西太平洋富钴结壳超微化石与分子化石地层划分及对比*
2014-03-19雷吉江李小虎张海生初凤友
雷吉江 李小虎 张海生 初凤友
(1. 国家海洋局第二海洋研究所 杭州 310012; 2. 国家海洋局海底科学重点实验室 杭州 310012;3. 国家海洋局海洋生态系统与生物地球化学重点实验室 杭州 310012)
富钴结壳富含Co、Pt等金属元素, 是最重要的海底矿产资源之一(Heinet al, 2000), 并且因生长速率缓慢, 记录下了超过60Ma的古环境信息, 被视为古海洋学研究的重要载体(Franket al, 1999)。近30年来, 富钴结壳的研究备受关注, 人们在富钴结壳分布规律、组成特征、生长机制、资源价值、年代框架以及古海洋学记录等方面取得了一系列重要的认识(Heinet al, 1988; Cowenet al, 1993; Banakaret al,2000; 苏新等, 2004; 初凤友等, 2005; Usuiet al, 2007;李江山等, 2008; Heinet al, 2009; Glasby, 2010; 任向文等, 2011; Muiñoset al, 2013)。
年代学研究是认识大洋环境历史演化问题的基础, 目前对富钴结壳测年的方法主要有同位素测年法、基质年龄推算法和古生物地层学法等。其中, 古生物地层学法根据超微化石的种属及形态确定地层时代或进行地层划分对比, 由于壳层中保存的超微化石通常含量丰富、分布广泛, 并具有生物演化快、垂直分布短暂和标本处理简单的特点, 且古生物地层学法测年结果不受结壳生长间断的影响, 相对而言该方法更简便可靠(潘家华等, 2007)。
分子化石是保存于地质体中、记录了原始生物母质信息、在演化过程中保持碳-碳骨架不变的有机质分子, 通常被用于原油对比和古沉积环境恢复(Seifertet al, 1981)。Grantham等研究了不同地质年代海相源岩中C28和C29甾烷的含量变化, 指出不同时代源岩的甾烷分布不同(Granthamet al, 1988)。此后Summons等根据原油中正常甾烷含量的碳数分布特征来确定油源的年代(Summonset al, 1990)。随着传统地层学向多学科相互渗透的综合地层学方向发展,将微体古生物学和有机地球化学结合起来研究会取得更令人满意的结果。本文根据太平洋马尔库斯-威克海山群CM1D03、麦哲伦海山群CM3D06富钴结壳中钙质超微化石生物地层学记录, 划分出年代框架, 尝试建立甾烷分布构型与富钴结壳生长壳层的协变关系, 为今后富钴结壳地层划分及对比研究提供支撑。
1 样品与方法
1.1 样品及特征
富钴结壳样品由“大洋一号”科考船DY105-12航次于太平洋海山区利用拖网取得, CM1D03、CM3D06结壳厚度分别为100mm、85mm, 具典型的三层构造。
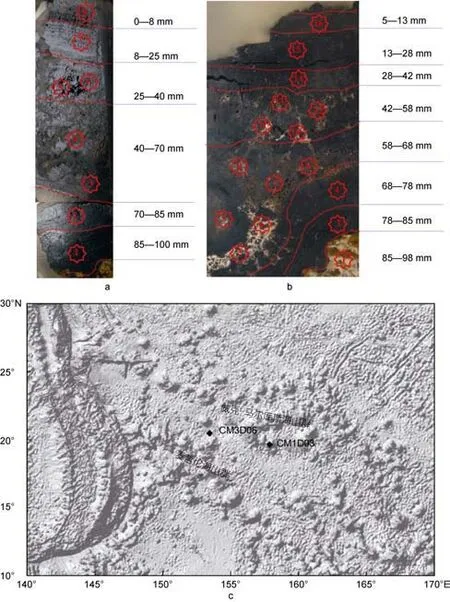
图1 研究样品及站位(a为CM1D03, b为CM3D06, c为站位分布图)Fig.1 Samples and sampling station (a: CM1D03; b: CM3D06; c: sampling station)
根据壳层构造特征, 将结壳样品划分出多个亚层(图1a、b)。CM1D03结壳内层(70—100mm)为亮黑色、致密老壳层, 硬度较大, 电子显微镜下呈柱状结构, 黄褐色碳氟磷灰石和碳酸盐以“胶结物”形式渗染于其间; 中间层(25—70mm)呈褐黑色、伴有棕黄色细脉, 质地疏松, 镜下局部呈枝状结构、其间夹杂大量岩屑块; 外层(0—25mm)呈碳黑色、具强金属光泽,较致密, 硬度较大, 镜下以柱状结构为主, 几无碎屑物质, 层间色泽变化不大。CM3D06结壳与CM1D03相似, 内层(68—85mm)为亮黑色, 质地致密(亮煤层),电子显微镜下以纹层-柱状结构; 中间层(28—68mm)质地疏松, 镜下显微结构呈多种构造叠层状, 如斑杂状构造、树枝状构造、指纹构造等, 层间夹杂大量脉石矿物和黄色碎屑物质, 与上下层界限分明; 外层(0—28mm)呈暗黑色, 较致密, 镜下为柱状构造、树枝状构造等, 夹杂一定量的脉石矿物。CM3D06样品85mm以下为灰褐色磷块岩。
1.2 微体古生物分析
对结壳典型的每个构造层的上、中、下的2—3个层位进行精细取样, 分别在每一壳层剥离2—5mm宽的新鲜样品(2—3片), 制作薄片并镀一层薄金膜,使用扫描电子显微镜(SEM)进行钙质超微化石鉴定。SEM工作条件: 加速电压18kV, 放大倍数: 1´103—2´105倍, 该工作由俄罗斯State Scientific Centre Yuzhmorgeologia Gelendzhik的Pulyaeva教授完成。
1.3 分子化石分析
对两块结壳样品的I—VI层分别取样, 在低于50°C条件下烘干, 粉碎至150μm以下, 用体积比为3︰1的二氯甲烷和甲醇混合溶液进行抽提, 抽提物经旋转蒸发至干, 得到样品中的可溶有机质(氯仿沥青“A”)。将上述浓缩液用正己烷沉淀脱去沥青质, 利用硅胶柱进行层析分离, 分别用正已烷、苯和二氯甲烷/甲醇(1︰1)淋洗分离出饱和烃、芳烃、非烃组分。选取饱和烃组分进行气相色谱(GC)和色谱-质谱(GC-MS)分析。
GC分析条件: HP5890气相色谱仪, 弹性石英毛细管柱(DB-5, 30m´0.25mm内径、0.17µm涂层厚度)。升温程序: 初始温度80°C, 升温速率5°C/min, 终止温度280°C, 保持30min。GC-MS分析条件: TSQ7000色质联用仪, HP-5MS石英毛细管柱(60m×0.25mm内径、0.25µm)。程序升温: 初始温度80°C, 恒温2min——升温速率8°C/min至220°C——升温速率2°C/min至300°C, 恒温保持25min。质谱条件: 离子源温度: 250°C; 离化电流: 150µA; 电子能量70eV,扫描周期1s以上。
2 结果
2.1 钙质超微化石地层年代
钙质超微化石地层年代的划分主要根据Martini(1971)建立的第三纪化石分带(NP1-NP25), 以及Bukry(1975)、Okada和Bukry(1980)建立的早第三纪19个带(CP1- CP19)和晚第三纪以来15个带(CN1-CN15)。CM1D03老结壳100—70mm层段, 钙质超微化石化种类较少,Discoaster multiradiatus是NP9的标志种, 出现并繁盛于晚古新世至早始新世, 对应于钙质超微化石带NP9,Coccolithus pelagicus(图2-1)时代跨度较大, 向上可以延续到始新世(中、晚期), 形成时代为晚古新世–早始新世(Martini, 1971; Okadaet al,1980); 70—25mm化石群组合为Calcidiscus simplex、Reticulofenestra reticulata(图2-2、图2-3), 对应于化石带NP14—NP20, 形成时代为中始新世; 25—8mm层段Cyclicargolithus floridanus(图2-4)对应于Bukry(1975)钙质超微化石带CN3—CN5a, 形成时代为早中新世-晚中新世; 新结壳层8—0mm识别出更新世的钙质超微化石组合, 其中Pseudoemiliania lacunosa(图2-5)最早出现于早上新世(Berggrenet al,1985), 而Helicosphaera seliil(图2-6)是早更新世的一个标志种(Raffiet al, 1993), 综合推测该层段时代为早上新世-早更新世。从钙质超微化石的面貌来看,CM1D03结壳在古新世—始新世适宜的气候条件下开始生长, 地层年代跨度从晚古新世到更新世。
CM3D06富钴结壳底层98—85mm,Zigodicus spiralis(图4-1)、Watznaneria bconesae(图4-2)钙质超微化石-颗石藻是白垩纪远洋沉积中最为常见暖水种之一(Lamoldaet al, 1986), 时代划分为晚白垩纪—70Ma (或许更古老); 85—68mm化石群组合有相当大的层间变化, 总体上对应于Okada和Bukry(1980)划分的化石带CP8—CP9(对应于55Ma), 代表种包括Discoaster multiradiatus(图4-3)、Discoaster sp、Discoaster mohleri等(图4-4), 时代划分为晚始新世-早始新世(Bukry, 1975); 68—58mm化石群组合为Coccolithus formosus、Reticulofenestra coentura、Chiasmolithus consuetus、Sphenolithus moriformis、Transversopontis pulcher、Ericsonia fenestrate, 结壳地层形成时代对应于NP14-NP20(中始新世-晚始新世); 58—42mm代表种为Trochoaster duplex(图4-5),结壳形成时代对应于NP14-NP20或CP12-15(中始新世-晚始新世); 42—28mm形成时代对应于CN3—CN5a (早中新世-晚中新世)(Bukry, 1975); 5.0—28.0mm代表种为Umbilicosphaera sibogae(图4-6),结壳形成时代划分为上新世-更新世(Gibbsaet al,2005)。根据超微化石的全貌来看, 这是一个从晚白垩纪——或可追溯到更古老地层到更新世的地层序列。
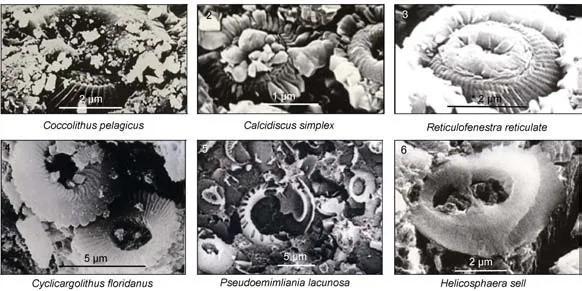
图2 CM1D03富钴结壳代表性钙质超微化石电子扫描显微照片Fig.2 Images of scanning electron micrography showing representative calcareous nannofossils in the cobalt-rich crust CM1D03

图3 CM3D06代表性钙质超微化石电子扫描显微照片Fig.3 Images of scanning electron micrography showing representative calcareous nannofossils in the cobalt-rich crust CM3D06
钙质超微化石化石观察结果显示, CM1D03、CM3D06均具有代表性的钙质超微化石组合, 能很好地反映出富钴结壳的生长时代。地层划分结果表明,两块样品在相同时代的钙质超微化石组合差别较大,反映出两站位水体沉降环境和结壳生长环境可能存在较大差别, 有利于对比研究。
2.2 分子化石地层
结壳甾烷分布构型的演化具有时代特征, 通过分析古海洋学事件对海底沉积层甾烷输入的影响,结合环境参数的变化研究, 可以大致确定不同分布构型所代表的时代。
利用特征离子m/z=217识别了富钴结壳层中的甾烷分子(图4)。C27-、C28-、C29甾烷的相对丰度显示, CM1D03、CM3D06的甾烷分布构型具有明显的分层特征: 从老至新分别为反“L”型、C27优势的“V”字型和C29优势的“V”字型(表1、表2)。对比两块结壳的构造分层及钙质超微化石地层发现,CM1D03与CM3D06结壳尽管均具有典型的三层结壳构造, 但各壳层所处的生长时代存在较大差别, 这可能与结壳成矿物质来源、基质条件等因素有关。两块富钴结壳中的甾烷构型变化在结壳生长时代上具有很好的一致性, C29优势的“V”字型对应中新世及以后, C27优势的“V”字型对应始新世, 反“L”型甾烷构型对应晚古新世及以前。由于富钴结壳中的甾烷主要由外源输入, 因此, 决定富钴结壳中甾烷构型分布的因素可能主要与甾烷的输入源有关。
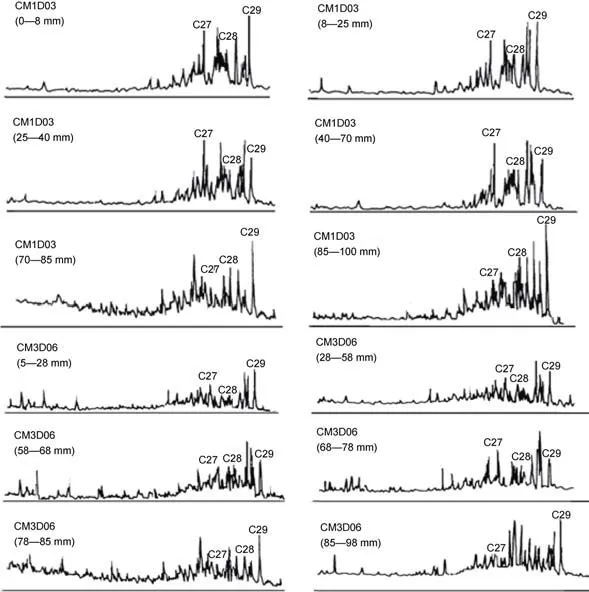
图4 CM1D03、CM3D06富钴结壳m/z=217质量色谱图Fig.4 Mass chromatogram of sterane(m/z=217) from the cobalt-rich crusts CM1D03 and CM3D06
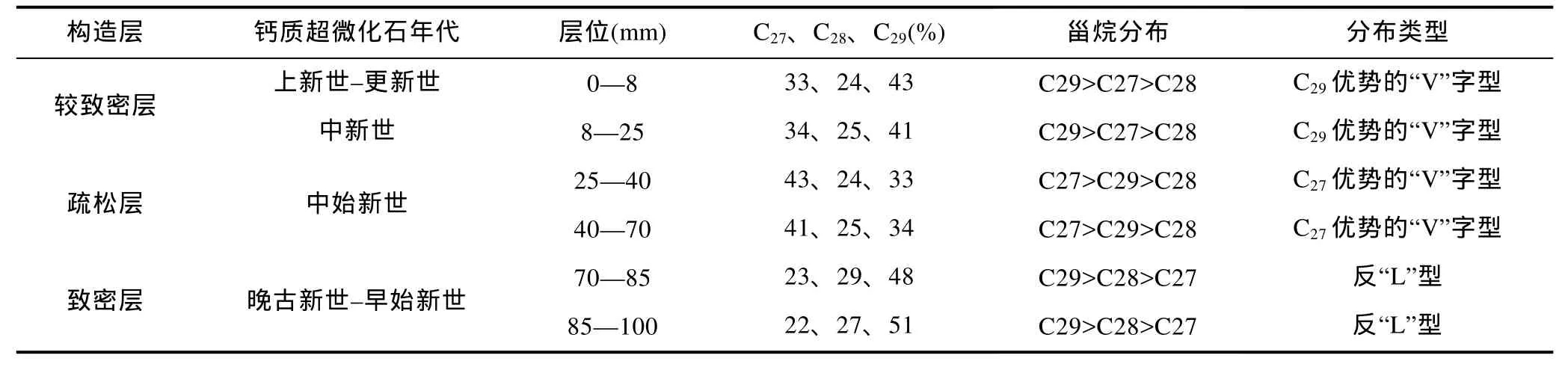
表1 CM1D03富钴结壳层甾烷分布特征Tab.1 The distribution of sterane from cobalt-rich crust CM1D03
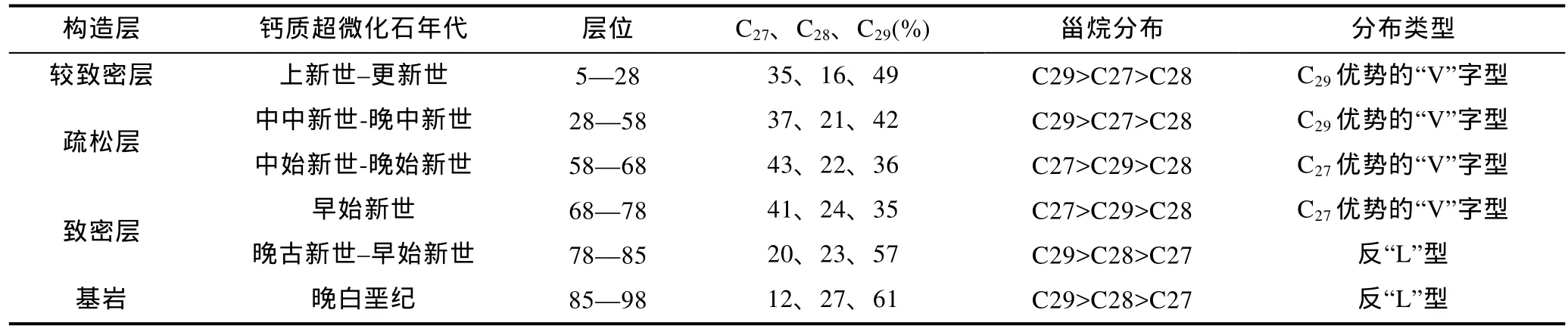
表2 CM3D06富钴结壳层甾烷分布特征Tab.2 The distribution of sterane from cobalt-rich crust CM3D06
3 讨论
甾类化合物主要来源于生物体中的甾醇类化合物,是反映生物有机质输入的常用参数之一(Mackenzieet al, 1982)。在有机质演化过程中, 甾醇通过加氢脱羧基转变为相同碳数的甾烷, 由于C27、C28、C29甾烷具有相同的热演化速率, 这使它们的相对含量不受或很少受成熟度的影响, 故能够反映原始母质中C27、C28、C29甾烷的比例, 是判识有机质类型、恢复古环境的重要指标之一(Granthamet al, 1988)。一般来讲, C27甾烷主要来源于海洋浮游动物, C28甾烷在海洋中更多地来自海洋中的硅藻; 而C29甾烷可能来自陆源高等植物, 但由于研究区远离陆地, 附近海沟、海山构造发育, 陆源有机质难以输送至此, 富钴结壳中的C29甾烷并非陆地高等植物输入, 其可能来自蓝绿藻(Volkman, 2006)。
测年数据表明, 马尔库斯-威克海山群、麦哲伦海山群的年龄介于85.5—101.6Ma之间(赵俐红等,2010), 因此, 海山上富钴结壳记录的古海洋学信息始于晚白垩世之后。白垩纪末期, 地球发生了一次导致生物灭绝的灾变事件, 海洋表层生产力急剧下降,依赖浮游植物为生的海洋浮游动物受到重创(许靖华等, 1980), 以致进入古新世海洋浮游动物生物量仍十分有限, 限制了C27甾烷向海底沉积层的输入, 而蓝绿藻作为最简单、原始的原核生物种群成为晚白垩纪-古新世海底沉积层主要的甾烷输入源, 在富钴结壳层中形成以C29占主要优势的甾烷分布构型。白垩纪末的硅藻灭绝程度并不严重, 近46%的硅藻物种继续存活到上古新世(MacLeodet al, 1997), 使得海底甾烷分布中C28甾烷占有较高比例。古新世的世界大洋以白垩纪时形成的洋流为主, 由面积较大边缘海的蒸发作用造成温暖高盐水下沉, 形成温暖、高盐度的底层海水, 由于氧气在水中的溶解度随温度增高而降低, 因而晚白垩纪-古新世的结壳层属于偏还原的生长环境。
早始新世进入气候最佳期, 此时南极地区开始降温, 中层水的循环开始加强, 从而赤道上升流也得到加强, 古生产力增加(Corfield, 1994), 浮游动物繁盛, 为海底沉积层提供了主要的甾烷来源, 此时结壳层中的甾烷出现以C27甾烷为主要优势的“V”字型分布特征, 而第三纪浮游生物分布研究显示, 始新世期间浮游有孔虫、颗石藻以及腰鞭毛藻等浮游生物量明显增加, 始新世过后明显降低(Bujaket al, 1979), 因此, C27甾烷优势可能是始新世期间的甾烷构型。
始新世与渐新世之交发生了一次大的变冷事件,与早始新世的最佳气候恰成对立, 造成海洋生物群落的长时间(长达4Ma)灭绝(Kaiho, 1994), 同时突发的天体撞击事件、气候急剧变冷对底流活动产生影响,打破结壳层生长的平衡状态, 导致明显的结壳生长间断(丁旋等, 2008)。
在中新世早期, 南极冰盖退缩, 迎来了海进期,全球气温开始回暖, 表层水和底层海水温度上升(Haq, 1981)。南极底流萎缩, 造成太平洋表层生产力下降, 在海水环境中普遍存在的蓝藻成为海底甾烷的主要输入源, 其次为浮游动物, 此外陆相有机质的增加也提供了一部分C29甾烷输入, 海底甾烷构型具有C29优势的“V”字型特征。海底表层水温的上升以及南极底流的萎缩造成结壳层的生长处于缺氧的还原环境, 生长速率较慢, 形成致密的结壳层。碳同位素研究也表明, 结壳层后期的生长主要与气候回暖事件以及南极底层流萎缩相关联(史跃中等, 2004)。
晚中新世以后, 南极冰盖扩张, 底层水环流增强,大洋肥力增加, 表层生产力逐步回升(丁旋等, 2008)。底层水氧化性增强, 海底沉积层中甾烷构型发生变化, C27输入比重增加, ODP钻孔资料研究表明, 渐新世以来, 海底沉积物中的硅藻沉降通量较低且稳定,但晚中新世开始急剧增加, 导致海底沉积物中硅质软泥占有绝对优势, 尽管上新世晚期硅藻沉降通量开始降低, 海底沉积物中显著的硅藻通量持续至第四纪(方孝悌, 1994; Reaet al, 1995)。因此, 上新世期间甾烷可能呈C27、C28为主的“L”型分布。
从表3可知, 本文结壳层中从老到新的甾烷分布构型分别与晚白垩世-古新世、早始新世-晚始新世、早中新世-晚中新世三个阶段的甾烷输入特征相吻合,因此可以认为富钴结壳层中甾烷构型的变化主要是古海洋学事件演化的影响, 具有一定的时代性, 能用于富钴结壳层的地层年代划分。
对比钙质超微化石与分子化石地层发现, 两种方法划分出的地层存在一些差异, 其原因可能与取样位置和年代指示的精度不同有关。尽管如此, 两种方法在富钴结壳层主要生长时代及生长间断的界定上具有一致性, 可以用于富钴结壳的地层划分和对比。
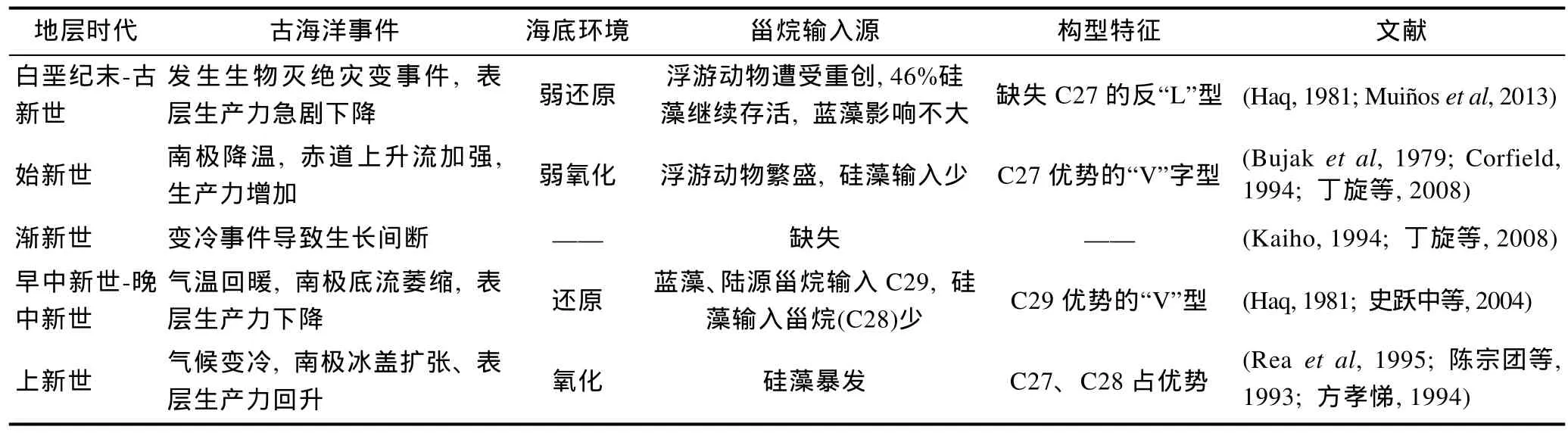
表3 古海洋环境演化与太平洋富钴结壳的甾烷输入Tab.3 The paleoceanographic evolution and sterane input in the cobalt-rich crust from the Pacific Ocean
4 结论
(1) 根据钙质超微化石记录划分出富钴结壳层生长时代: CM1D03分为晚古新世-早始新世、始新世中期、中中新世-更新世; CM3D06分为晚古新世-早始新世、中始新世–晚始新世、中中新世-晚中新世、上新世-更新世。
(2) 结壳层的甾烷分布特征存在一定变化趋势,从老至新分别为: 反“L”型、C27优势的“V”型、C29优势的“V”型。古海洋事件演化与富钴结壳甾烷输入的关系表明, 此三种甾烷构型分别是晚白垩世-古新世、早始新世-晚始新世、早中新世-晚中新世三个生长阶段的特征, 因此, 富钴结壳中的甾烷构型可能具有一定的时代性, 可以作为参考指标用于富钴结壳生长时代和间断的划分。
(3) 钙质超微化石和分子化石地层学划分方法在富钴结壳层主要生长时代及生长间断的界定上具有一致性, 可以用于富钴结壳的地层划分和对比。
陈宗团, 许东禹, 1993. 东太平洋晚新生代生产力演化特征的初步研究. 海洋通报, 12(1): 43—48
初凤友, 孙国胜, 李晓敏等, 2005. 中太平洋海山富钴结壳生长习性及控制因素. 吉林大学学报(地球科学版), 35(3):320—325
丁 旋, 高莲凤, 方念乔等, 2008. 太平洋海山富钴结壳生长过程与新生代海洋演化关系. 中国科学(D辑: 地球科学),38(10): 1297—1308
方孝悌, 1994. 北太平洋新生代古海洋学的变迁——ODP第145航次的成果. 海洋地质译丛, 4: 74—77
李江山, 方念乔, 屈文俊等, 2008. 中太平洋富钴结壳的Os同位素定年与结壳生长间断. 中国科学(D辑: 地球科学),38(9): 1122—1129
潘家华, 张 静, 刘淑琴等, 2007. 西北太平洋富钴结壳的钙质超微化石地层学研究及意义. 地球学报, 28(5): 411—417
任向文, 刘季花, 石学法等, 2011. 麦哲伦海山群M海山富钴结壳成因与成矿时代: 来自地球化学和Co地层学的证据.海洋地质与第四纪地质, 31(6): 65—74
史跃中, 胡超涌, 方念乔等, 2004. 富钴结壳中有机碳同位素组成特征及其古海洋意义. 地球科学, 29(2): 148—150
苏 新, 马维林, 程振波, 2004. 中太平洋海山区富钴结壳的钙质超微化石地层学研究. 地球科学, 29(2): 141—147
许靖华, 何起祥, 1980. 彗星冲击作用——白垩纪末期地球上发生灾变的原因. 长春地质学院学报, 2: 1—8
赵俐红, 齐 君, 杨 慧, 2010. 火山岩浆活动与富钴结壳成矿间的关系——以中太平洋海山群和麦哲伦海山链所在的中西太平洋区为例. 海洋地质动态, 26(2): 1—7
Banakar V K, Hein J R, 2000. Growth response of a deep-water ferromanganese crust to evolution of the Neogene Indian Ocean. Marine Geology, 162(2—4): 529—540
Berggren W A, Kent D V, Flynn J J, 1985. Jurassic to Paleogene:Part 2: Paleogene geochronology and chronostratigraphy. In:Snelling N. J ed. The Chronology of the Geological Record,Geological Society of London Memoirs, Vol. 10. London:Geological Society, 141—195
Bujak J P, Williams G L, 1979. Dinoflagellate diversity through time. Marine Micropaleontology, 4: 1—12
Bukry D, 1975. Coccolith and sillicoflagellate stratigraphy,northwestern Pacific Ocean, Deep Sea Drilling Project, Leg 32. Initial Reports of the Deep Sea Drilling Project, 32:677—701
Corfield R M, 1994. Palaeocene oceans and climate: An isotopic perspective. Earth-Science Reviews, 37(3): 225—252
Cowen J P, DeCarlo E H, McGee D L, 1993. Calcareous nannofossil biostratigraphic dating of a ferromanganese crust from Schumann Seamount. Marine Geology, 115(3—4): 289—306
Frank M, O'Nions R K, Hein J Ret al, 1999. 60 Myr records of major elements and Pb-Nd isotopes from hydrogenous ferromanganese crusts: reconstruction of seawater paleochemistry.Geochimica Et Cosmochimica Acta, 63(11—12): 1689—1708
Gibbsa S J, Young J R, Bralowerc T Jet al, 2005. Nannofossil evolutionary events in the mid-Pliocene: an assessment of the degree of synchrony in the extinctions ofReticulofenestra pseudoumbilicusandSphenolithus abies.Palaeogeography, Palaeoclimatology, Palaeoecology, 217(1):155—172
Glasby G P, 2010. Incorporation of Transition and Platinum Group Elements(PGE) in Co-rich Mn Crusts at Afanasiy-Nikitin Seamount (AFS) in the Equatorial S Indian Ocean.Resource Geology, 60(2): 212—215
Grantham P J, Wakefield L L, 1988. Variations in the sterane carbon number distributions of marine source rock derived crude oils through geological time. Organic Geochemistry,12(1): 61—73
Haq B U, 1981. Paleogene paleoceanography: Early Cenozoic oceans revisited. Oceanol Acta, 4(suppl): 71—82
Hein J, Koschinsky A, Bau Met al, 2000. Cobalt-rich ferromanganese crusts in the Pacific. In: Cronan, D S ed.Handbook of marine mineral deposits. Boca Raton, USA:CRC Press, 239—279
Hein J R, Conrad T A, Dunham R E, 2009. Seamount characteristics and mine-site model applied to exploration- and mining-lease-block selection for cobalt-rich ferromanganese crusts. Marine Georesources and Geotechnology, 27(2): 160—176
Hein J R, Schwab W C, Davis A, 1988. Cobalt- and platinum-rich ferromanganese crusts and associated substrate rocks from the Marshall Islands. Marine Geology, 78(3—4): 255—283
Kaiho K, 1994. Planktonic and benthic foraminiferal extinction events during the last 100 my. Palaeogeography,Palaeoclimatology, Palaeoecology, 111(1): 45—71
Lamolda M A, Proto-Decima F, 1986. The Turonian-Coniacian boundary in Ollogoyen (Basque Country); foraminifers and nannoplankton. Cretaceous Research, 7(1): 63—75
Mackenzie A S, Brassell S C, Eglinton Get al, 1982. Chemical fossils: The Geological fate of steroids. Science, 217(4559):491—504
MacLeod N, Rawson P F, Forey P Let al, 1997. The Cretaceous–Tertiary biotic transition. Journal of The Geological Society,154(2): 265—292
Martini E, 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation. In Farinacci A ed. Proceedings of the Second Planktonic Conference Roma 1970. Rome, Italy:Edizioni Tecnoscienza, 739—785
Muiños S B, Hein J R, Frank Met al, 2013. Deep-sea Fe-Mn crusts from the northeast Atlantic Ocean: composition and resource considerations. Marine Georesources and Geotechnology, 31(1):40—70
Okada H, Bukry D, 1980. Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation. Marine Micropaleontology, 5:321—325
Raffi I, Backman J, Rio Det al, 1993. Plio-Pleistocene nannofossil bbiostratigraphy and calibration to oxygen isotope stratigraphies from Deep Sea Drilling Project Site 607 and Ocean Drilling Program Site 677. Paleoceanography,8(3): 387—408
Rea D K, Basov I A, Krissek L Aet al, 1995. Scientific results of drilling the North Pacific Transect. In Rea D Ket aled.Proceedings of the Ocean Drilling Program, Scientific Results. 145: 577—596
Seifert W K. Moldowan J M, 1981. Paleo-reconstruction by biological makers. Geochimica et Cosmochimica Acta, 45(6):783—794
Summons R E, Walter M R, 1990. Molecular fossils and microfossils of prokaryotes and protists from proterozoic sediments. American Journal Science, 290(A): 212—244
Usui A, Graham I J, Ditchburn R Get al, 2007. Growth history and formation environments of ferromanganese deposits on the Philippine Sea Plate, northwest Pacific Ocean. Island Arc, 16(3): 420—430
Volkman J K, 2006. Marine Organic Matter: Biomarkers,Isotopes, and DNA. Berllin: Springer-Verlag, 52
