嵊泗列岛海域三种贻贝贝体框架特征的差异*
2014-03-19白晓倩邹李昶任夙艺刘达博刘祖毅王志铮
白晓倩 杨 阳 邹李昶, 任夙艺 刘达博 刘祖毅 王志铮①
(1. 浙江海洋学院 舟山 316022; 2. 余姚市水产技术推广中心 余姚 315400; 3. 嵊泗县海洋与渔业局 嵊泗 202450)
贝壳表型作为贝类外部形态信息的综合反映和种质遗传规定性的外在体现, 不仅可为贝类种类鉴别和生长性能评估提供证据支持, 也可为贝类品系辨析和种质遗传特性研究提供重要线索。因此, 利用多元统计方法探析决定目标养殖贝类贝体框架特征的变量组合, 对于提高贝类种间分类的精细化程度、指导地理种群形态量化标记的构建和种质变异程度评估具有重要的学术研究价值。
嵊泗列岛海域既是我国厚壳贻贝(Mytilus coruscus)的重要原产地, 也是浙江省贻贝养殖重点区域。研究发现, 同域分布或养殖的贻贝科种类间普遍存在杂交或渐渗(introgressive hybridization)现象(Andersonet al, 1938; McDonaldet al, 1988; Sarveret al, 1991,1993; Rawsonet al, 1996; Inoueet al, 1997; Comesanaet al, 1999; Hilbishet al, 2002; Toroet al, 2002; Carenet al, 2006); 近年来, 形态、出肉率和口感介于厚壳贻贝与紫贻贝(Mytilus edulis)之间的“杂交贻贝”在嵊山列岛贻贝养殖海区也时有采获(张义浩等, 2003),经查上述两种贻贝已在该养殖区域发生了杂交与基因渐渗(沈玉帮等, 2006)。因此, 从定量水平弄清厚壳贻贝、紫贻贝和“杂交贻贝”贝体框架特征间的本质差异, 无疑对于切实规避种间种质污染具现实意义,但迄今国内外有关贝类相近种间贝体框架特征差异的系统研究尚未见报道。鉴于此, 本文作者于2011年10月22日以人工培育的2+龄厚壳贻贝同生群养殖个体(野生亲贝源自非养殖海区, 稚贝出池时间为2009年6月、海区中间培育时间为2009年6月至2010年4月, 大规格苗种筏式养殖起始时间为2010年4月)、同域生长的1+龄紫贻贝同生群养殖个体(苗种源自大连海区)以及混于厚壳贻贝和紫贻贝养殖筏架中的“杂交贻贝”为实验对象, 采用多元统计方法开展了上述3种贻贝贝体框架特征的差异研究, 以期为嵊泗本地厚壳贻贝的种质保护和“杂交贻贝”杂种生长优势的利用提供基础资料。
1 材料与方法
1.1 样品来源
本研究所用样品均取自嵊泗县金盟海水养殖专业合作社在枸杞岛干斜村邻近海域(32°42’16’—32°42’38’N, 122°45’29’—122°45’53’E)的贻贝养殖筏架。
1.2 样品贝体框架性状参数值的测定
样品运回实验室后各随机选取其中壳形完整的112枚活体作为测定群体。测定样品经清除壳表附着物后, 用电子数显游标卡尺(精度0.02mm)测量壳长SL(壳前、后端间的最大水平距离)、壳宽SW(左右两壳紧密时的最大距离), 并在壳内面逐一编号保存备用; 采用扫描像素法(李宝光等, 2006)依次测量壳高SH (BD, 壳背、腹间的最大垂直距离)以及线段OA(壳顶至韧带末端的直线距离)、OB(壳顶至壳背缘最高点的直线距离)、OC(壳顶至壳后端最远点的直线距离)、OD(壳顶至壳高性状在腹缘的落点的直线距离)、AB(韧带末端至壳背缘最高点的直线距离)、BC(壳背缘最高点至壳后端最远点的直线距离)、CD(壳后端最远点至壳高性状在腹缘的落点的直线距离)等8个贝体框架性状。具体测量部位如图1所示。
1.3 分析方法
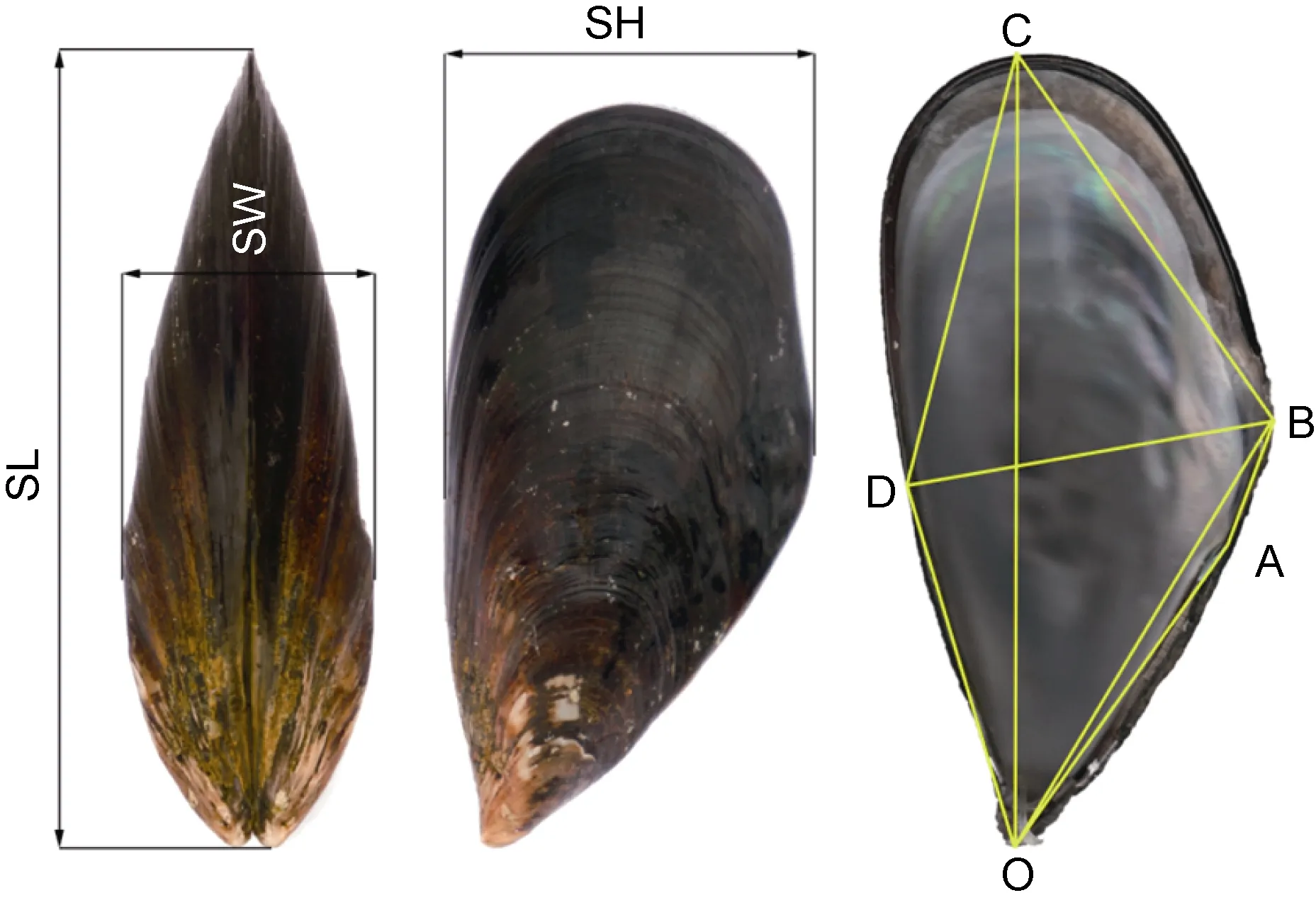
图1 实验贝贝体框架性状的测量部位Fig.1 The measurements of the morphological traits of Mytilus
整理所测结果, 计算三种实验贝各项贝体框架特征指标的均值、标准差和变异系数后, 采用LSD多重比较法检验组间差异显著性(P<0.05视为显著水平); 根据三种实验贝各项贝体框架特征指标均值计算欧氏距离, 并度量和评价三者间贝体形态特征的相似程度(P<0.05视为显著水平); 根据所有实验样本贝体框架特征指标的均值进行主成分分析和判别分析, 其中主成分分析以特征值大于1且累计贡献率大于80%为原则确定主成分(PC)的提取个数, 采用逐步导入剔除法进行判别分析, 并参照李思发等(2005)的方法计算判别准确率(P1为某种实验贝判别正确的个体数占该贝实测总个数的百分比,P2为某种实验贝判别正确的个数占判入该实验贝的总个数的百分比)和综合判别率(P为各实验贝判别正确的个数之和占实测总个数的百分比)验证所建判别方程组的可靠性。上述计算均借助SPSS 17.0软件来实现。
2 结果与分析
2.1 三种实验贝间贝体框架特征的差异比较
由表1可见, 三种实验贝在贝体框架特征上均存在一定程度的差异, 主要表现为: (1)L1、L3均呈紫贻贝>厚壳贻贝>“杂交贻贝”(P<0.05),L2、L4、L8和L9均呈“杂交贻贝”>紫贻贝>厚壳贻贝(P<0.05),L5、L7均呈“杂交贻贝”>紫贻贝≈厚壳贻贝,L6呈厚壳贻贝>紫贻贝>“杂交贻贝”(P<0.05), 即在本研究所涉9项贝体框架特征指标中, 厚壳贻贝与紫贻贝间有2项指标无差异(P>0.05), 两者相似性指数达22.22%, 而“杂交贻贝”与紫贻贝和厚壳贻贝的指标相似性指数则均为0; (2) 在本研究所涉9项贝体框架特征指标中,三种实验贝变异系数大于10%的指标均仅为1项, 其中厚壳贻贝和紫贻贝均为L7, 而“杂交贻贝”则为L3。综上可知, 三种实验贝在贝体框架特征上具良好的区分度, 尤以“杂交贻贝”为甚。
2.2 三种实验贝间贝体框架特征的相似程度比较
欧氏距离系指n维空间中两点之间的真实距离,故常被作为度量和评价两信息间相似程度的重要指标。为进一步揭示本研究所涉三种实验贝间贝体框架特征的相似程度, 据表1中各项贝体框架特征指标的均值分别计算实验贝间的欧氏距离得表2。由表2可见, 厚壳贻贝与紫贻贝间的欧氏距离最短(P<0.05),仅为0.160; 厚壳贻贝与“杂交贻贝”间和紫贻贝与“杂交贻贝”间的欧氏距离相近(P>0.05), 分别为0.452和0.418。表明“杂交贻贝”在贝体框架特征上均显著乖离于紫贻贝和厚壳贻贝(P<0.05), 其与紫贻贝、厚壳贻贝间贝体框架特征的相似程度均远低于紫贻贝与厚壳贻贝。
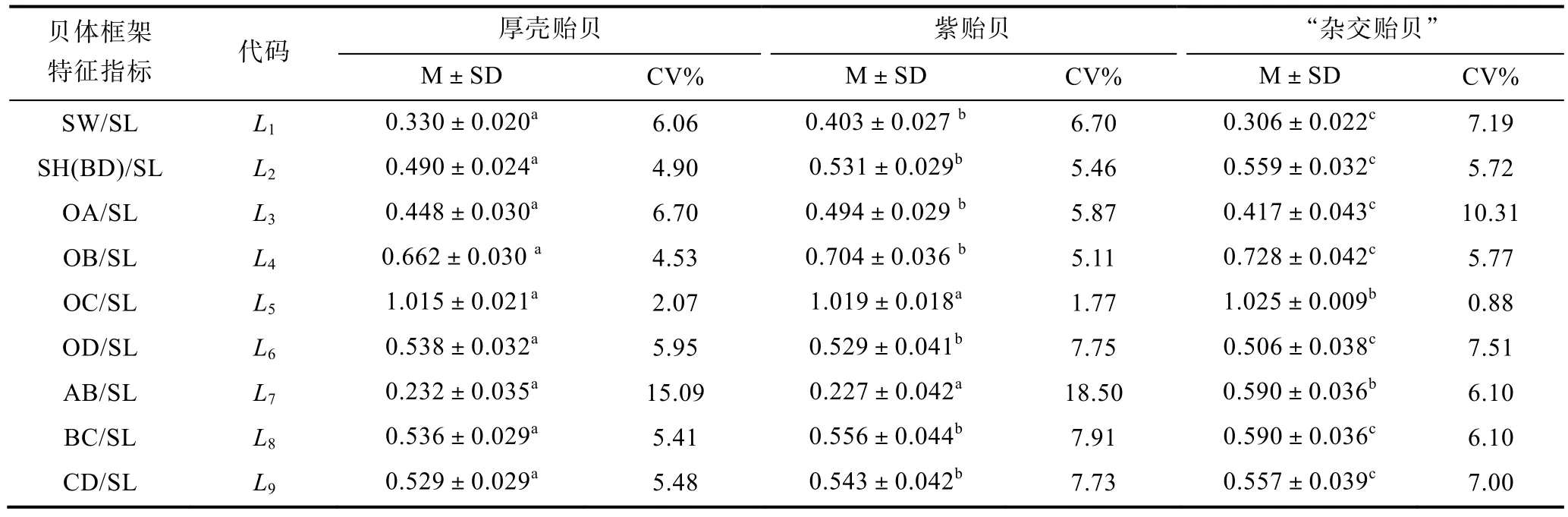
表1 三种实验贝的贝体框架特征指标Tab.1 Frame characteristic indexes of three species of Mytilus

表2 三种实验贝贝体框架特征指标间的欧氏距离Tab.2 The Euclidean distance of frame characteristics among three species of Mytilus
2.3 三种实验贝贝体框架特征的主成分分析
运用SPSS 17.0软件对贝体框架特征指标进行Bartlett球形检验和KMO适合度检验, 发现其相关系数矩阵与单位阵有显著差异(P=0.00<0.05)且适合度尚可(KMO=0.71>0.70), 表明本研究所涉贝体框架特征指标适合做因子分析, 即有进一步做主成分分析的必要性。
由表3可见, 所列3个主成分的特征值均大于1且方差累计贡献率达82.928%, 故可认定它们为能概括本研究所涉3种实验贝贝体框架特征差异的公共因子。其中, PC1的贡献率最大(41.458%)且远高于贡献率相近的PC2(21.943%)和PC3(19.527%)。将载荷绝对值P>0.5的变量确定为主要影响变量, PC1正相关主要变量的影响力排序为L8>L7>L9>L2, 负相关主要变量的影响力排序为L3>L6; PC2正相关主要变量的影响力排序为L4>L6, 负相关主要变量仅为L9; PC3正相关主要变量的影响力排序为L1>L3>L4, 无负相关主要变量。由此可知, 贡献率的大小跟各公共因子所包含的主要影响变量个数的多寡密切相关, 其中PC1不仅正相关和负相关主要影响变量个数均最多, 而且所有载荷绝对值达到0.75以上的主要影响变量也均仅存在于PC1中, 无疑其在标定本研究所涉3种实验贝贝体框架特征差异上具重要作用。

表3 三种实验贝贝体框架特征的主成分分析Tab.3 The principal component analysis of frame characteristics in three species of Mytilus
为进一步阐释PC1在标定3种实验贝贝体形态差异上的重要性, 分别绘制PC1与PC2和PC3间的得分散布图(图2)。由图2可见, 厚壳贻贝和“杂交贻贝”沿FAC1轴自左向右分别占据各自相对独立的区域,紫贻贝混于两者之间不易被区分, 而在FAC2和FAC3轴上紫贻贝、厚壳贻贝和“杂交贻贝”三者间均有较大程度的重叠。无疑, 本研究所提取到的三个主成分中惟有PC1蕴含着可较清晰标定厚壳贻贝和“杂交贻贝”间贝体框架特征差异的关键变量, 而要区分紫贻贝与厚壳贻贝和“杂交贻贝”贝体框架特征间的差异则需依赖PC1、PC2和PC3间相互结合才能最终标定。
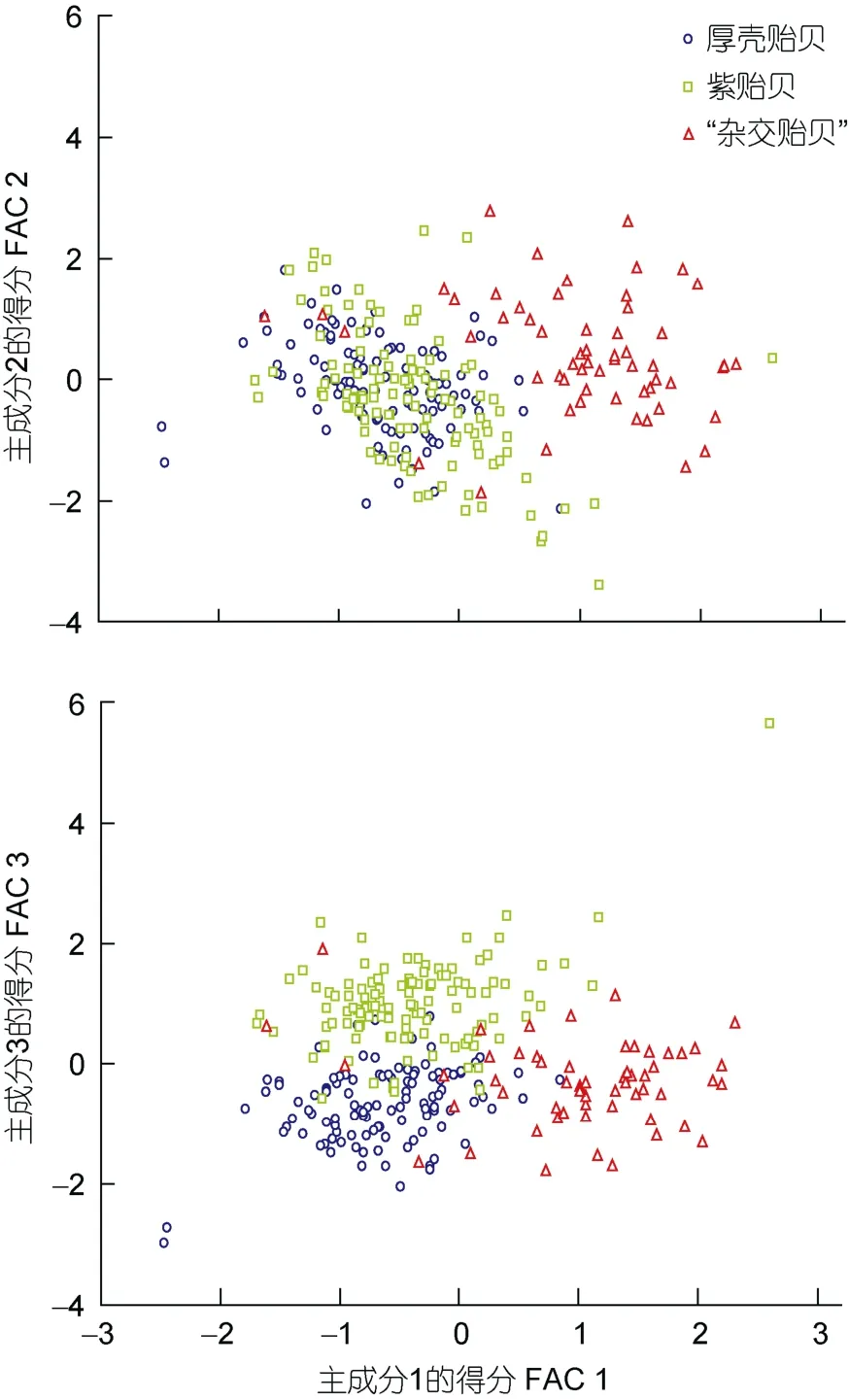
图2 三种实验贝贝体框架特征的主成分散布图Fig.2 The scatter diagram of principal component of frame characteristics in three species of Mytilus
2.4 三种实验贝贝体框架特征的判别分析
采用逐步导入剔除法, 从9个贝体框架特征变量中筛选出对判别贡献较大的L1、L3、L4、L5、L6和L7等6个变量进行判别分析,F检验表明各变量均达到极显著水平(P<0.01)。根据上述6个变量建立研究所涉各实验贝的Fisher分类函数方程组于表4, 经验证, 厚壳贻贝、紫贻贝和“杂交贻贝”的判别准确率依次为94.6%、94.6%和100%, 综合判别准确率为96.4% (表5)。另外,所绘制的典型判别函数判别得分散布图(图3), 也进一步表征了上述判别分析的可靠性。
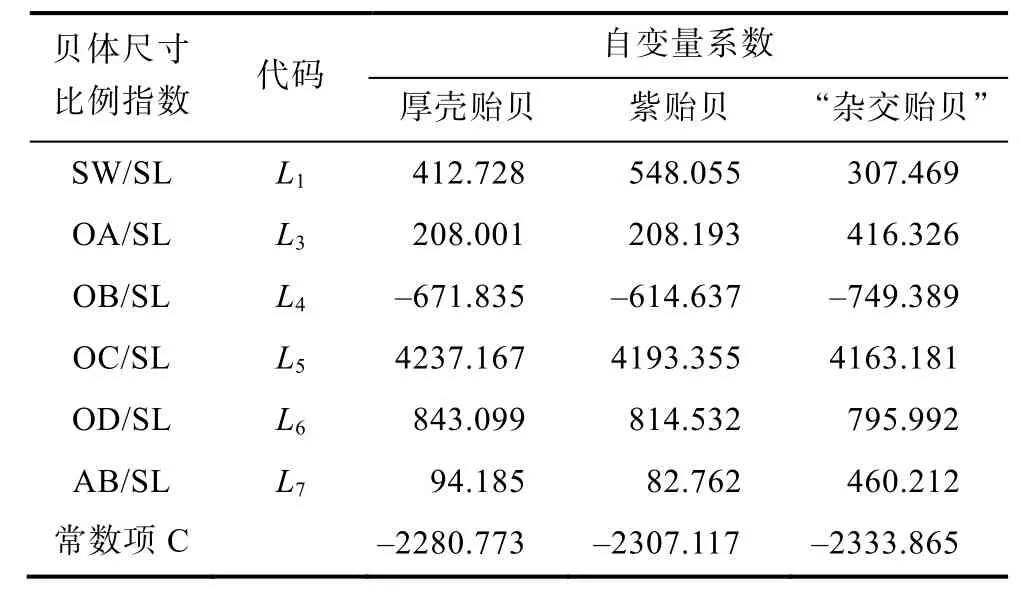
表4 三种实验贝贝体框架特征Fisher分类函数方程组的各项自变量系数及常数项Tab.4 The coefficient of variables and constant of the Fisher classification functions of frame characteristics in three species of Mytilus

表5 三种实验贝贝体框架特征的判别分类结果Tab.5 The discrimination classification of frame characteristics in three species of Mytilus
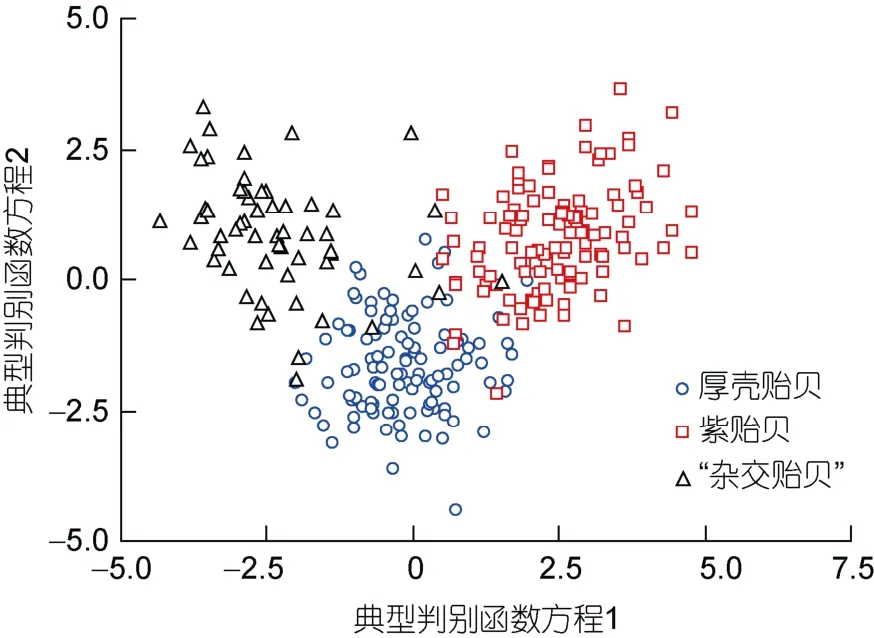
图3 三种实验贝贝体框架特征的典型判别函数判别得分散布图Fig.3 The scatter diagram of discriminant score of the Fisher classification functions of frame characteristics in three species of Mytilus
3 讨论
3.1 “杂交贻贝”贝体框架特征乖离紫贻贝和厚壳贻贝的原因
贻贝属种类系典型的滤食性附着生活型经济贝类。其外套膜属二孔型, 即具作为排泄粪便和废物出口的肛门孔与作为水流和食物进入通道的鳃足孔,两者均位于贝体后端(蔡英亚等, 1979; 王如才等,1993)。由图1可见, 实验贝贝体框架四边形ABCD内含外套腔、鳃足孔、肛门孔、直肠、肛门以及鳃区的大部分, 为其滤食功能区水平剖面; △OAD内含唇瓣、口裂、食道、胃、消化盲囊和肠, 为其消化功能区水平剖面。因此, 表4中以L2、L7、L8和L9为正相关核心变量的PC1为与滤食功能区水平剖面占比相关的贝体框架因子, 以L4、L6为正相关核心变量的PC2为与消化功能区水平剖面占比相关的贝体框架因子, 以L1、L3和L4为正相关核心变量的PC3为与消化功能区垂直剖面占比相关的贝体框架因子。上述3个公共因子所涵盖的体现实验对象组间差异的滤食功能区水平剖面、消化功能区水平与垂直剖面等一系列特征信息, 揭示了贝体滤食功能区与消化功能区空间配置状况是引起本研究所涉3种实验贝间贝体框架特征差异的主因, 从而客观地反映了贝体滤食功能区与消化功能区空间配置状况对实验贝生长速度和养成周期差异影响的形态重要性。
表1所显现的本研究所涉3种实验贝贝体框架特征差异, 也充分支持了上述滤食功能区与消化功能区配置状况为影响贝体框架特征差异主因的观点,即: (1) 已有的研究表明, 贻贝属种类对饵料种类无选择性, 只要颗粒大小适合即可(蔡英亚等, 1979; 王如才等, 1993), 故贝体滤食功能区占比值的高低将直接决定其滤食功能的强弱。由表1可见, PC1正相关核心变量L2、L8和L9均呈“杂交贻贝”>紫贻贝>厚壳贻贝(P<0.05), 仅L7呈“杂交贻贝”>紫贻贝≈厚壳贻贝, 表明三者滤食功能强度应呈“杂交贻贝”>紫贻贝>厚壳贻贝(P<0.05); (2) PC2正相关核心变量L4、L6分别呈“杂交贻贝”>紫贻贝>厚壳贻贝 (P<0.05)和厚壳贻贝>紫贻贝>“杂交贻贝”(P<0.05), PC3正相关核心变量L1、L3均呈紫贻贝>厚壳贻贝>“杂交贻贝”(P<0.05),L4呈“杂交贻贝”>紫贻贝>厚壳贻贝(P<0.05), 各实验贝上述正相关核心变量均值均呈L4>L6>L3>L1(P<0.05), 且PC2贡献率与PC3相近的结果(表1, 表3), 表明三者贝体消化功能区占比值呈紫贻贝>“杂交贻贝”≈厚壳贻贝; (3) 本研究所提取的三个主成分中PC1(41.458%)≈PC2(21.943%) + PC3(19.527%)的结果(表4), 表明滤食功能区水平剖面占比与消化功能区占比在对实验贝贝体框架特征的影响上具同等重要性。上述分析既与本研究样本采集地嵊泗列岛紫贻贝的生长速度和性成熟速度均明显较厚壳贻贝快的事实(张义浩等, 2003; 常抗美等, 2008)相吻合, 也与该海域浮游植物丰富(欧阳怡然等, 1993;张义浩, 2003), 滤食性贝类对摄食率具有很强的调节能力, 在食物保障较充足时往往会出现因来不及消化而直接将未经消化的活饵以“假粪”形式成簇散失于体外(王伟定等, 2008), 以使其摄食率在饵料浓度较高时始终保持在一定的水平的情形(Aldridgeet al,1995; Jinet al, 1996; 董波等, 2000), 以及常抗美等(2008)研究发现正交F1代(厚壳贻贝♂×紫贻贝♀)生长性能指标与紫贻贝相当而显著高于厚壳贻贝(P<0.05)的结果相符, 进一步揭示了本研究所涉“杂交贻贝”通过显著提高滤食功能区占比值来强化其杂种生长优势的本质。无疑, 表2中厚壳贻贝与紫贻贝间的欧氏距离最短(P<0.05), 而厚壳贻贝与“杂交贻贝”间和紫贻贝与“杂交贻贝”间的欧氏距离相近(P>0.05)的结果, 也充分反映了贻贝贝体滤食功能区占比较消化功能区占比对种间贝体框架特征相似性程度更具影响力的实质。
3.2 紫贻贝、厚壳贻贝、“杂交贻贝”贝体框架特征间关键判别指标组合的认定
“杂交贻贝”各项贝体框架特征指标值与紫贻贝和厚壳贻贝均具显著差异(P<0.05), 紫贻贝与厚壳贻贝间的欧式距离最短(P<0.05), 而“杂交贻贝”与紫贻贝和“杂交贻贝”与厚壳贻贝间的欧式距离相近的结果(表1, 表2), 并不妨碍“杂交贻贝”与紫贻贝间,以及紫贻贝与厚壳贻贝间在主成分FAC1、FAC2和FAC3轴上均有较大程度重叠, 而惟有FAC1轴上可将“杂交贻贝”和厚壳贻贝较清晰分开的事实(图2),既客观反映了“杂交贻贝”和厚壳贻贝在FAC1轴上隔离的情形缘于两者在贝体滤食功能区水平剖面PC1占比值间存在巨大差异的实质, 也深刻揭示了因紫贻贝与“杂交贻贝”间在贝体滤食功能区水平剖面和消化功能区占比值差异上均未达到可完全分离的程度, 致使两者在FAC1、FAC2和FAC3轴上出现相互干扰的本质(表1, 表2, 图2), 更进一步指示了PC1、PC2和PC3主要正相关变量在判别本研究三种实验贝归属问题时存在信息缺损和重叠的情形, 故须通过判别分析在补充缺损变量的同时, 剔除相关重叠变量加以校正, 以提高判别的准确性。由图1可见, 四边形OACD和OBCD共同决定着实验贝贝体水平剖面的总体框架特征, 故以壳顶O为基准的基本线段OA、OB、OC和OD势必成为构筑实验贝贝体水平剖面总体框架的形态学几何信息要素。基于基本线段OC受同一框架属性变量SL的掩盖而缺损,线段CD、SH(BD)和BC的形态学几何信息已在基本线段OA、OB、OC和OD组合中获得体现之考量, 笔者认为本研究所涉实验贝间贝体框架特征的关键判别变量组合应为L1(SW/SL)、L3(OA/SL)、L4(OB/SL)、L5(OC/SL)、L6(OD/SL)和L7(AB/SL)。
王伟定, 王志铮, 杨 阳等, 2008. 黑暗条件下缢蛏(Sinonovacula constricta)对牟氏角毛藻(Chaeroeeros moelleri)和青岛大扁藻(Platymonas subcordiformis)的滤食效应. 海洋与湖沼, 39(5): 523—528
王如才, 王昭萍, 张建中, 1993. 海水贝类养殖学. 青岛: 青岛海洋大学出版社, 119—154
李宝光, 陶秀花, 倪国平等, 2006. 扫描像素法测定植物叶片面积的研究. 江西农业学报, 18(3): 78—81
李思发, 王成辉, 程起群, 2005. Morphological variations and phylogenesis of four strains inCyprinus carpio. 水产学报,29(5): 606—611
沈玉帮, 李家乐, 牟月军, 2006. 厚壳贻贝与贻贝遗传渗透的分子生物学鉴定. 海洋渔业, 28(3): 195—200
张义浩, 赵盛龙, 2003. 嵊山列岛贻贝养殖种类生长发育调查.浙江海洋学院学报(自然科学版), 22(1): 67—73
欧阳怡然, 陈逸华, 于 波, 1993. 嵊泗列岛养殖海区浮游植物、赤潮生物的研究. 浙江水产学院学报, 12(4): 257—264
常抗美, 刘慧慧, 李家乐等, 2008. 紫贻贝和厚壳贻贝杂交及F1代杂交优势初探. 水产学报, 32(4): 552—557
董 波, 薛钦昭, 李 军, 2000. 环境因子对菲律宾蛤仔摄食生理生态的影响. 海洋与湖沼, 31(6): 636—642
蔡英亚, 张 英, 魏若飞, 1979. 贝类学概论(修订版). 上海:上海科学技术出版社, 28—214
Aldridge D W, Payne B S, Miller A C, 1995. Oxygen consumption, nitrogenous excretion and filtration rates ofDreissena polymorphaat acclimation temperatures between 20—32°C. Can J Fish Aquat Sci, 52: 1761—1767
Anderson E, Hubricht L, 1938. Hybridizatin in Tradescantia. The evidence for introgressive hybridization. Amer J Botany, 25:396—402
Caren E B, George N S, 2006. Ecological gradients and relative abundance of native (Mytilus trossulus) and invasive(Mytilus galloprovincialis) blue mussels in the California hybrid zone. Marine Biology, 148: 1249—1262
Comesana A S, Toro J E, Innes D Jet al, 1999. A molecular approach to the ecology of a mussel (Mytilus edulis,Mytilus trossulus) hybrid zone on the east coast of Newfoundland,Canada. Marine Biology, 133: 213—221
Hilbish T J, Carson E W, Plante J Ret al, 2002. Distribution ofMytilus edulis,M. galloprovincialis, and their hybrids in open-coast populations of mussels in southwestern England.Marine Biology, 140: 137—142
Inoue K S, Odo T, Noda S, 1997. A possible hybrid zone in theMytilus eduliscomplex in Japan revealed by PCR markers.Marine Biology, 128: 91—95
Jin L, Barry S P, Shiao Y W, 1996. Filtration dynamics of the zebra mussel,Dreissena polymorpha. Can J Fish Aquact Sci,53: 29—37
McDonald J H, Koehn R K, 1988. The musselsMytilus galloprovincialisandM. trossuluson the Pacific coast of North America. Marine Biology, 99(1): 111—118
Rawson P D, Secor C L, Hilbish T J, 1996. The effects of natural hybridization on the regulation of doubly uniparental mtDNA inheritance in blue mussels (Mytilusspp.). Genetics,144: 241—248
Sarver S K, Foltz D W, 1993. Genetic differentiation inMytilus galloprovincialisLarmarck throughout the world. Ophelia,47(1): 13—31
Sarver S K, Loudenslager E J, 1991. The genetics of California population of the blue mussel: further evidence for the existence of electrophoretically distinguishable species of subspecies. Biochem Syst Ecol, 119(2): 183—188
Toro J E, Thomopson R J, Innes D J, 2002, Reproductive isolation and reproductive output in two sympatric mussel species (Mytilus edulis,Mytilus trossulus) and their hybrids from Newfoundland. Marine Biology, 141: 897—909
