, 2011, 22(6): 1409-1415.
[12]王 巍,李庆康,马克平.东灵山地区辽东栎幼苗的建立和空间分布[J].植物生态学报,2000,24(5): 595-600.
[13]张国平,周伟军.植物生理生态学[M].杭州:浙江大学出版社,2005.
[14]Prado C H B A, de Moraes J A P V.Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field conditions[J].Photosynthetica, 1997, 33:103-112.
[15]Condon A G, Richards R A, Rebetzke G J. Breeding for high water-use efficiency[J]. Journal of Experimental Botany, 2004, 55(407):2447-2460.
[16]任红松,吕 新,曹连莆,等.通径分析的SAS 实现方法[J].计算机与农业, 2003, 4: 17-19.
[17]许大全,徐宝基,沈允刚.C3植物光合效率的日变化[J].植物生理学报,1990, 16(1):1-5.
[18]Harley P C, Baldocchi D B. Scaling carbon dioxide and water vapor exchange from leaf to canopy in a deciduous forest Leaf model parameterization[J]. Plant, Cell and Environment, 1995, 18: 1146-1158.
[19]严昌荣,韩兴国,陈灵芝.北京山区落叶阔叶林优势种叶片特点及其生理生态特性[J].生态学报,2000,20(1): 53-60.
[20]孙书存,陈灵芝. 东灵山地区辽东栎叶养分的季节动态与回收效率[J]. 植物生态学报, 2001, 25(1): 76-82.
[21]孟 陈,徐明策,李俊祥,等.栲树冠层光合生理特性的空间异质性[J]. 应用生态学报, 2007, 18(9): 1932-1936.
[22]安 慧,上官周平.黄土高原植被不同演替阶段优势种的光合生理特性[J].应用生态学报, 2007, 18(6): 1175-1180.
[23]郑 元,赵 忠,周 慧,等.刺槐树冠光合作用的空间异质性[J]. 生态学报, 2010, 30(23): 6399-6408.
[24]Horton J L, Kolb T E, Hart S C. Leaf gas exchange characteristics differ among Sonoran Desert riparian tree species[J]. Tree Physiology, 2001, 21:233-241.
[25]Barron-Gafford G A, Grieve K A, Murthy R. Leaf-and stand-level responses of a forested mesocosm to independent manipulations of temperature and vapor pressure deficit[J].New Phytologist, 2007, 174:614-625.
[26]张小全, 徐德应, 赵茂盛, 等.CO2增长对杉木中龄林针叶光合生理生态的影响[J].生态学报, 2000, 20(3): 390-396.
[27]刘遵春,包东娥.‘金光杏梅’叶片净光合速率与生理生态因子的关系[J].西北植物学报,2008,28(3): 564-568.
[28]Lloyd J, Shibistova O, Zolotoukhine D, et al. Seasonal and annual variations in the photosynthetic productivity and carbon balance of a central Siberian pine forest[J]. Tellus, 2002, 54: 590-610.
[29]张小垒,徐德应. 杉木中龄林不同部位和叶龄针叶光合特性的日变化和季节变化[J].林业科学,2000, 36(3): 19-26.
