塔里木盆地古生界原油中高丰度C29规则甾烷的分布及意义
2021-03-25张敏程青松
张敏,程青松
1.长江大学资源与环境学院,湖北 武汉 430100 2.中国石油浙江油田分公司勘探开发一体化中心,浙江 杭州 310023
C27、C28、C29甾烷最早从美国绿河页岩和前寒武系页岩中鉴定出[1,2]。所有规则甾烷都由一个环戊烷基全氢化菲环(甾核)和烷基侧链组成[3]。甾烷来自生物体的甾醇,甾醇在成岩阶段厌氧环境微生物作用下,经氢化形成甾烷醇,氧化环境下甾醇会被优先降解[4-9],然后甾醇、甾烷醇脱水形成甾烯[10-12],最后甾烯还原形成甾烷[10-13]。因此,沉积物中甾烷的分布特征必然是继承了沉积有机质中甾醇类的分布特征。甾醇是真核细胞特有且至关重要的一类代表性脂质分子,甾醇参与细胞发育调控,是信号传导和激素分子的前体,对研究生命进化具有重要意义[14-15]。真核生物的细胞膜中存在丰富的甾醇,细胞膜中甾醇与磷脂的摩尔比很高,每个磷脂双链与2个甾醇分子接触[16]。正因为大量的甾醇存在才实现了对细胞膜复杂的调节作用。此外,HUANG等和MEINSCHEIN等通过对各类生物体和现代沉积物中C27、C28、C29这3种甾醇丰度和相对组成特征的研究,指出C27规则甾烷主要为海相来源,C29规则甾烷是高等植物来源[17,18]。通常,地质体中高丰度的C29甾烷被认为是较多陆源有机质输入的标志(因为C29甾烷的前身物为C29甾醇),如24-乙基胆甾-5,22-二烯醇 和 24-乙基胆甾-5-烯醇在维管束植物中很丰富[19]。值得注意的是,在阿曼元古代原油[20]、华北地区下古生界和元古界烃源岩[21]、塔里木盆地寒武系-下奥陶统和中上奥陶统烃源岩[22]中,也发现了高丰度的C29规则甾烷。然而上述烃源岩和原油形成的时代还未有高等植物出现。例如,在一些藻类如硅藻和绿藻中也有高丰度的24-乙基胆甾-5-烯醇和24-乙基胆甾-5,22-二烯醇[20,23,24]。因此,在古代沉积物和原油中C29甾烷的高比例不一定标志大部分有机质来自维管束植物。GRAY等[25]认为陆地维管束植物在泥盆纪才开始大面积繁盛,早古生代和前寒武纪的海相石油和烃源岩中的C29甾烷优势只能来源于海洋藻类。陈致林等[26]发现宏体褐藻C29甾醇优势,宏体绿藻和宏体红藻C27甾醇占优势,而宏体褐藻生活在深水区[27]。GRANTHAM等[20]认为古生代和前寒武纪的海相原油和烃源岩中的C29甾烷来自浮游绿藻,而现生浮游绿藻的确具有C29甾醇优势[28,29]。甾烷的不同优势组合可能代表了不同的生物来源,也代表了不同的水深。C27甾烷优势代表浅海环境,C29甾烷优势可能代表河口或者深海环境,强烈的C29甾烷优势则代表了远岸深水环境[30]。来自塔里木盆地寒武-奥陶系烃源岩的海相原油中亦存在高丰度的C29规则甾烷。由于甾醇(成岩转化成甾烷类)在地质时期是稳定的,它们可以起到“分子化石”的作用,甚至在没有实体化石保存的情况下记录生物体的进化[31]。所以对塔里木盆地海相原油中高丰度C29规则甾烷的来源进行研究是一项必要且有意义的工作。
1 地质背景
塔里木盆地是中国陆上最大的含油气叠合盆地 (面积达 56×104km2,见图1),其区域地质具有多盆地原型、多期构造运动及多期调整再分配等特点[32]。多年勘探实践表明,塔里木盆地油气储量巨大,具有多生油层系、多油气源区、长期且多期生烃及多期聚集等特点[33,34]。塔里木盆地海相原油主要聚集于以下3个油气区: 雅克拉-塔北-哈德逊构造带、塔中构造带及巴士托-和田河构造带[35]。塔里木盆地寒武系烃源岩平面上分布范围广,其有机相呈现明显的分异:东部为欠补偿盆地有机相、中部为开阔台地和局限台地相,而西部为蒸发潟湖有机相,高有机质丰度的烃源岩主要分布于台盆区的东部和西部。盆地东部寒武系欠补偿盆地相,高有机质丰度烃源岩厚度较薄、分布广泛;盆地中部寒武系开阔台地和局限台地相,碳酸盐岩地层较厚;在盆地西部下、中寒武统烃源岩主要为蒸发潟湖相烃源岩[36]。
塔中地区晚奥陶世“台地-斜坡-盆地”沉积体系发育,受构造作用、海平面升降以及气候、物源、水深、地形、水动力等环境因素的控制,桑塔木组为灰-灰黑色钙质泥岩;良里塔格组沉积早中期(良三段~良五段)主要为厚层灰色-深灰色泥晶灰岩;良里塔格组沉积晚期(良一段~良二段)主要为生物灰岩和粒屑灰岩。中、下奥陶统经历长时间的风化剥蚀,不同区带残留地层岩性差异较大。塔中井区残留鹰山组二段及其以下地层,自上而下发育泥晶灰岩、泥晶生屑、砂屑灰岩、含云灰岩。
很多地球化学家都对塔里木盆地台盆区的海相原油的油源问题进行了相关的研究,并提出了很多颇有见地的观点。HANSON等[37]利用甾烷与甲基三芳甾烷的分布特征进行油源对比研究认为,塔东2井和塔中62井的原油主要来自寒武系烃源岩;孙永革等[38]在轮南原油和沥青裂解产物中发现芳基类异戊二烯烃以及异常丰富的四甲基苯,并推断轮南原油有寒武系烃源岩的贡献。张水昌等[39,40]则通过萜烷参数进行油源对比研究后认为,塔里木盆地海相原油除了少数来自寒武系烃源岩外,主要来自中上奥陶统烃源岩;杨福林等[41]的研究则认为,塔里木盆地寒武系烃源岩与寒武系储层中的原油具有较好的亲缘性,而与奥陶系储层中的原油没有亲缘关系。现今的观点普遍趋同于这些海相油来源于寒武-奥陶系烃源岩[42],下古生界海相原油异常C29甾烷的分布特征和意义都值得深入研究。
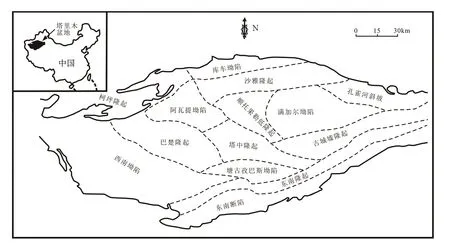
图1 塔里木盆地构造简图(据文献[38,41])Fig.1 Map showing the location of the Tarim Basin (quoted from references[38,41])
2 样品和试验
该研究所用61个原油样品主要采自塔里木盆地塔中和塔北地区的奥陶系储层,个别原油样品采自志留系和石炭系储层,所有样品均源自寒武-奥陶系烃源岩。对原油样品的饱和烃和芳烃进行了色谱质谱分析(GC-MS)。GC-MS分析条件为:Agilent 6890N/5995MSD气相色谱-质谱联用仪。色谱柱为HP5-MS石英弹性毛细柱(30m×0.25mm×0.25μm.i.d.)。升温程序:初始温度为50℃,恒温2min后,从50℃至100℃的升温速率为20℃/min,再以3℃/min的速率升温至310℃,维持恒温15min。以脉冲不分流方式进样,进样器温度300℃,载气为氦气,流速为1mL/min,电离能量为70eV,检测方式为全扫描(SCAN)/多离子检测(SIN)。试验获得的结果与参数如表1所示。
3 结果与讨论
图2为研究区评价原油成熟度的各参数之间的相关关系图。图2(a)是C29规则甾烷异构化成熟度参数判别图版[43],从图2(a)可见,研究区的原油主要处于成熟-高成熟阶段。图2(b)显示原油成熟度参数甲基菲比值(F1)与二甲基萘比值(DNR)正相关,同时图2(c)显示F1与4-/1-甲基二苯并噻吩(4-/1-MDBT)也正相关,可见F1是比较合理的成熟度参数。此外,图2(d)显示F1与C29规则甾烷20S/(20S+20R)负相关,表明原油已经达到成熟,C29规则甾烷20S/(20S+20R)开始发生倒转。由于F1与其他成熟度参数有较好的相关性(见图2),因此采用F1来表征研究区原油的热演化程度。

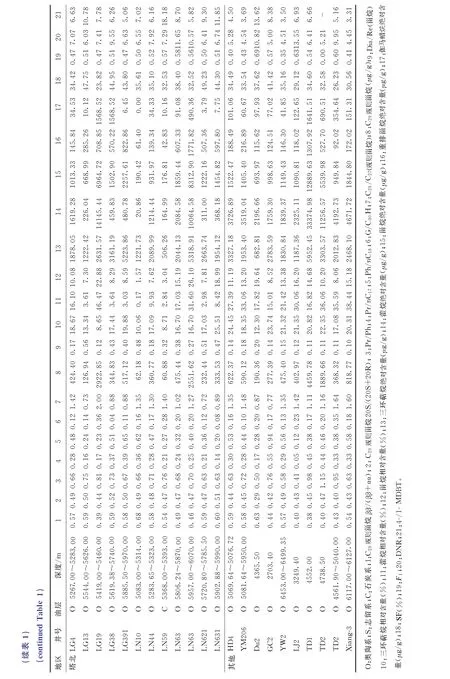
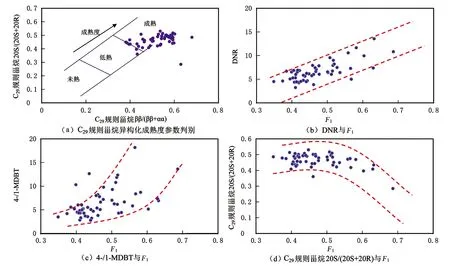
图2 研究区原油成熟度各参数之间的相关关系图(图2(a)图版据文献[43])Fig.2 Correlation diagrams of relevant maturity parameters of crude oil in the study area (quoted from reference[43])
3.1 链烷烃
链烷烃主要包括正构烷烃系列和异构烷烃系列。图3(a)为TZ58号油样的总离子流图,图3(b)为其异构烷烃质量色谱图,图3(c)为姥鲛烷与C17正构烷烃比值(Pr/nC17)和植烷与C18正构烷烃比值(Ph/nC18)的相关关系图。研究区原油样品的正构烷烃以低碳数(C14~C23)烷烃为主,呈前锋型分布,主峰碳为C14~C17(见图3(a))。原油的正构烷烃系列均不具奇偶优势,其正构烷烃碳优势分布指数(CPI)和奇偶优势指数(OEP)均已趋近1,显示出成熟原油的特征。样品中存在丰富的异构烷烃,异构烷烃系列化合物中2-和3-甲基支链占优势,异构烷烃的碳数范围与正构烷烃相当(iC13~iC32),亦呈现出前锋型分布,主峰碳为iC15(见图3(b))。高丰度的低碳数异构烷烃通常代表蓝细菌和藻类的输入[44-47]。Pr/Ph在0.4~1.2之间,主要集中在0.6~0.8,Pr/nC17在0.14~0.94之间,Ph/nC18在0.13~5.37之间。Pr/nC17与Ph/nC18的相关关系图显示研究区原油是典型的在还原环境形成的海相油(见图3(c))。
图3 研究区原油正构烷烃和异构烷烃分布特征及Pr/nC17与Ph/nC18相关关系图(图版引自文献[43])Fig.3 Distribution characteristics of n-alkanes and isomers alkanes of crude oil in the study area and correlation diagrams of Pr/nC17 and Ph/nC18 (quoted from reference[43])
3.2 规则甾烷
根据C29/C27(规则甾烷)的数据分布(见表1),可以将研究样品分为3类:C29/C27(规则甾烷)<1,1
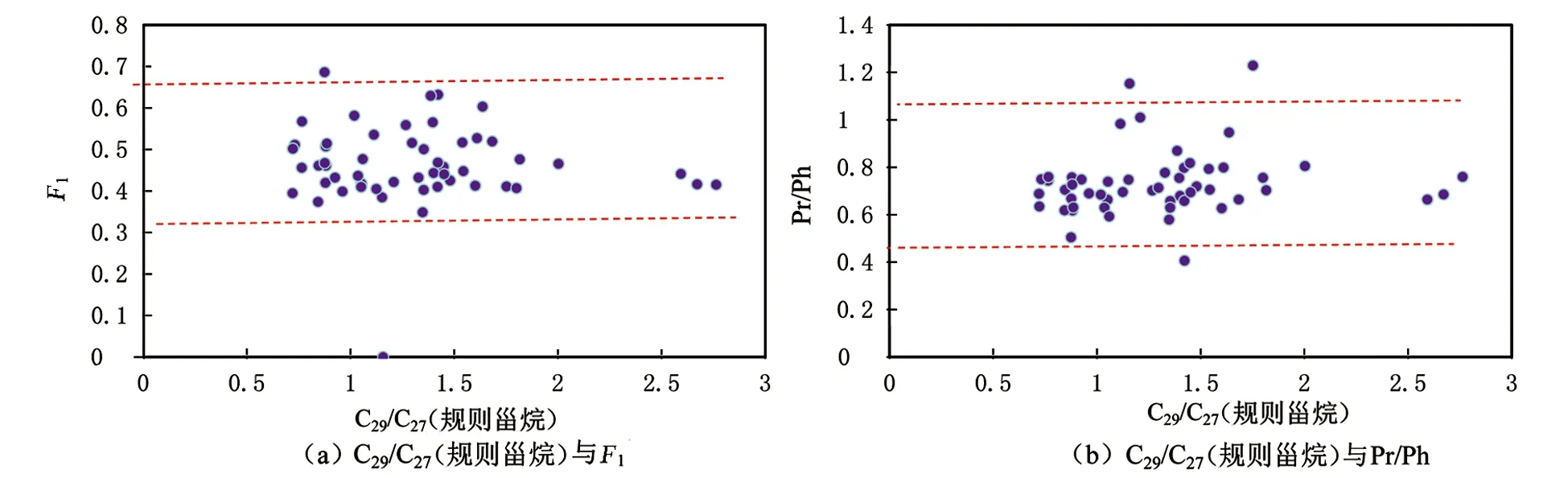
图4 C29/C27(规则甾烷)与F1及Pr/Ph的相关关系图Fig.4 Correlation diagrams of C29/C27 (regular sterane) with F1 and Pr/Ph
图5是研究区3类典型原油样品的甾烷和萜烷分布图,其中图5(a)是C29/C27(规则甾烷)<1型原油的甾烷分布图;图5(b)是C29/C27(规则甾烷)<1型原油的萜烷分布图;图5(c)是1
图6为研究区样品中甾烷、藿烷以及三环萜烷相对含量和绝对含量与C29/C27(规则甾烷)和C29规则甾烷绝对含量的相关关系图。其中图6(a)为C29/C27(规则甾烷)与甾烷相对含量的相关关系图;图6(b)为C29规则甾烷绝对含量与甾烷绝对含量的相关关系图。图6(c)为C29/C27(规则甾烷)与藿烷相对含量的相关关系图;图6(d)为C29规则甾烷绝对含量与藿烷绝对含量的相关关系图。图6(e)为C29/C27(规则甾烷)与三环萜烷烷相对含量的相关关系图;图6(f)为C29规则甾烷绝对含量与三环萜烷绝对含量的相关关系图。由图6可知,C29/C27(规则甾烷)与甾烷相对含量正相关(见图6(a))、C29规则甾烷绝对含量与藿烷绝对含量具有极好的正相关关系(见图6(b),R2=0.942);C29/C27(规则甾烷)与藿烷相对含量正相关(见图6(c)),C29规则甾烷绝对含量与藿烷绝对含量也有好的正相关关系(见图6(d),R2=0.886)。而C29/C27(规则甾烷)与三环萜烷相对含量(见图6(a))、C29规则甾烷绝对含量与三环萜烷绝对含量的相关性都较差(见图6(f))。从相对含量与绝对含量2个方面的分析结果来看,C29甾烷的成因与甾烷和藿烷的生物来源有关,而与三环萜烷的生物来源关系不大。通常,人们认为藿烷来自细菌[48],甾烷类化合物一般由真核生物细胞壁中的甾醇、甾酮以及甾酸等转化而成[49],在一些藻类为主要原始生物母质的烃源岩中三环萜烷丰度高,细菌来源的有机质三环萜烷丰度低;且高丰度的三环萜烷通常与藻类有关[50]。高丰度的C29规则甾烷与甾烷和藿烷的相关性比三环萜烷好,表明C29规则甾烷的生物母质除了藻类可能还有其他的来源。
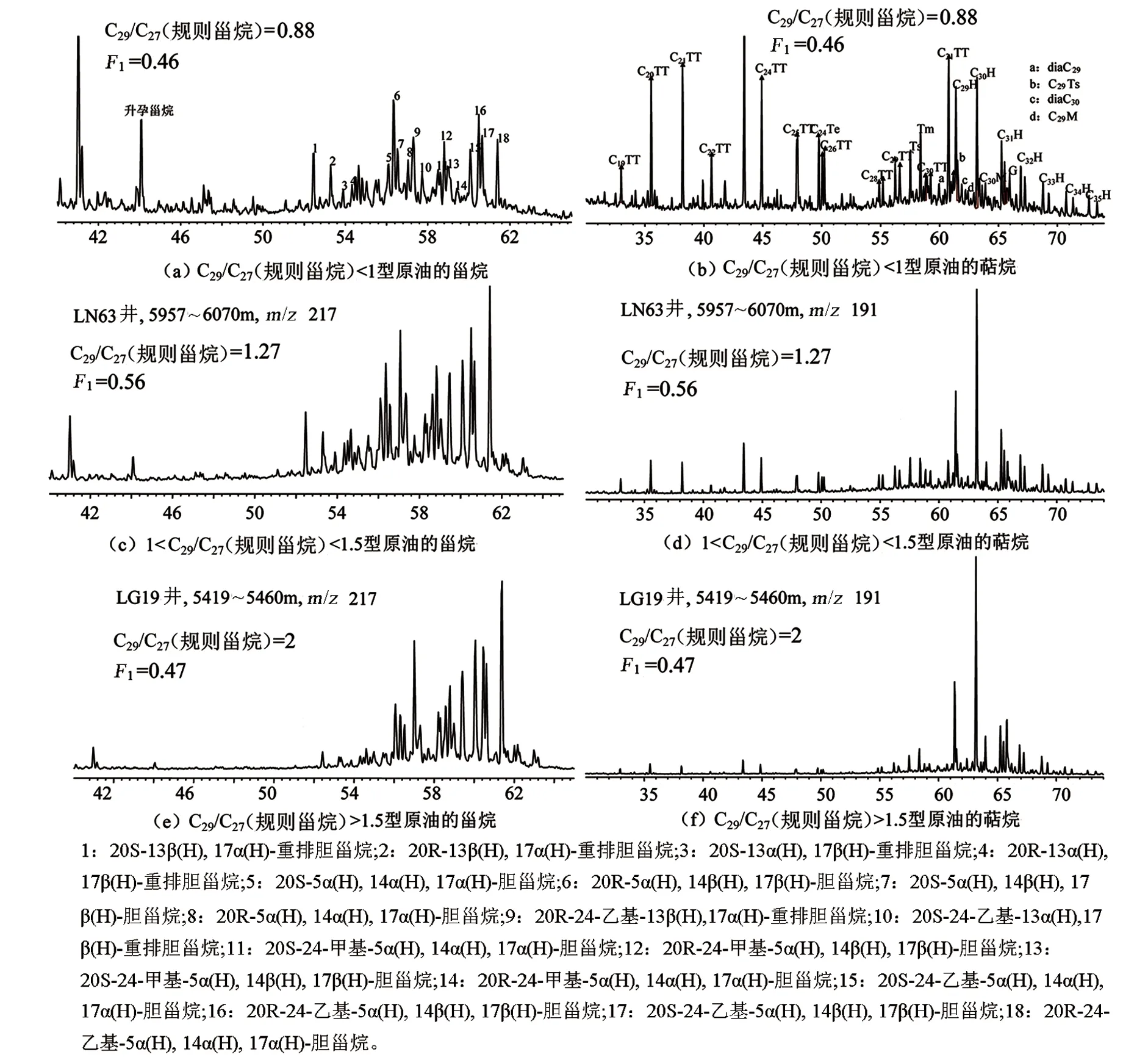
图5 研究区典型原油样品的甾烷和萜烷分布特征Fig.5 Distribution characteristics of sterane and terpenes in typical crude oil samples from the study area
3.3 重排甾烷
图7为规则甾烷各异构化参数及重排甾烷参数与C29/C27(规则甾烷)、C29规则甾烷绝对含量的相关关系图。图7(a)中重排甾烷与规则甾烷的比值(Dia/Re(甾烷))与C29/C27(规则甾烷)呈负相关关系;图7(b)中C29规则甾烷异构化比值(C29规则甾烷αββ20S/ααα20R)与C29/C27(规则甾烷)相关性很差;图7(c)中C28规则甾烷异构化比值(C28规则甾烷αββ20S/ααα20R)与C29/C27(规则甾烷)相关性很差;图7(d)中C27规则甾烷异构化比值(C27规则甾烷αββ20S/ααα20R)与C29/C27(规则甾烷)相关性很差。Dia/Re(甾烷)与C29/C27(规则甾烷)负相关,表明重排甾烷与C29规则甾烷可能在成因上有联系(见图7(a))。而C27、C28、C29规则甾烷各自的异构化参数C27规则甾烷αββ20S/ααα20R、C28规则甾烷αββ20S/ααα20R及C29规则甾烷αββ20S/ααα20R均与C29/C27(规则甾烷)无明显相关关系(图7(b)、(c)、(d))。图8为Dia/Re(甾烷)与成熟度参数F1以及环境氧化还原参数Pr/Ph的相关关系图。图8(a)显示,Dia/Re(甾烷)与F1无相关性,但是图8(b)显示,高丰度的重排甾烷主要分布在Pr/Ph为0.7的位置。由此可见重排甾烷的形成受环境氧化还原性的影响[51,52]。
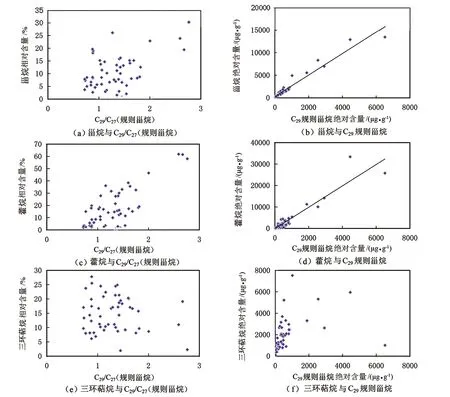
图6 研究区原油甾烷、藿烷、三环萜烷相对含量和绝对含量 与C29/C27(规则甾烷)、C29规则甾烷的相关关系图Fig.6 Diagrams of correlation between relative and absolute contents steranes,hotanes and tricyclic terpanes in cruid oil and C29/C27 (regular sterane) and C29 regular sterane in the study area
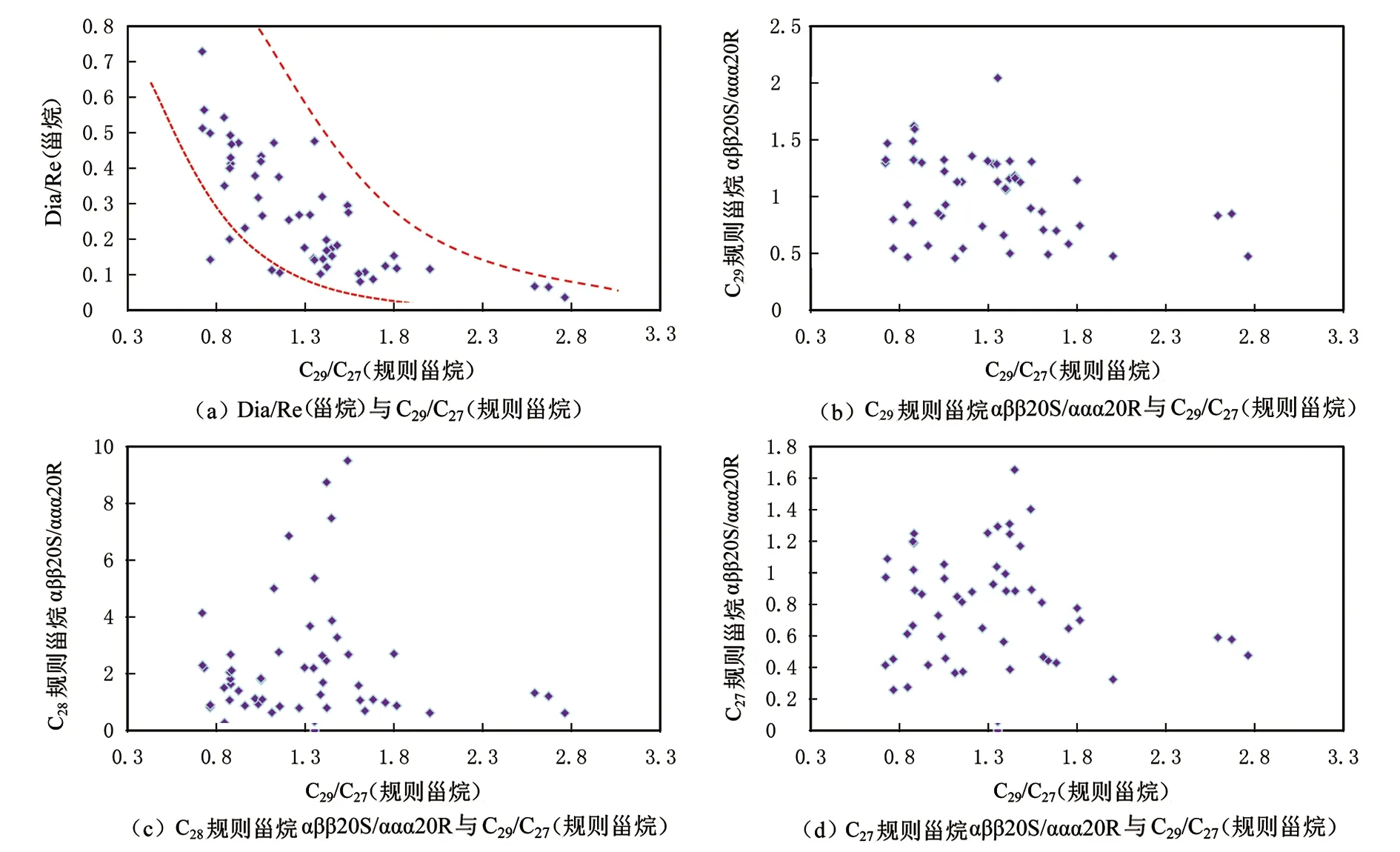
图7 研究区原油样品中甾烷异构化参数及重排甾烷参数与C29/C27(规则甾烷)的相关关系图Fig.7 Correlation diagrams of sterane isomerization parameters and diasterane with C29/C27(regular sterane) of crude oil samples in the study area
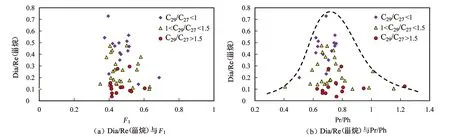
图8 研究区Dia/Re(甾烷)与F1和Pr/Ph的相关关系图Fig.8 Correlation diagrams of Dia/Re(sterane) with F1 and Pr/Ph in the study area
图9(a)为Dia/Re(甾烷)(重排甾烷与规则甾烷的比值)与藿烷相对含量相对含量的相关关系图,图9(b)为重排甾烷绝对含量与藿烷绝对含量的相关关系图,图9(c)为Dia/Re(甾烷)及与甾烷相对含量的相关关系图,图9(d)为重排甾烷绝对含量与甾烷绝对含量的相关关系图,图9(e)为Dia/Re(甾烷)与三环萜烷与藿烷的相对含量比值(TT/H)的相关关系图,图9(f)为重排甾烷绝对含量与三环萜烷绝对含量的相关关系图。研究区样品中Dia/Re(甾烷)与甾烷和藿烷的相对含量负相关(见图9(a)、(c)),与TT/H正相关(见图9(e))。此外,重排甾烷的绝对含量与藿烷和甾烷的绝对含量的相关性较差(见图9(b)、(d)),而与三环萜烷绝对含量具有较好的相关性(见图9(f),R2=0.655)。从相对含量与绝对含量的分析结果来看,重排甾烷的成因与三环萜烷的生物来源的相关性更大。综合上述分析,重排甾烷的成因既受环境的影响,又受生源的影响。
3.4 伽马蜡烷
图10为研究区原油样品中伽马蜡烷相关参数分别与C29规则甾烷、重排甾烷相关参数及F1、Pr/Ph的相关关系图。图10(a)为伽马蜡烷/C30藿烷(G/C30H)与C29/C27(规则甾烷)的相关关系图,图10(b)为G/C30H与Dia/Re(甾烷)的相关关系图,图10(c)为伽马蜡烷绝对含量与C29规则甾烷绝对含量的相关关系图,图10(d)为伽马蜡烷绝对含量与重排甾烷绝对含量的相关关系图,图10(e)为G/C30H与F1的相关关系图,图10(f)为G/C30H与Pr/Ph的相关关系图。G/C30H与C29/C27(规则甾烷)正相关而与Dia/Re(甾烷)负相关(见图10(a)、(b)),且G/C30H与F1和Pr/Ph无明显相关性(见图10(e)、(f))。此外,伽马蜡烷绝对含量与C29规则甾烷绝对含量有较好的正相关关系(见图10(c),R2=0.794),而与重排甾烷绝对含量则没有相关性(见图10(d))。结果表明,伽马蜡烷丰度的受控因素可能与C29规则甾烷类似,主要是受生物母质来源的影响,而热演化和沉积环境的影响较小。
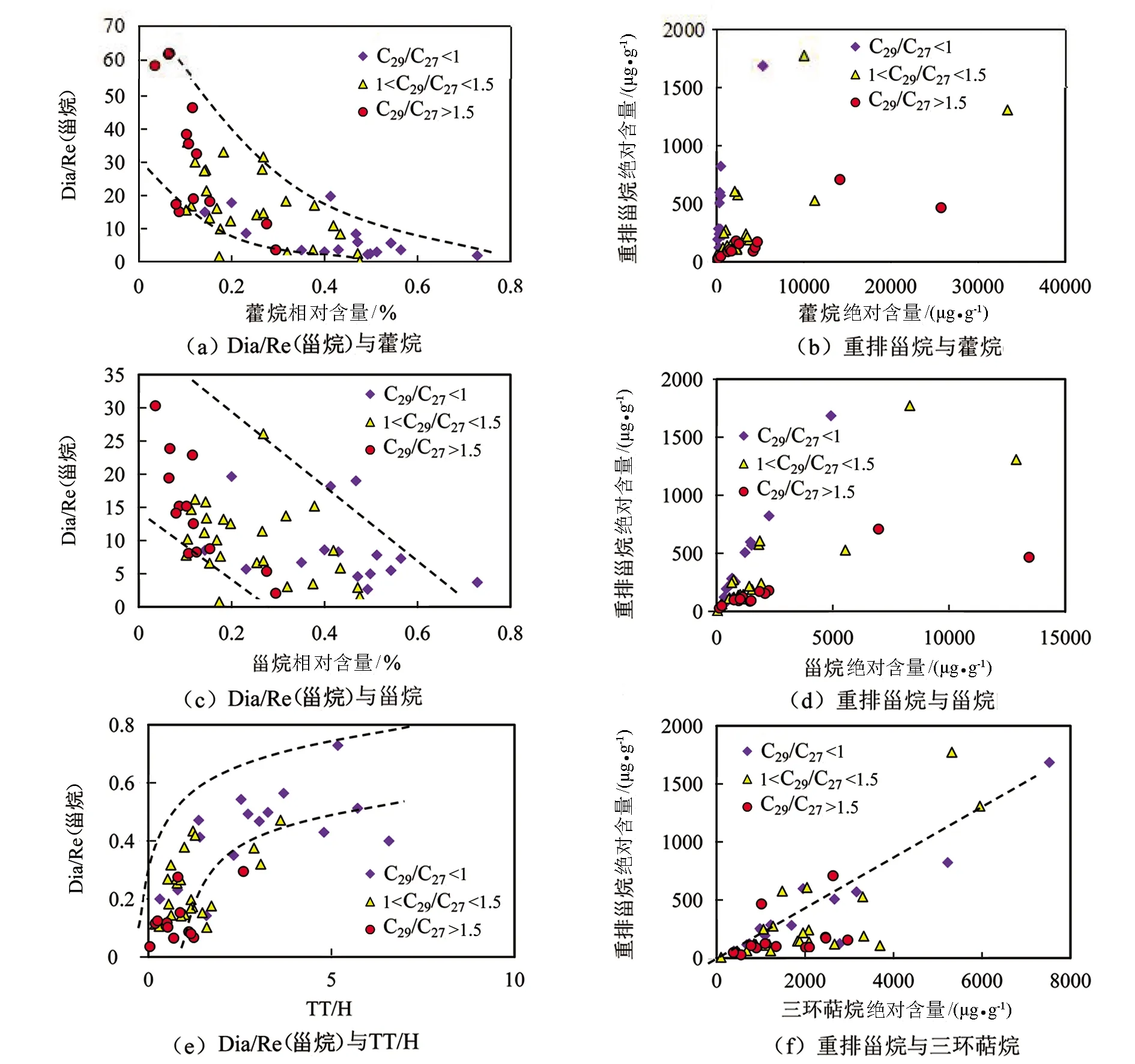
图9 研究区原油中重排甾烷相关参数与藿烷、甾烷、三环萜烷相对含量和绝对含量的相关关系图Fig.9 Correlation diagrams of relative and absolute contents of diasteranes with steranes,hopanes and tricyclic terpanes in the study area
图11为研究区原油样品中伽马蜡烷相关参数分别与甾烷、藿烷及三环萜烷相对含量和绝对含量的相关关系图。图11(a)为G/C30H与甾烷相对含量的相关关系图,图11(b)为伽马蜡烷绝对含量与甾烷绝对含量的相关关系图,图11(c)为G/C30H与藿烷相对含量的相关关系图,图11(d)为伽马蜡烷绝对含量与藿烷绝对含量的相关关系图,图11(e)为G/C30H与三环萜烷相对含量的相关关系图,图11(f)为伽马蜡烷绝对含量与三环萜烷绝对含量的相关关系图。由图11可知,G/C30H与甾烷相对含量、藿烷相对含量正相关(见图11(a)、(c)),而与三环萜烷相对含量无相关关系(见图11(e))。与此同时,伽马蜡烷绝对含量与甾烷绝对含量、藿烷绝对含量正相关(见图11(b)、(d)),而与三环萜烷绝对含量无相关性(见图11(f))。且甾烷绝对含量与伽马蜡烷绝对含量的相关性(见图11(b),R2=0.7079)比藿烷绝对含量与伽马蜡烷绝对含量的相关性(见图11(d),R2=0.6851)要好。综合分析表明,伽马蜡烷的来源与甾烷和藿烷的母质来源有关,而与三环萜的母质来源无关,且与C29规则甾烷的母质来源一致。
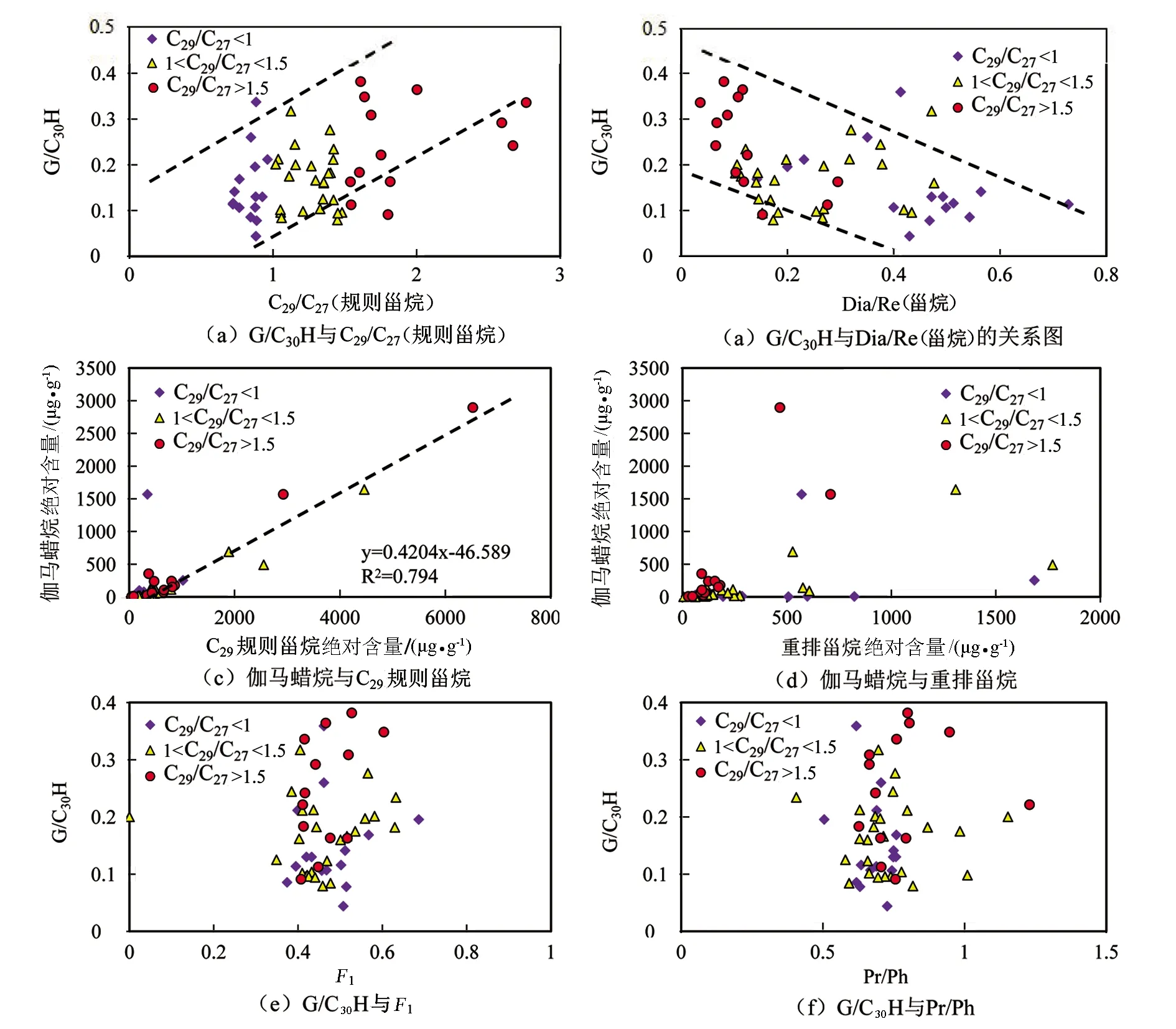
图10 研究区原油中伽马蜡烷相关参数与C29规则甾烷、重排甾烷相关参数及F1、Pr/Ph的相关关系图Fig.10 Correlation diagrams of the relative parameters of Gammacerane with that of C29 regular steranes, rearranged steranes and F1,Pr/Ph of crude oil in the study area
3.5 C29规则甾烷与生物来源及水体环境的关系
图12为硫芴相对含量(简称SF)分别与C29/C27(规则甾烷)及G/C30H的相关关系图。图12(a)显示C29/C27(规则甾烷)与SF负相关,图12(b)显示G/C30H也与SF负相关。C29/C27(规则甾烷)<1的样品其SF分布在18.04%~32.47%之间,1< C29/C27(规则甾烷)<1.5的样品其SF分布在32.53%~40.35%之间,C29/C27(规则甾烷)>1.5的样品其SF分布在40.1%~49.4%之间(见表1)。因为该研究中的样品采自同一地区和同一时代,其SF可以表征水体的硫化程度,C29与伽马蜡烷的丰度均受生物母质来源的控制。
图13为甾烷、藿烷及三环萜烷的相对含量及绝对含量与SF相关关系图。图13(a)中甾烷相对含量与SF为负相关关系;图13(b)中甾烷绝对含量与SF也为负相关关系;图13(c)中藿烷相对含量与SF为负相关关系;图13(d)中藿烷绝对含量与SF也为负相关关系。不同之处在于,图13(e)中三环萜烷相对含量与SF为正相关关系;图13(f)中三环萜烷绝对含量与SF也为正相关关系。分析表明,随着SF相对含量的增加即水体硫化程度的增加,甾烷的相对含量和绝对含量均降低(见图13(a)、(b)),藿烷的相对含量和绝对含量也降低(见图13(c)、(d)),而三环萜烷的相对含量与绝对含量均增大(见图13(e)、(f))。也就是说随着水体硫化程度的增加,利于产生三环萜烷的藻类生存,但是不利于易产生高丰度C29规则甾烷的生物生存,同时,这种产出高丰度C29规则甾烷的生物也产生伽马蜡烷。
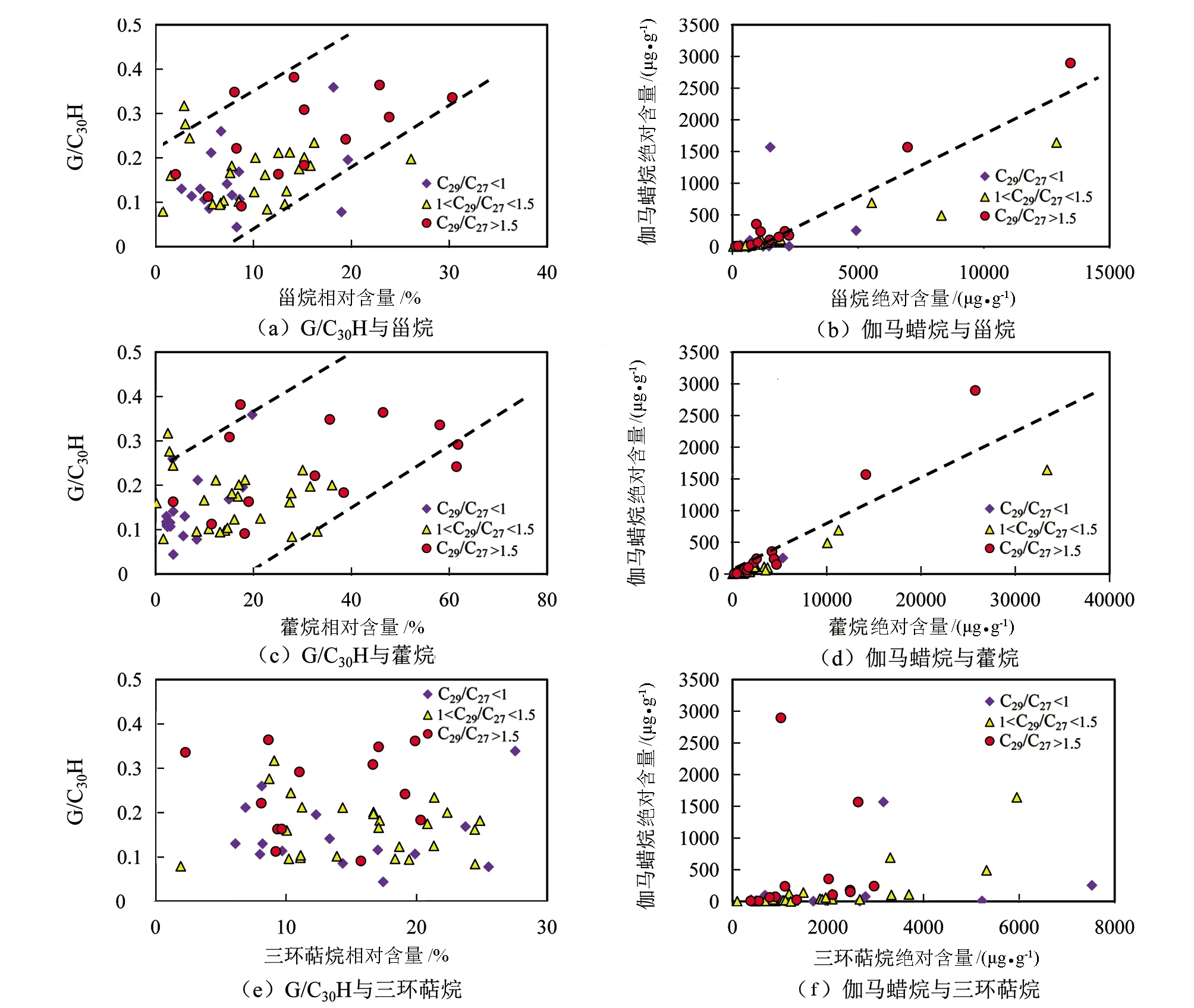
图11 伽马蜡烷相关参数与甾烷、藿烷及三环萜烷相对含量和绝对含量的相关关系图Fig.11 Correlation diagrams of the relative parameters of gammacerane with the relative and absolute contents of sterane,hopane and tricyclic terpene
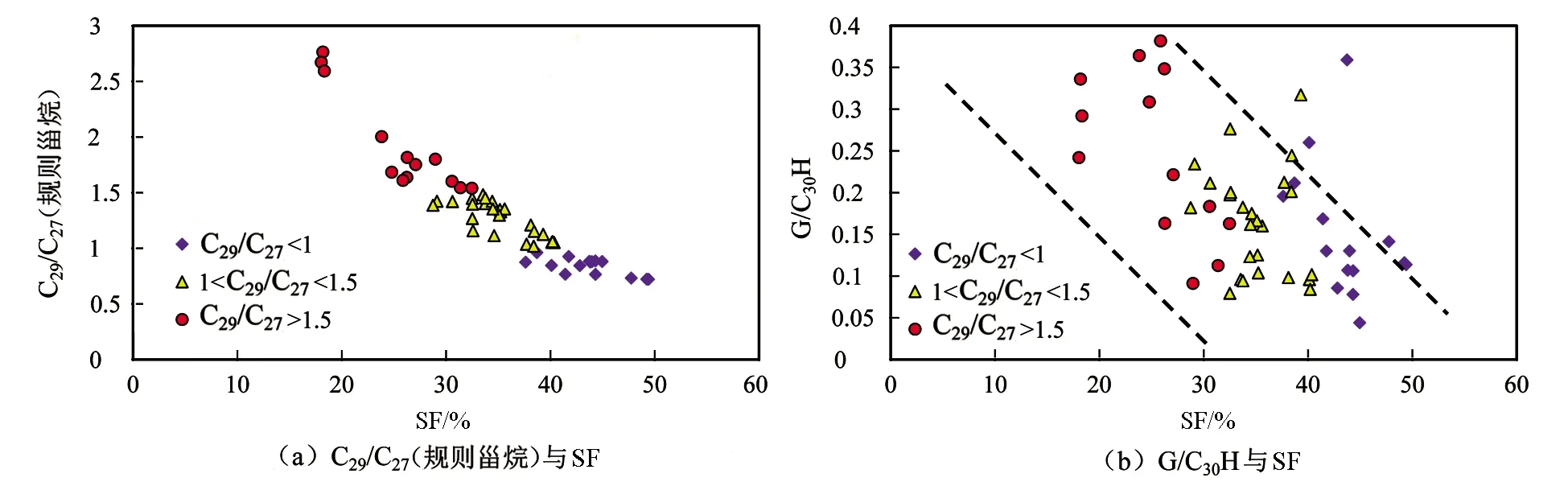
图12 SF与C29/C27(规则甾烷)及G/C30H的相关关系图Fig.12 correlation diagrams of SF with C29/C27(regular steranes) and G/C30H
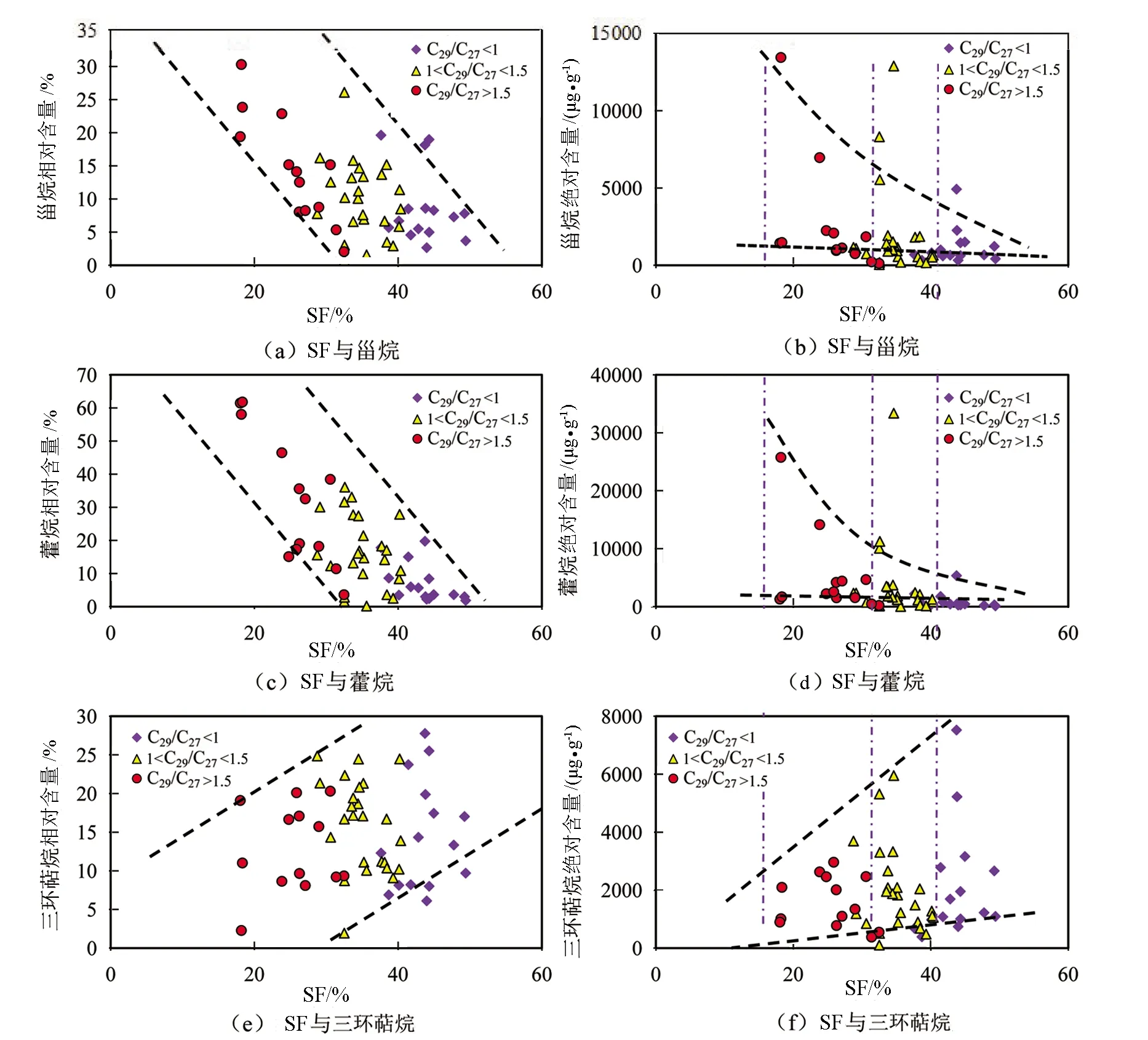
图13 甾烷、藿烷和三环萜烷的相对含量及绝对含量与SF的相关关系图Fig.13 Correlation diagrams of the relative and absolute contents of hopanes, steranes and tricyclic terpenes with SF
图14为Dia/Re(甾烷)和重排甾烷绝对含量与SF的相关关系图。
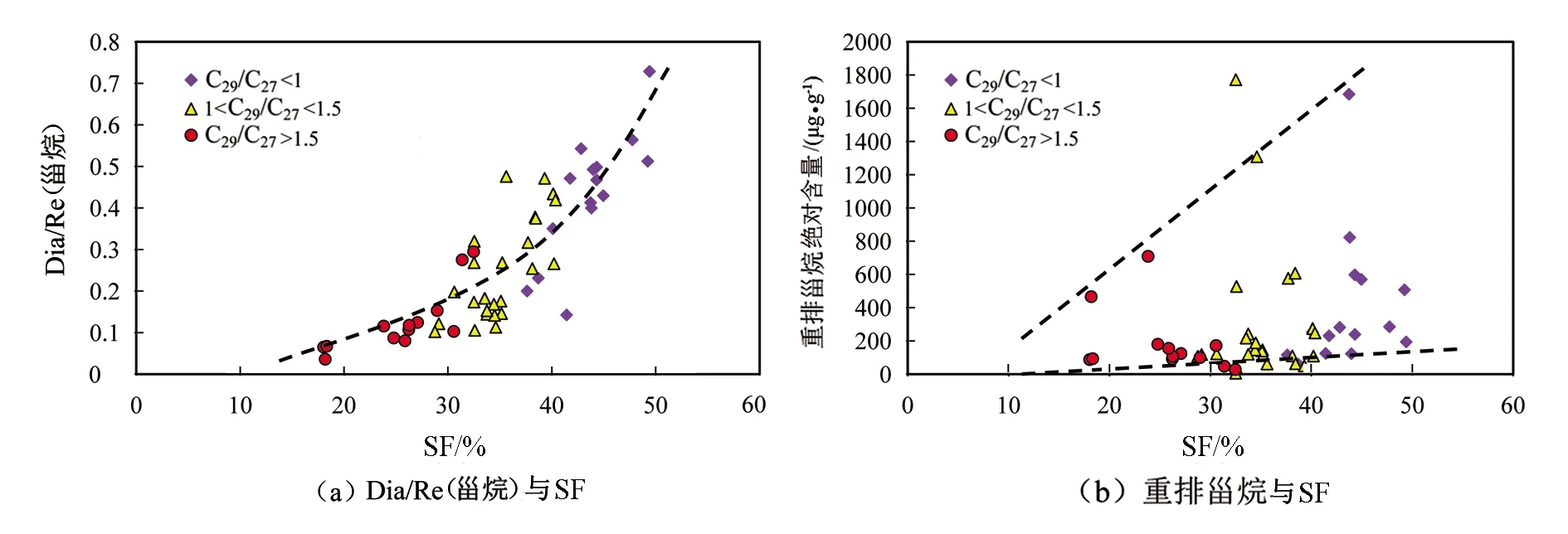
图14 Dia/Re(甾烷)及重排甾烷绝对含量与SF的相关关系图Fig.14 Correlation diagrams of Dia/Re(sterane) and the absolute contents of rearranged sterane with SF
图14(a)为Dia/Re(甾烷)与SF的相关关系图;图14(b)为重排甾烷绝对含量和SF的相关关系图。通常水体的硫化程度越大则其还原性越强,但是Dia/Re(甾烷)以及重排甾烷绝对含量均与SF表现出正相关关系(见图14(a)、(b))。这与三环萜烷绝对含量与SF的相关性相似(见图13(f))。通常弱氧化的环境有利于重排的发生,但是代表强还原的硫化水体中重排甾烷的含量反而更高,从另一方面验证了该研究中的重排主要受生物来源的控制。
4 讨论
细菌通常不生成甾醇,对沉积物中甾烷的碳数变化无明显的影响。细菌甾醇合成只能到鲨烯阶段,但是有些细菌会利用甾醇作为电子受体[53],因此细菌所需的甾醇只能从环境中获取[54]。光合自养的蓝细菌也几乎不生产甾醇,以蓝藻为主要初级生产者的湖泊为例,以浮游植物为食的水溞(Daphnia)对蓝细菌的碳转化效率非常低,主要是因为蓝藻含甾醇非常少,水溞在取食蓝藻之后不能获得足够其生理活动所需的甾醇,因而影响其生长繁殖[55]。最终,当蓝藻占浮游植物的80%以上时,水溞不能有效控制蓝藻的生物量,为了治理蓝藻,需要人工投放甾醇养料。动物来源的甾醇多为C27甾醇,但对甾核和/或侧链的修饰允许多样性的结构,通常为C26~C31甾醇。BERENBERG在用藻类Thalassiosirapseudonana(海链藻属,海链藻)和Isochrysissp(等鞭金藻属)对牡蛎进行喂养之后,研究牡蛎与藻类体内的甾醇组成,结果显示牡蛎体内存在的甾醇种类更多,绝对含量也更高,其中24-乙基胆甾-5,22 -二烯醇(24-Ethylcholesta-5,22-dienol)、24-去胆甾-5,22二烯醇(24-Norcholesta-5,22-dienol)和22-脱氢胆固醇(22-Dehydrocholesterol)只在牡蛎体内检测到[56]。24-脱氢胆固醇(Desmosterol)只在被喂养了藻类的牡蛎中检测出(即藻类和未被喂食藻类的牡蛎体内未检出)。表明牡蛎可能存在甾醇的生物转化,重新合成,或选择性摄取。这表明动物也是C29甾醇的重要来源。来自海绵的24-异丙基胆甾烷是6.50~5.40亿年(新元古代至早寒武世)岩石中一种丰富的甾烷[57],被认为是最古老的动物标志化合物。GOLD等[58]通过对海绵和藻类产生甾醇甲基转移酶(SMT)的基因进行了研究,遗传数据表明,藻类植物和海绵分别独立地进化出利用SMT复制生产24-异丙基胆甾醇的能力。用分子钟的方法证实,海绵SMT复制事件与24-异丙基胆甾烷的出现在新元古代重叠,但藻类SMT复制事件发生在显生宙,比海绵晚了1亿年,说明动物可能比藻类更早具备生产高碳数甾醇的能力。该研究各项参数表明,高丰度的C29规则甾烷与藿烷及伽马蜡烷的生物母质有关,与三环萜烷的母质藻类无关,因此,可以推测C29规则甾烷可能来源于以细菌为食的浮游动物。
大量来自寒武纪地层的 Fe-S-C 化学系统数据、微量元素数据、C-N-Mo 同位素和黄铁矿形态学等证据则表明,深部海洋可能依旧以还原水体为主且缺氧硫化的水体广泛分布[59-65]。此外,华南扬子地台寒武纪早期地层水体化学空间对比表明,华南这一时期的水体化学可能与晚新元古代相似,仍旧是高度动态分层的,陆架中等水深的硫化水体可与表层氧化和深部铁化水体长期保持动态共存[66-68]。同一地理位置的硫化水体范围是动态变化的,这种变化直接影响了这一地区的生物种群分布。浮游动物不适宜在硫化水体中生存,随着水体硫化程度增加,其生物量降低,C29规则甾烷和伽马蜡烷的相对含量和绝对含量都降低;而藻类则对硫化水体有较强的适应能力,由于捕食者的减少,藻类的生物量反而随水体的硫化程度增加而增加,因此三环萜烷和重排甾烷的相对含量和绝对含量也随着增加。
5 结论
1)C29/C27(规则甾烷)与成熟度参数甲基菲比值(F1)以及沉积环境参数Pr/Ph无明显相关性,但是与甾烷、藿烷的相对含量正相关,与三环萜烷的相对含量无关;此外,C29规则甾烷的绝对含量与甾烷和藿烷的绝对含量正相关,与三环萜烷的绝对含量没有相关性,表明高丰度的C29规则甾烷可能主要是受生物母质来源影响。C29/C27(规则甾烷)与C27、C28、C29规则甾烷的异构化比值没有相关性,但是与Dia/Re(甾烷)负相关。Dia/Re(甾烷)也与成熟度参数F1无关,当Pr/Ph为0.7时存在高丰度的重排甾烷分布。C29/C27(规则甾烷)与三环萜的相对含量正相关,与甾烷、藿烷的相对含量相关;此外,C29规则甾烷的绝对含量与三环萜烷的绝对含量有较好的正相关性,与甾烷和藿烷的绝对含量的相关性较差,表明高丰度的重排甾烷可能受控于生物母质来源和沉积环境的影响。
2)C29/C27(规则甾烷)与G/C30H正相关,Dia/Re(甾烷)与G/C30H负相关,C29规则甾烷的绝对含量与伽马蜡烷的绝对含量有好的正相关关系,而重排甾烷的绝对含量与伽马蜡烷的绝对含量相关性较差。此外伽马蜡烷的相关参数与成熟度参数F1以及沉积环境参数Pr/Ph无明显相关性,但是与甾烷、藿烷、三环萜烷的相对含量以及绝对含量的相关性与C29规则甾烷一致,表明伽马蜡烷的生物来源可能与高丰度的C29规则甾烷相同。C29/C27(规则甾烷)与SF负相关,而Dia/Re(甾烷)与SF正相关,且随着SF的变化,甾烷与藿烷的相对含量与SF负相关。三环萜烷的绝对含量与SF相关性不明显,三环萜烷的相对含量和绝对含量均与SF有较好的正相关性,表明水体的硫化程度是控制C29规则甾烷和重排甾烷的一个重要因素。
3)高丰度的C29规则甾烷和伽马蜡烷来自浮游动物,而重排甾烷则来自藻类。下古生代动态分层的硫化水体直接影响了生物种群的发育,当硫化水体收缩时,生物繁盛,细菌和浮游动物生物量增多,因此,藿烷、甾烷、伽马蜡烷的相对含量和绝对含量都增加。当硫化水体扩张时,海洋生物都受到抑制,但是,细菌和浮游动物受到的影响可能更显著。由于竞争者和捕食者的减少,藻类的生物量反而增大。因此,三环萜烷的相对含量和绝对含量随之增大。藻类来源的甾烷更易发生重排,因此,Dia/Re(甾烷)上升。
