博斯腾湖浮游植物群落结构特征及其影响因子分析
2014-03-17马燕武谢春刚
李 红 马燕武 祁 峰 谢春刚 陈 朋
(新疆维吾尔自治区水产科学研究所, 乌鲁木齐 830000)
博斯腾湖浮游植物群落结构特征及其影响因子分析
李 红 马燕武 祁 峰 谢春刚 陈 朋
(新疆维吾尔自治区水产科学研究所, 乌鲁木齐 830000)
2011年对博斯腾湖大湖区17个采样站位的浮游植物及水体主要理化因子进行了4次系统调查。结果表明, 在17个站位共鉴定出浮游植物127种(属), 其中优势种(属)9种。浮游植物群落全年均以硅藻为主导,冬、春季节, 浮游植物组成呈硅藻-甲藻型, 优势类群主要为贫-中营养型浮游藻类, 到夏、秋季节逐渐形成硅藻-绿藻型, 以富营养型的浮游藻类为优势类群。浮游植物总平均生物量为(2.51±2.95) mg/L, 生物量季节变动显著, 峰值出现在夏季, 冬季最低。基于Canoco的多变量分析表明: 环境变量共解释了浮游植物群落总变异的54.5%, 水温是影响浮游植物分布最重要的环境因子, 其次为枝角类丰度。水中氮含量是影响浮游植物丰度的主要因子, 同时浮游植物对水体有机物含量也有较大的影响。
博斯腾湖; 浮游植物; 群落结构; 影响因子
博斯腾湖(N41°49′—42.08, E86°42′—87°25′)地处中亚干旱区, 位于新疆焉耆盆地东南部最低处,是中国干旱区最大的淡水湖, 也是唯一的吞吐湖,平均水面面积约992.2 km2, 地理位置独特[1], 其生态环境效应非常明显, 对巴音郭楞蒙古族自治州乃至整个新疆南部地区的生态都有不可替代的作用[2]。干旱区湖泊的浮游藻类生态学研究一直是水生态学的研究重点[3], 而博斯腾湖作为理想的干旱区湖泊研究对象, 其浮游植物群落动态也一直是被水环境科学、渔业资源等方面的研究所关注。20世纪 70年代至 21世纪初对博斯腾湖浮游植物有过 5次调查[4,5], 重点对浮游植物种、量及变化等情况进行了报道, 但关于浮游植物与环境因子相关关系的研究还未见报道。本文通过对博斯腾湖浮游植物和水体主要理化因子的调查研究, 阐述了博斯腾湖浮游植物群落结构特征及其演替变动情况, 并通过冗余分析(RDA)对大量的数据进行直接梯度排序, 分析了博斯腾湖浮游植物物种对环境变量的响应, 以期探索内陆干旱区湖泊浮游植物动态变动机制, 并为博斯腾湖水环境评价与保护、渔业资源合理利用和可持续发展提供背景数据。
1 材料与方法
1.1 研究湖泊概况及样点布设
分别于2011年1月、5月、8月和10月对博斯腾湖大湖区进行4次调查采样, 湖区共布设17个采样站位(图 1)。视水深情况, B05、B07-B12、B14站位(水深变幅 1.5—3.6 m)取水体中层水样; B01-B03、B13(水深变幅4.5—8.6 m)取水体上层(距水面0.5 m)和下层(距湖底0.5 m)混合水样; 其余各采样站位(水深变幅7.1—12.5 m)分别在水体表层、中层和底层分层采样。
1.2 理化因子样品的采集和测定
现场采集浮游植物样品的同时测定部分水体理化参数并采集测定其他理化参数的水样。透明度用塞氏盘测定, pH、水温、铵态氮、电导率、氯离子及溶氧用Hydrolab DS5数字化水质分析仪测定, 总有机碳用CE440型元素分析仪测定, BOD用OxiTop IS 12型分析仪测定, 其他参数的测定按照《水和废水检测分析方法(第四版)》进行测定[6]。

图1 博湖各采样站位分布Fig. 1 The distribution of each sampling station in Bosten Lake
1.3 浮游植物样品采集与观察
定性、定量标本按照章宗涉、黄祥飞《淡水浮游生物研究方法》[7]中所描述的方法进行采集、计数和现存量的计算, 浮游植物种类鉴定主要参照胡鸿钧等文献[8]。除按上述方法进行定量外, 还结合该水体的特点, 对绿裸藻、光甲藻、角甲藻、多甲藻和个体较大的硅藻等用低倍镜(10×20)数全片(0.1 mL)的办法计数, 避免了视野计数带来的误差。
1.4 数据分析
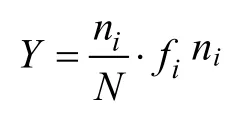
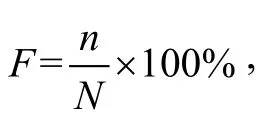
贡献率(F)计算公式:其中F为贡献率(%), n为浮游植物中某一类群的生物量, N为浮游植物群落总生物量。
浮游植物生物量均值比较采用单因素方差分析和Duncan多重比较, 应用PAWS Statistics 18.0软件进行。种类分布与环境变量之间关系分析通过canoco 4.5 软件进行, 理化因子矩阵进行lg(x+1)转化和标准化处理后再用于分析。
2 结果
2.1 博斯腾湖理化因子特征
17个站位四次采样的水体理化参数见表 1, 水温季节变化明显, 春季高于秋季; 水体溶解氧含量充沛, 监测到的最低值为5.60 mg/L, 平均值达到国家Ⅰ类水质标准[10]; 透明度变化较大, 一般呈现近岸低、中心区高的特点; 矿化度含量很高, 已属微咸水; 电导率平均为207.05 ms/m, 有升高趋势; 营养盐含量较低, 总氮年平均值为0.99 mg/L, 总磷年平均值仅有0.01 mg/L, 许多站位甚至检测不到总磷含量; 有机物含量较高, 介于Ⅲ类与Ⅳ类水质标准。
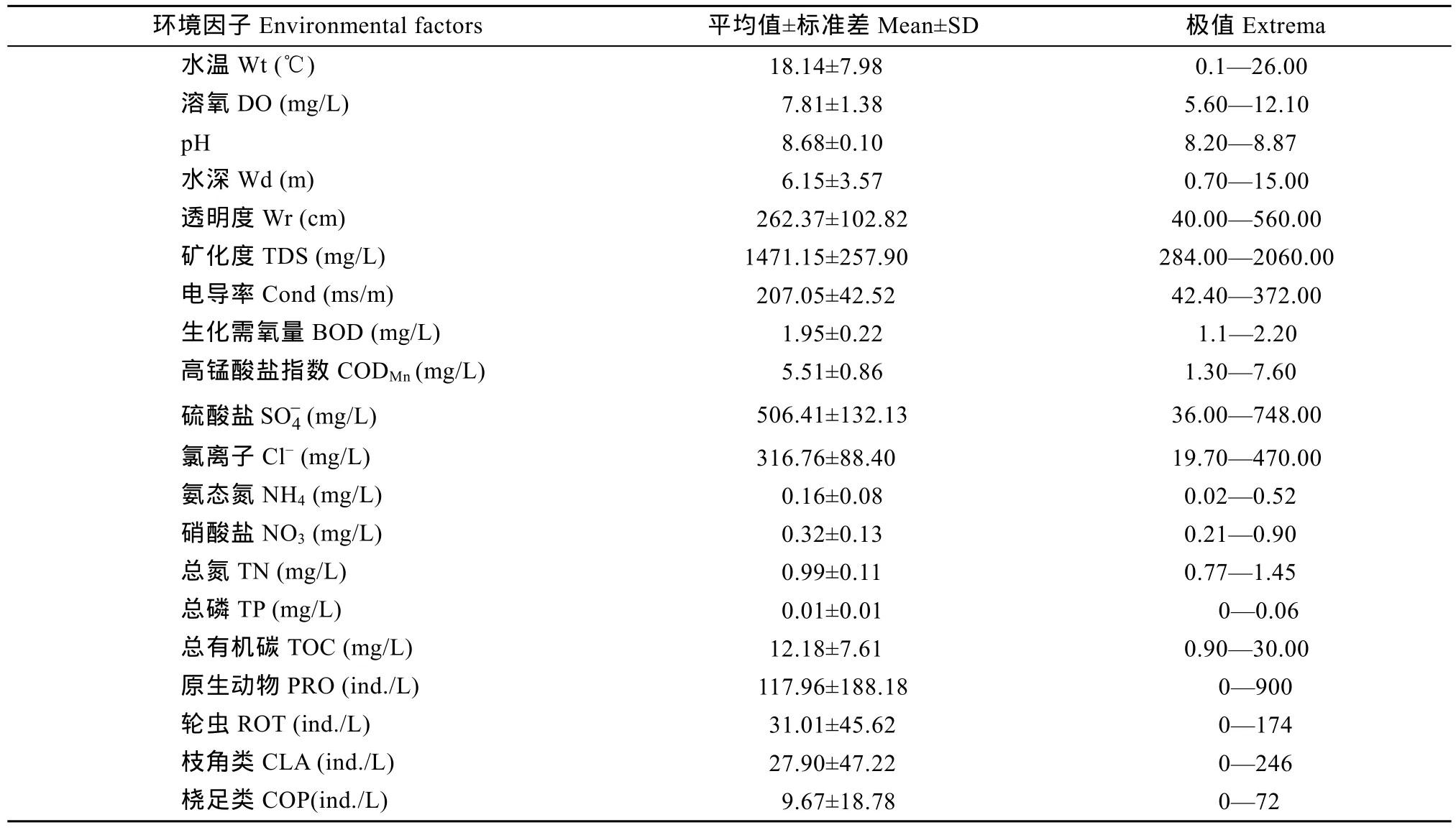
表1 博斯腾湖理化因子特征Tab. 1 Characteristics of environmental factors in Bosten Lake

表2 博斯腾湖浮游植物种类组成Tab. 2 Phytoplankton species composition in Bosten Lake

硅藻门Bacillariophyta空星藻Coelastrum sp.直链藻Melosira sp.十字藻Crucigenia sp.角刺藻Chaetoceros sp. 微芒藻Micractinium sp.小环藻Cyclotella sp. 疏刺多芒藻Golenkinia paucispina具星小环藻C. stelligera 被刺藻Franceia sp.纤细等片藻Diatoma tenue 单角盘星藻Pediastrum simplex普通等片藻D. vulgare 二角盘星藻P. duplex平板藻Tabellaria sp.四角盘星藻P. tetras脆杆藻Fragilaria sp. 短棘盘星藻P. borganum钝脆杆藻F. capucina 四尾栅藻Scenedesmus quadricauda尖针杆藻Synedra acus 双对栅藻S. bijuga肘状针杆藻S. tenera 弯曲栅藻S. arcuatus尺骨针杆藻 S. ulan 四刺顶棘藻Chodatella quadriseta头状针杆藻S. capitata 十字顶棘藻C. wratislaviensis美丽星杆藻Asterionella formosa 纤毛顶棘藻C. cillita窗纹藻Epithemia sp. 卵囊藻Oocystis sp.弧形蛾眉藻Ceratoneis arcus 单生卵囊藻O. solitaria布纹藻Gyrosigma sp. 湖生卵囊藻O. lacustris尖布纹藻G. acuminatum 转板藻Mougeotia sp.辐节藻Stauroneis sp. 新月藻Closterium sp.双头辐节藻S. anceps 纤细新月藻C. giacile曲壳藻Achnanthes sp. 鞘藻Oedogoniu sp.卵圆双眉藻Amphora ovalis 鼓藻Cosmarium sp.舟形藻Navicula sp.
续表2
2.2 浮游植物群落结构特征
种类组成及优势种类 17站位共鉴出浮游植物127种(属), 其中硅藻门最多45种(属), 占总种(属)数的35.4%。绿藻门次之41种(属), 占总种(属)数的32.3%。其余依次为蓝藻门25种(属), 甲藻门5种(属), 金藻门4种(属), 裸藻门4种(属), 隐藻门3种, 分别占总种(属)数的19.7%、3.1%、3.1%和2.4%。4次采样调查形成过优势类群(Y>0.02)的种类均为淡水习见种类(表3), 共9种(属), 分别为尖针杆藻、小环藻、舟形藻、鼓藻、小球藻、四角藻、席藻、多甲藻、锥囊藻, 其中尖针杆藻全年分布, 在群落中占绝对优势(8月和10月Y>0.7), 飞燕角甲藻和圆筒锥囊藻亦是全年分布的种类, 每次调查均可观测到,调查中在 B7采样点尚发现一种在淡水中少有出现的角毛藻。
浮游植物群落组成及生物量的变动 2011年 1月份, 博斯腾湖各站位浮游植物平均生物量为(0.231±0.18) mg/L, 其中硅藻占总生物量的 51.8%,其次为甲藻占总生物量的 33.6%, 其余各类群对总生物量的贡献率均不到5%。5月各站位浮游植物平均生物量为(0.681±0.41) mg/L, 硅藻对生物量的贡献率升高至63.6%, 甲藻有所降低, 约为23.4%, 绿藻的贡献率增至8.2%。8月各站位浮游植物平均生物量为(6.695±5.44) mg/L, 硅藻对生物量的贡献率达81.5%, 绿藻所占比例为11.4%, 甲藻仅占5.6%。与8月份比较, 10月份浮游植物各类群对生物量的贡献率变化不大, 但平均生物量降至(2.414± 1.82) mg/L。经方差分析(Duncan多重比较), 8月和10月浮游植物生物量与其他各月生物量差异均显著或极显著(P<0.05, P<0.01),1月和5月之间差异不显著。表明: 博斯腾湖浮游植物1到5月份生长较慢, 8月(夏季)生长旺盛, 之后逐渐减少。

表3 博斯腾湖浮游植物优势种(属)及优势度Tab. 3 The dominant phytoplankton species and their dominance in Bosten Lake

图2 2011年浮游植物不同类群对浮游植物生物量的贡献率(%)及生物量(mg/L)变动Fig. 2 Relative biomass (%) of phytoplankton classes (upper panel) and mean biomass (mg/L) (lower panel) in Bosten Lake in 2011
2.3 种类分布与环境因子RDA分析
DCA分析显示最大梯度长度为 2.469, 意味着浮游植物与环境因子之间的关系呈线性, 依此将20个环境变量(表 1)与浮游植物优势种类(表 3)进行RDA 分析。经向前选择和蒙特卡洛(Monte Carlo) P<0.05检验。共保留6个环境变量, 分别为水温(Wt, P=0.002)、桡足类(COP, P=0.002)、总氮(TN, P=0.06)、硝酸盐(NO3, P=0.012)、枝角类(CLA, P=0.004)和CODMn(P=0.02), 前4个轴共解释浮游植物群落结构总变异的54.5%。在RDA排序图中Wt与物种轴1呈最大相关性, 相关系数为–0.9171, 其次为 CLA,相关系数为–0.5902。
图3中两物种射线之间夹角的余弦值在数值上等于两者的相关系数, 物种射线和变量射线之间夹角的余弦值代表了二者之间的相关性。B1、C4和D1这些冬季优势类群分布在第四和第三象限, 与其他优势种及变量呈明显的负相关; 夏、秋季节浮游植物优势种类差异不大, 与环境变量之间呈很好的正相关; 春季优势种类较多, 有的是冬季的优势种类(C4和D1), 到夏季失去优势地位, 有的继续繁衍为夏、秋季优势种类(B3、B5和C5), 且大多集中在第二轴附近, 与变量之间相关性较弱。
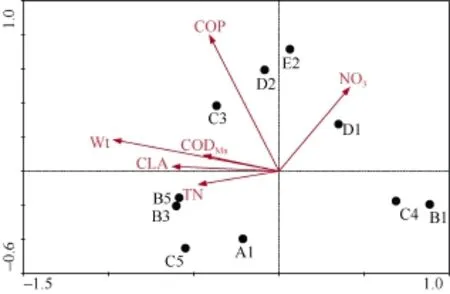
图3 浮游植物种类分布与环境因子之间的关系Fig. 3 Species associations of phytoplankton with environmental factors
3 讨论
3.1 博斯腾湖浮游植物群落结构特征及变动
博斯腾湖为典型的干旱区湖泊, 与其他干旱、高寒地区湖泊类似, 极易形成以硅藻为优势类群的群落特征[11—14], 湖区浮游植物群落全年均以硅藻为主导。冬春季节浮游植物组成呈硅藻-甲藻型, 优势种类主要为贫-中营养型湖泊常见的优势浮游藻类, 如小环藻、多甲藻、锥囊藻等; 到夏秋逐渐形成硅藻-绿藻型, 浮游植物以富营养的针杆藻、绿藻、蓝藻等为优势类群, 有些站位还检测到裸藻的发生。
1989—2001年的调查表明博斯腾湖浮游植物组成呈典型的硅藻-蓝藻-绿藻型。本次调查与历史数据相比较, 博斯腾湖浮游植物以硅藻为主导的优势地位没有改变, 硅藻优势种类变化也不大, 如同为针杆藻、小环藻、舟形藻等淡水普生种类, 但优势种类的主导地位更加突出, 如尖针杆藻在8月和10月其优势度达到了 0.7以上, 硅藻对生物量的贡献率由2001年的51.8%升至67.7%。其他类群的组成也发生了较为明显的变化, 表现在:
(1)蓝藻对生物量的贡献率有比较大的减少, 由17.2%降至1.4%, 其优势种类也发生了变化。1989—2001年蓝藻的优势种类, 如微囊藻、鱼腥藻、颤藻及平裂藻, 虽然在本次调查中也可发现, 但均未形成过优势种群, 可形成优势种群的是席藻(出现在10月)。此外, 鉴出的蓝藻种(属)数也有所降低, 由34种(属)减少到25种(属)。(2)大型浮游藻类减少。如1989—2001年飞燕角甲藻是全湖的优势种类, 几乎每个站位都占据优势地位, 本次调查每次采样均可发现其分布, 但未形成优势种群, 只有多甲藻在藻类竞争中曾占一席之地。另一方面, 小型浮游藻类明显增多, 如小环藻、小型绿球藻类、四角藻、蹄形藻、顶极藻等, 在一定时期内甚至可形成优势种群。(3)另外一个变化特点是金藻出现率增加, 每次采样均可观测到, 主要在 5月繁盛并形成优势类群。(4)咸水种的出现。此次调查中在 B7采样点中发现了角毛藻, 这是一种通常只在咸水中常见的种类, 淡水中仅偶有出现, 但联系本次调查的浮游动物, 其中10余个样本中都有一定数量的咸水习见种——褶皱臂尾轮虫(Brachionusplicatilis)的出现, 说明角毛藻的出现并非偶然。本次调查发现湖区水体矿化度均值达1741.15 mg/L, 已属微咸水。可见, 随着博斯腾湖水咸化的加剧, 对其水生生物资源产生了一定深远影响, 也在一定程度上反映了干旱地区水质和水生生物群落状况。
本次调查浮游植物生物量季节变动较大, 8月峰值约是1月平均生物量的289倍, 冬春季生长缓慢,夏季生长迅速, 秋季以后逐渐衰退, 这与其他一些温带干旱区湖泊浮游植物变动情况类似[12,15—17]。在进入夏季以后, 由于水温回升, 天然水源补给增多以及养殖生产、生活、旅游和工业污水等排放的加剧, 湖区营养盐、有机物含量增加, 如本次监测结果表明: 各站位8月和1月均值相比, CODMn增长了141.2%、总氮增长了117.8%、氨氮增长了 257.1%,因此夏季浮游植物进入快速生长期。博斯腾湖封冰期有 4个月, 夏秋季节积累的营养盐消耗后, 加之水温条件的限制, 所以冬、春季浮游植物生长非常缓慢。与历史同期数据相比, 浮游植物平均生物量约是1989—2001年调查数据的1.2倍, 变幅较小。与地理位置近似的乌伦古湖[13,15]、艾比湖[11]浮游植物平均生物量近似; 低于湿润区和云贵高原富营养化湖泊, 如武汉太湖[18]、上海淀山湖[19]、巢湖[20]以及云南长桥海、大屯海、滇池等湖泊[21]; 高于新疆赛里木湖[22]以及云南的草海、阳宗海、扶仙湖等湖泊[21]。
3.2 环境因子与浮游植物群落关系
湖泊理化因子的显著季节变动对浮游植物群落演替起着重要的调控作用, 以致对浮游植物群落性质和量的改变[23,24]。RDA分析初步反映了博斯腾湖浮游植物群落与主要环境因子之间的相关关系。
在所测的环境因子中, 水温是影响博斯腾湖浮游植物群落演替最重要的环境因子, 不同浮游植物类群有着不同的最适温度。博斯腾湖硅藻为全年的优势种群, 其现存量呈现明显的季节变动。监测期间博斯腾湖平均水温为16.14 , ℃ 检测到最高水温不过 26.5 , ℃ 适合多喜低温的硅藻生长繁殖。蓝藻主要出现在秋季, 金藻在春季繁盛, 绿藻在各季节均可形成优势类群, 甲藻主要在低水温期占据一定优势。其他学者的研究也同样表明: 金藻、硅藻适宜在较低的水温下生长, 故多出现早春、晚秋和冬季;绿藻在中等或较高的水温下占优势, 而蓝藻更适宜生长在温暖的水体中, 故多出现于夏秋季[25—26], 甲藻经常在我国北方越冬水体中占据优势地位[27,28]。
通过限制浮游植物生长率和影响其种类组成, 营养盐的变动能够影响浮游植物群落的繁盛[29]。TN︰TP经常被用于衡量可能的限制性营养盐, 通过对一些湖泊的研究, Guildford等认为如果TN︰TP > 50, 表现磷限制; TN︰TP < 20 则是氮限制; 介于20—50之间氮、磷均不会成为限制因子[30]。博斯腾湖TN︰TP约为99, 表现为磷限制。实测数据也表明总磷含量最高不过 0.06 mg/L, 很多站位经常检测不到。RDA分析表明浮游植物种类与硝酸态氮和总氮有很好的相关性, 说明氮含量的变化很大程度上影响浮游植物丰度, 而磷含量的基础值很低, 变化也不大, 所以与浮游植物丰度的相关性很低。
RDA分析也显示了17个站点浮游植物丰度与CODMn有很好的相关性, 表明浮游植物对博斯腾湖水体有机物含量有较大的影响。其他学者也发现在COD含量较高时, 浮游植物的密度相对较高, 与浮游植物种群的变化表现出明显的相关性[31,32]。
枝角类丰度是除水温之外最显著的环境变量,可见其群落变动可较显著地影响博斯腾湖浮游植物群落动态。在 RDA分析图中仅从枝角类与浮游植物群落的关系看, 枝角类与夏、秋季的浮游植物类群呈正相关, 与冬季的优势种类呈负相关。这可能由于二者的动态变化均主要受水温的调控(水温与枝角类丰度的相关系数r=0.5433, 与浮游植物群落 r= –0.9171)。而轮虫由于在物种丰度和摄食压力上较弱, 在本研究中其对博斯腾湖浮游植物的影响较小。RDA分析尚表明桡足类丰度与浮游植物丰度有一定的相关性。博斯腾湖桡足类的优势种类为台湾温剑水蚤(Thermocyclops taihokuensis)和特异中剑水蚤(Mesocyclops dissimilis), 关于其生物学的研究还很少, 分析可能通过营养级联效应影响浮游植物群落动态。
由于博斯腾湖生态环境的复杂性, 除主要的环境因子外, 水文条件及浮游植物食性鱼类等的下行效应也在很大程度上影响着浮游植物的群落结构,这些都有待进行深入研究。
致谢:
大连海洋大学李永函老师和中国科学院水生生物研究所冯伟松老师在浮游植物鉴定和文章撰写过程中进行重要的指导, 谨致谢忱。
[1] Wang S M, Dou H S. Lake of China [M].Beijing: Science Press. 1998, 345 [王苏民, 窦鸿身.中国湖泊志.北京: 科学出版社. 1998, 345]
[2] Ran X J, Shen L, Li X H.Analysis of ecological environment effects in Bosten Lake [J].Journal of Water Resources and Water Engineering, 2010, 21(1): 105—108 [冉新军, 沈利, 李新虎. 博斯腾湖生态环境效应分析.水资源与水工程学报, 2010, 21(1): 105—108]
[3] Rothhaupt K O.Plankton population dynamics: food web interactions and abiotic constraints [J].Freshwater Biology, 2000, 45: 105—109
[4] Comprehensive Expedition of Resource Exploitation in Xinjiang, Chinese Academy of Sciences.Hydrobiology and Fishery in Xinjiang [M].Beijing: Science Press. 1989, 18—42 [中国科学院新疆资源开发综合考察队.新疆水生生物与渔业.北京: 科学出版社. 1989, 18—42]
[5] Guo Y, Zhang R M, Cai L G, et al.Fish Resources and Fishery [M].Urumqi: Xinjiang Science and Technology Press. 2005, 2—72 [郭焱, 张人铭, 蔡林刚, 等.博斯腾湖鱼类资源及渔业.乌鲁木齐: 新疆科学技术出版社.2005, 2—72]
[6] Editorial Committee of Water and Waste Water Testing Methods, State Environmental Protection Administration. Testing Methods of Water and Waste Water (the 4th Edition) [M]. Beijing: China Environmental Science Press. 2002, 38—47 [国家环境保护总局水和废水检测分析方法编委会.水和废水检测分析方法(第四版).北京: 中国环境科学出版社. 2002, 38—47]
[7] Zhang Z S, Huang X F.Research Methods of Freshwater Plankton [M].Beijing: Science Press. 1991, 88—186 [章宗涉, 黄祥飞.淡水浮游生物研究方法.北京: 科学出版社. 1991, 88—186]
[8] Hu H J, Wei Y X.The Freshwater Algae of China—Systematic, Taxonomy and Ecology [M].Beijing: Science Press. 2006, 79—285 [胡鸿钧, 魏印心.中国淡水藻类—系统、分类及生态.北京: 科学出版社.2006, 79—285]
[9] Asknes L, Wassman P.Modeling the significance of zooplankton grazing for export production [J].Limnology Oceanography, 1933, 38: 978—985
[10] GB3838-2002, Environmental Quality Standards for Surface Water [S] [GB3838-2002, 地表水环境质量标准]
[11] Dong Z W.Dynamic of plant community in rehabilitating wetland—A case study in Jinghe River Estuary [D].Urumqi: Xinjiang Normal University. 2011 [董正武.退化湿地恢复过程中浮游植物群落的变化——以艾比湖精河入湖口为例.乌鲁木齐: 新疆师范大学. 2011]
[12] Çetin A K, Şen B.Seasonal distribution of phytoplankton in Orduzu Dam Lake (Malatya, Turkey) [J].Turkish Journal of Botany, 2004, 28: 279—285
[13] Li Z, Jiang Z F, Ma B, et al. Cluster analysis and multidimensional scaling analysis in spring and autumn of phytoplankton community structure of Ulunger Lake, Xinjiang [J].Journal of Fishery Sciences of China, 2008, 15(6): 984—961 [李 喆 , 姜作发, 马波, 等.新疆乌伦古湖春、秋季浮游植物群落结构的聚类和多维分析.中国水产科学, 2008, 15(6): 984—991]
[14] Serra T, Colomen J, Baserba C, et a1.Quantified distribution of diatoms during the stratified period of Boadella reservoir [J]. Hydrobiologia, 2002, 589: 235—244
[15] Liu Y, Shen J Z, LI H, et al.Phytoplankton community structure in Lake Ulungur in Xinjiang Uygur Autonomous Region [J].Journal of Lake Sciences, 2009, 21(6): 855—864 [刘宇, 沈建忠, 李鸿, 等.新疆乌伦古湖浮游植物群落结构.湖泊科学, 2009, 21(6): 855—864]
[16] Dong Z W.?Dynamic of plant community in rehabilitating wetland—A case study in Jinghe River Estuary [D].Xinjiang Normal University, Urumqi. 2011 [董正武.退化湿地恢复过程中浮游植物群落的变化——以艾比湖精河入湖口为例.新疆师范大学, 乌鲁木齐. 2011]
[17] Liu W Y, Gao R H, Zhang Q L, et al.The diversity of ?phytoplankton in salina-alkaline wetland on the Erdos ?Plateau [J]. Journal of Arid Land Resources and Environment, 2009, 5(6): 143—148 [刘文盈, 高润宏, 张秋良,等.鄂尔多斯高原盐沼湿地浮游植物的多样性与评价. 干旱区资源与环境, 2009, 5(6): 143—148]
[18] Qian K M, Chen Y W, Song X L. Long-term development of phytoplankton dominant species related to eutrophication in Lake Taihu [J].Ecological Science, 2008, 27(2): 65—70 [钱奎梅, 陈宇炜, 宋晓兰. 太湖浮游植物优势种长期演化与富营养化进程的关系. 生态科学, 2008, 27(2): 65—70]
[19] Wang L Q, Shi R, Ji G H, et al.Phytoplankton community structure and its succession in Dianshan Lake [J]. Biodiversity Sciences, 2011, 19(1): 48—56 [王丽卿, 施荣,季高华, 等.淀山湖浮游植物群落特征及其演替规律.生物多样性, 2011, 19(1): 48—56]
[20] Meng R X, Liu Z Q. Evaluation of water pollution and eutrophication in the Chaohu Lake by means of phytoplankton [J]. Acta Hydrobiologica Sinica, 1988, 12(1): 13–26 [蒙仁宪, 刘贞秋.以浮游植物评价巢湖水质污染及富营养化.水生生物学学报, 1988, 12(1): 13—26]
[21] Zhang M, Yu Y, Qian S Q, et al.Phytoplankton community structure and biodiversity in summer Yunnan-Guizhou Plateau lakes [J].Journal of Lake Sciences, 2010, 22(6): 829—836 [张民, 于洋, 钱善勤, 等.云贵高原湖泊夏季浮游植物组成及多样性.湖泊科学, 2010, 22(6): 829—836]
[22] Yang W R, Guo Y, Cai L G, et al.Research of fishery situation and feed creature of Sailimu lake [J].Chinese Journal of Fisheries, 2000, 13(1): 1—10 [杨文荣, 郭焱, 蔡林刚, 等.赛里木湖饵料生物及渔业现状的研究.水产学杂志, 2000, 13(1): 1—10]
[23] Kalin M, Cao Y, Smith M, et al.Development of the phytoplankton community in a pit-lake in relation to water quality changes [J].Water Research, 2011, 35(13): 3215—3225
[24] Fathi A A, Abdelzaher H, Flower R J, et al.Phytoplankton communities of North African wetland lakes: the Cassarina Project [J].Aquatic Ecology, 2001, 35: 303—318
[25] Dokulil M T, Teubner K.Cyanobacterial dominance in lakes [J].Hydrobiologia, 2000, 438: 1—12
[26] Shen Y F, Zhang Z S, Gong X J.Modern Biomonitoring Techniques Using Freshwater Microbiota [M].Beijing: China Architecture and Building Press. 1990, 67—228 [沈韫芬, 章宗涉, 龚循矩.微型生物监测新技术.北京: 中国建筑工业出版社. 1990, 67—228]
[27] Li Y H, Qiu R B, Guo J E, et al.Studies on the under-ice plankton of the overwintering fishponds [J]. Acta Hydrobiologica Sinica, 1985, 9(2): 117—128 [李永函, 邱日彬, 郭继娥, 等.越冬池冰下浮游生物的研究.水生生物集刊, 1985, 9(2): 117—128]
[28] Qi F, Liu X, Zhao Y H, et al.Study on plankton in overwintering seawater ponds [J].Fisheries Science, 2013, 32(1): 7—14 [祁峰, 刘胥, 赵艳红, 等.海水越冬池冰下浮游生物群落结构特征的研究.水产科学, 2013, 32(1): 7—14]
[29] Haande S, Rohrlack T, Semyalo R P, et al.Phytoplankton dynamics and cyanobacterial dominance in Murchison Bay of Lake Victoria (Uganda) in relation to environmental conditions [J].Limnologia, 2011, 41(1): 20—29
[30] Guildford S J, Hecky R E.Total nitrogen, total phosphorous, and nutrient limitation in lakes and oceans: is there a common relationship [J]? Limnology Oceanography, 2000, 45: 1213—1223
[31] Lü J, Wu H, Chen M.Effects of nitrogen and phosphorus on phytoplankton composition and biomass in 15 subtropical, urban shallow lakes in Wuhan, China [J].Limnologia, 2011, 41(1): 48—56
[32] Chen J F, Qi Y Z, Xu N, et al.Annual dynamics of phytoplankton community and abundance at aotou cove, Daya Bay: 1997—1998 [J].Acta Hydrobiologica Sinica, 2006, 30(3): 311—317 [陈菊芳, 齐雨藻, 徐宁, 等.大亚湾澳头水域浮游植物群落结构及周年数量动态.水生生物学报, 2006, 30(3): 311—317]
PHYTOPLANKTON IN BOSTEN LAKE: COMMUNITY CHARACTERISTICS AND DRIVING FACTORS
LI Hong, MA Yan-Wu, QI Feng, XIE Chun-Gang and CHEN Peng
(Xinjiang Fishery Research Institute, Urumqi 830000, China)
In order to study the community characteristics and their driving factors in the great region of Bosten Lake, four investigations on phytoplankton and physiochemical factors were carried out in 2011. We identified 127 species, 9 species out of which once formed dominant populations. Diatom was predominant in the phytoplankton community all over the year. In winter and spring the phytoplankton community mainly consisted of diatom and dinoflagellate, floating algae being predominant in dys-mesotrophic lakes. However in summer and fall the major composition of phytoplankton were diatom and green alga, eutrophic type being the dominant in the population. The total mean biomass of phytoplankton was (2.51±2.95) mg/L. The peak of the mean biomass was reached in summer and the lowest in winter. Canoco multivariate analysis indicated that 54.5% of the total variance in the phytoplankton community could be explained with environment variables. The water temperature and the abundance of Cladocera were found to be the most and the second most important factors affecting the distribution of phytoplankton. The phytoplankton abundances were largely affected by the variations of nitrogen contents. Moreover, phytoplankton had profound effects on the concentrations of organism in Bosten Lake.
Bosten Lake; Phytoplankton; Community structure; Driving factors
Q145+.2
A
1000-3207(2014)05-0921-08
10.7541/2014.137
2013-07-09;
2014-02-23
水体污染控制与治理科技重大专项项目(2009ZX07106-004)资助
李红(1963—), 女, 山东人; 高级工程师; 主要从事浮游生物研究工作。E-mail: lihong6092@163.com
马燕武, 男, 研究员; E-mail: myw0012@126.com
