水解鱼蛋白对大菱鲆幼鱼消化率的影响
2014-03-17卫育良梁萌青郑珂珂王新星
卫育良梁萌青郑珂珂王新星
(1. 中国海洋大学教育部海水养殖重点实验室, 青岛266003; 2. 中国水产科学研究院黄海水产研究所, 青岛 266071)
水解鱼蛋白对大菱鲆幼鱼消化率的影响
卫育良1,2梁萌青2郑珂珂2王新星2
(1. 中国海洋大学教育部海水养殖重点实验室, 青岛266003; 2. 中国水产科学研究院黄海水产研究所, 青岛 266071)
设计5组等氮等脂等能的饲料, 在室内流水系统进行68d的养殖实验, 探讨高植物蛋白饲料中添加不同分子量水解鱼蛋白对大菱鲆Scophthalmus maximus L.[(16.05±0.03) g]幼鱼消化能力的影响。分别在高植物蛋白饲料中添加5.4%超滤水解鱼蛋白(UF)、5.5%未经超滤水解鱼蛋白(FPH)、5.5%超滤截留水解鱼蛋白(RF),其均占饲料蛋白的 10%, 以及不添加水解蛋白(PP), 以上各组鱼粉含量均为 18%, 对照组(FM)鱼粉含量为67.5%。研究结果表明, FM组的大菱鲆特定生长率与UF、FPH及PP组无显著差异(P>0.05); 饲料效率、蛋白质效率和蛋白质沉积率在FM组与UF组无显著差异(P>0.05), 但显著高于FPH、RF及PP组(P<0.05); 饲料干物质和蛋白质消化率在UF、FPH及RF组显著高于PP组(P<0.05), 但显著低于FM组(P<0.05), 在UF组显著高于FPH和RF组(P<0.05); 不同处理对饲料氨基酸和牛磺酸的消化率均产生显著影响(P<0.05), 趋势为FM组最高, 其次为UF组, PP组最低; 半胱氨酸和牛磺酸的消化率在添加水解鱼蛋白的3组(UF、FPH和RF组)和不添加水解鱼蛋白的2组(FM和PP组)呈相反的趋势。上述结果表明, 在高植物蛋白饲料中添加低分子量水解鱼蛋白(UF), 大菱鲆幼鱼的生长和饲料利用有升高的趋势, 但UF、FPH以及RF都显著提高了大菱鲆对饲料干物质、蛋白质和氨基酸的消化率, 且UF效果优于FPH和RF。此外, 添加不同分子量水解鱼蛋白都降低了牛磺酸的消化率。
大菱鲆; 水解鱼蛋白; 生长; 饲料利用; 消化率
世界水产养殖产量在最近十年间持续增加使得对饲料需求量越来越大[1], 而水产饲料中蛋白原料的选取不仅直接影响饲料的质量和价格, 还影响整个水产业的综合效益[2]。鱼粉是肉食性鱼类饲料主要的蛋白源, 也是构成饲料成本的主要成分, 由于鱼粉需求量的增加而供应量保持相对稳定, 因此鱼粉在近几年一直维持较高的价格[2]。这些都迫使人们不得不一方面尽量寻找廉价的植物蛋白源来替代鱼粉, 另一方面充分利用现有海洋性蛋白。在植物蛋白的利用上, 当植物蛋白替代大部分或全部鱼粉后, 某些肉食性鱼类即使在补充外源营养物质的前提下仍然会出现生长显著降低[3,4]。Hardy等[5]提出解决鱼粉短缺问题目前条件下一种可行的方法是充分利用现有的渔业资源, 特别是低价值的水产品加工副产品或者废弃物。水解鱼蛋白主要以低值鱼或水产副产品为原料, 通过用酶或酸等方法水解鱼体蛋白而获得富含生物活性肽的蛋白产品[6,7], 其作为优质蛋白源受到广泛的关注。将水解鱼蛋白以适当的比例添加至高植物蛋白的饲料中, 能够显著提高肉食性鱼类的生长、饲料利用、血浆 IGF-I水平和肝脏的IGF-I的mRNA表达量[8,9]。进一步研究发现, 水解鱼蛋白中低分子量组分是维持肉食性鱼类在饲喂高植物蛋白饲料条件下保持正常生长必不可少的成分[9,10]。
当前, 在对水解鱼蛋白替代鱼粉的研究中, 更多的关注集中在鱼蛋白水解物中含有的非蛋白氮如牛磺酸、羟脯氨酸、鹅肌肽等的作用上[10,11], 而对水解鱼蛋白这种提前在体外水解的蛋白源添加到饲料中对饲料消化的影响却很少报道, 且主要集中在饲料营养物质对蛋白质、脂肪、能量和总磷的表观消化率[8,10,12,13]。植物蛋白影响肉食性鱼类生长的一个重要的原因是降低了鱼类对饲料的消化率[14],因而, 研究能否通过添加水解鱼蛋白来改善植物蛋白饲料的消化成为研究水解鱼蛋白在高植物蛋白饲料中利用效果的一个重要方面。
本文研究在高植物蛋白饲料中添加不同分子量的水解鱼蛋白对大菱鲆(Scophthalmus maximus L.)幼鱼生长、饲料利用和消化率的影响, 为水解鱼蛋白对肉食性鱼类的生长作用提供进一步的数据。
1 材料与方法
1.1 水解鱼蛋白的制备及分子量分布测定
实验用去头和内脏的太平洋狭鳕鱼(Theragra chalcogramma), 购自亿路发水产有限公司(中国,青岛)。水解鱼蛋白的制备参考 Zheng等[12]的方法,选用碱性蛋白酶和风味蛋白酶, 鱼肉和水比例 1︰1,调节pH到8.5—9.0, 55℃水解3h, 过滤, 然后在室温下离心(1700×g, 10min)。将所得酶解液体用Pellicon2 超滤膜堆装置(1000 D)超滤, 通过超滤膜得到水解鱼蛋白记做 UF(分子量低), 经超滤膜截留的水解鱼蛋白记做 RF(分子量高), 未经超滤的水解蛋白记做 FPH, 分别将其冷冻干燥保存备用。制备的水解鱼蛋白分子量分布采用排阻色谱法测定, 色谱柱用TSK G2000 SWXL 300 mm×7.8 mm, 检测波长为220 nm, 选用碳酸酐酶(MW 29000)、细胞色素C(MW12400)、抑肽酶(MW6511.44)、人血管紧张素II(MW1046.18)、谷胱甘肽(MW 307.32)、乙氨酸—乙氨酸—乙氨酸(MW189)和甘氨酸(MW75.07)为标品作标准曲线。3种水解鱼蛋白分子量分布见表 1,化学组成和氨基酸组成见表2。
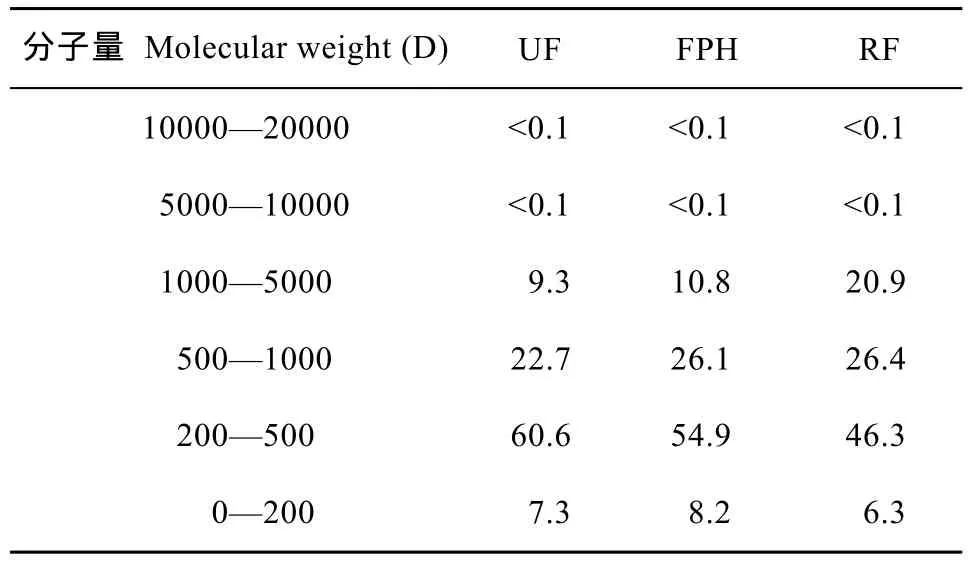
表1 UF、FPH和RF三种水解鱼蛋白的分子量分布Tab.1 Molecular weight distribution of UF, FPH and RF
1.2 实验饲料
实验选用鱼粉、豆粕、大豆浓缩蛋白和玉米蛋白为主要蛋白源, 鱼油为主要脂肪源, 小麦粉为主要糖源, 制成5组等氮等脂等能的配合饲料。UF、FPH和RF组分别添加5.4%、5.5%及5.5%三种不同分子量水解鱼蛋白, 占饲料总蛋白大约 10%, 鱼粉添加量均为 18%, 饲料中植物蛋白占总蛋白均为大约68%; PP组为不添加水解蛋白组, 鱼粉添加量为18%, 饲料中植物蛋白占总蛋白大约 78%; 对照组(FM组)鱼粉为唯一蛋白源, 添加量为67.5%。饲料配方及化学组成见表3。
将所有原料粉碎过80目筛网, 按照配方充分混合, 制成粒径为3 mm的饲料, 然后放入60℃烘箱内烘12h, 在–20℃下保存备用。
1.3 实验动物及饲养管理
实验选用初始体重为(16.05±0.03) g大菱鲆幼鱼,购自青岛通用水产养殖有限公司(中国, 青岛), 养殖实验在烟台开发区天源水产有限公司(中国, 烟台)进行。在实验前, 先用自制的对照组饲料饲喂两周, 使其适应养殖环境和饲料的硬度、粒径。实验采用自然光, 在养殖水桶(水体体积 120 L)内养殖,养殖用海水为深井海水, 采用流水养殖模式, 流速5 L/min, 水温(15±0.5) , ℃ 溶氧高于 7 mg/L, 盐度32 g/L, pH 7.5—8.0。
在实验开始前, 停食24h, 选择大小均匀、体格健壮且体表无病的大菱鲆幼鱼。随机将鱼分于15个养殖桶内(5个处理组, 每个处理3个重复), 每桶25尾; 每天人工饱食投喂两次(06:30和16:30), 投饲结束 30min后, 对每个桶内的残饵计数, 根据每 100粒残饵的平均重量, 来计算未摄食的残饵重量。养殖实验共计进行 68d, 期间鱼生长良好, 没有出现发病或死亡。
1.4 样品采集及生化分析
实验开始前, 随机取20尾大菱鲆, 用于常规分析; 在实验过程中, 采用虹吸法收集粪便; 实验结束后, 停食 5h, 开始收集大菱鲆。分别称量每桶鱼的总重量, 记录鱼的尾数, 计算每桶鱼的平均重量。
原料、饲料和鱼体均在 105℃烘干至恒重求得干物质重量; 粗蛋白的测定用凯氏定氮法(VELP, UDK142 automatic distillation unit,VELP, Usmate, MB, 意大利); 粗脂肪的测定用索氏抽提法(Foss Tecator, Hoganas, 瑞典), 以石油醚为抽提液进行抽提; 灰分的测定先将样品在电炉上炭化, 再用马福炉 550℃灼烧 5h, 失重法测得; 能量的测定用弹式热量仪(6100 Compensated Jacket Calorimeter; Parr Instrument Company, Moline, IL, 美国)测得; 钇元素的测定在高氯酸水解后用等离子发射光谱仪(ICP-OES, Vista-MPX, Varian, 美国)测得, 表观消化率的测定和计算按 Sugiura等[15]的方法进行; 总磷和钙的测定在高氯酸水解后用等离子发射光谱仪(ICP-OES, Vista-MPX, Varian, 美国)测得。
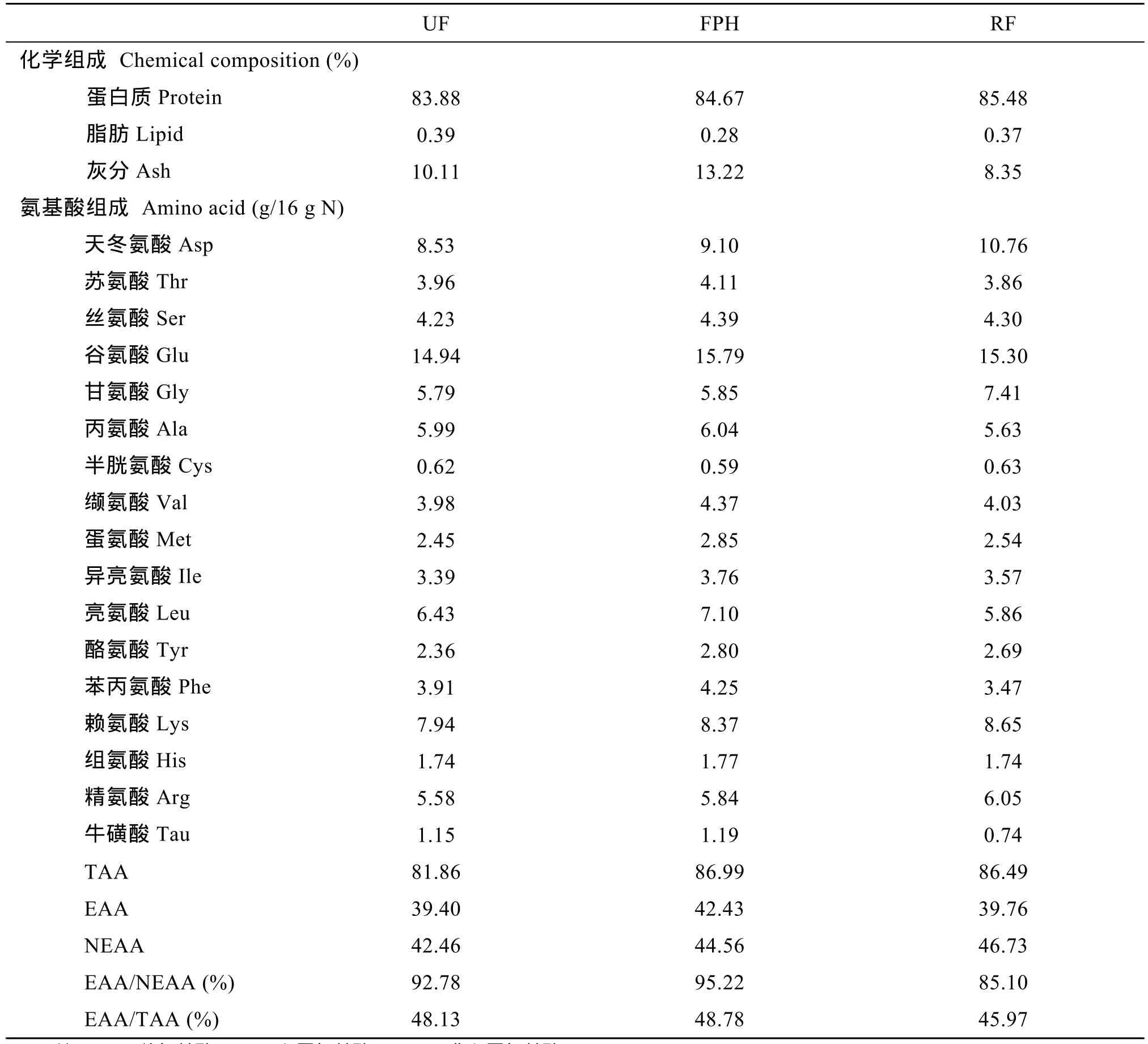
表2 UF、FPH 和RF 三种水解鱼蛋白的化学组成和氨基酸组成Tab. 2 Chemical composition and amino acid composition of UF, FPH and RF
氨基酸和牛磺酸的测定在6 mol/L HCl中110℃水解22—24h, 采用日立8900全自动氨基酸测定仪(Hitachi L-8900 automatic amino acid analyzer, Hitachi, 日本)测得; 饲料水解度的表示采用游离 α-氨基氮与总α-氨基氮的比, 其中游离α-氨基氮采用三硝基苯磺酸测定法[16], 总 α-氨基氮用 6 mol/L HCl中110℃水解22—24h后测得。
1.5 计算及统计分析方法
大菱鲆的成活率、摄食率、特定生长率、饲料效率、蛋白质沉积率、蛋白质效率和营养物质的表观消化率参考以下公式:


表观消化率(Apparent digestibility coefficient, ADC, %)= [1–(饲料中 Y2O3含量×粪便中的营养物质)/(饲料中的营养物质 × 粪便中 Y2O3的含量)] ×100其中, N1、N0表示实验前每桶鱼尾数和试验后鱼每桶尾数, W、W0和W1分别表示每尾鱼摄食的饲料干物质重(g)、实验开始时鱼体重(g)和实验结束时鱼体重(g), P、P0和P1表示饲料粗蛋白含量(干重%)、实验开始时鱼体粗蛋白含量(湿重%)和实验结束时鱼体粗蛋白含量(湿重%), t表示养殖实验天数(d)。

表3 实验饲料配方及化学组成(% 干物质)Tab. 3 Formulation and chemical composition of experimental diets (% in dry matter)
所有实验数据均用平均值±标准误表示。采用SPSS 16.0进行单因素方差分析(One-way ANOVA),当差异达到显著水平(P<0.05), 进行邓肯多重比较(Duncan’s multiple range tests)。
2 结果
2.1 三种水解鱼蛋白的分子量分布、化学组成、氨基酸组成及饲料的氨基酸组成
UF、FPH和RF的分子量分布见表1。三种水解鱼蛋白的分子量主要分布在 0—5000 D, 均达到99.9%, 其主要的差异在于0—5000 D不同肽段所占的比例不同, UF在200—500 D(主要为二肽和三肽)之间占的比例为60.6%, 高于FPH的54.9%和RF的46.3%, 但UF在500—1000 D的比例仅占22.7%, 低于FPH的26.1%和RF的26.4%。RF在1000—5000 D比例最高, 是UF和FPH的大约2倍, UF比例最低, 不到10%。
UF、FPH和RF的化学组成和氨基酸组成见表2。三种水解鱼蛋白的蛋白含量均超过80%, RF最高为85.48%, 脂肪含量均低于1%; 每100 g蛋白中, FPH和RF总氨基酸含量相近, 分别为86.99 g/16 g N和86.46 g/16 g N, 高于UF的81.86 g/16 g N, 总必需氨基酸含量FPH最高为42.43 g/16 g N, RF的总非必需氨基酸含量最高为46.73 g/16 g N; 牛磺酸含量UF和FPH组含量相近, 分别为1.15 g/16 g N和1.19 g/16 g N, 高于RF组的0.74 g/16 g N。
实验饲料的氨基酸组成见表4。FM组每100 g蛋白中总氨基酸含量最低为79.02 g/16 g N, 在添加水解鱼蛋白的3组中, 含量均超过85 g/16 g N, 分别为86.60 g/16 g N(UF)、88.77 g/16 g N(FPH) 和87.36 g/16 g N(RF), PP组为84.85 g/16 g N; 每100 g蛋白中的总必需氨基酸各处理组含量相近为(38.45—41.27) g/16 g N, 非必需氨基酸在UF、FPH及RF组含量较高, 均超过45 g/16 g N; FM组的必需氨基酸(EAA)与非必需氨基酸(NEAA)比值最高,为 94.79%, 牛磺酸含量也最高, 是 UF、FPH、RF及PP组含量的2倍多。
2.2 不同饲料处理对大菱鲆幼鱼生长及饲料利用的影响
不同饲料处理对大菱鲆幼鱼生长及饲料利用的影响见表5。在68d的生长实验中, 所有处理组的大菱鲆没有出现生病或死亡的情况, 存活率均为100%; FM 组特定生长率显著高于 RF组(P<0.05),与FPH、UF及PP组无显著性差异(P>0.05), RF组与FPH、UF及PP组也无显著性差异(P>0.05); 大菱鲆幼鱼的终末体重与特定生长率有相似的变化趋势。摄食率在各处理组之间无显著性差异(P>0.05);饲料效率、蛋白质效率和蛋白质沉积率在各处理组之间有相同的变化趋势, 即FM组显著高于FPH、RF及PP组(P<0.05), 但与UF无显著差异(P>0.05), UF组与FPH、RF及PP组差异也不显著(P>0.05)。

表4 实验饲料的氨基酸组成(g/16 g N)Tab. 4 The amino acid composition of experimental diets (g/16 g N)
2.3 不同饲料处理对大菱鲆幼鱼体组成的影响
不同饲料处理对大菱鲆幼鱼鱼体化学成分的影响见表 6。在饲料中添加不同分子量水解鱼蛋白显著影响大菱鲆鱼体的水分、粗蛋白和灰分含量, 水分含量在 UF、FPH、RF及 PP组之间无显著差异(P<0.05), 但在FM组显著低于FPH组(P<0.05); 粗蛋白含量在 FM组中显著高于 FPH组(P<0.05), 但FPH组与UF、RF及PP组差异不显著(P>0.05); 灰分含量在 FM组含量最高, 显著高于 UF、FPH和RF组(P<0.05), 但与PP组差异不显著(P>0.05)。
不同饲料处理对鱼体氨基酸组成的影响见表7。结果显示, 测得的16种鱼体氨基酸有14种氨基酸含量差异不显著(P>0.05), 仅有组氨酸和精氨酸在各处理组之间产生显著的变化(P<0.05); 鱼体牛磺酸含量在FM组是其他实验组的2倍多, 达到(1.78± 0.03) g/16 g N, FPH组的鱼体牛磺酸含量也较高,显著高于PP组(P<0.05), 但与UF和RF组差异不显著(P>0.05)。

表5 不同分子量水解鱼蛋白对大菱鲆幼鱼生长和饲料利用的影响(平均值±标准误)Tab. 5 The effects of different molecular weight fish protein hydrolysate on growth performance and feed utilization of juvenile turbot (mean ± SE)

表6 不同分子量水解鱼蛋白对大菱鲆幼鱼体成分的影响(平均值±标准误)Tab. 6 The effects of different molecular weight fish protein hydrolysate on body composition of juvenile turbot (mean ± SE)

表7 不同分子量水解鱼蛋白对大菱鲆幼鱼鱼体氨基酸组成的影响(g/16 g N)(平均值±标准误)Tab. 7 The effects of different molecular weight fish protein hydrolysate on amino acid composition of juvenile turbot (g/16 g N) (mean ± SE)
2.4 不同饲料处理对大菱鲆幼鱼饲料消化率的影响
不同处理对大菱鲆幼鱼饲料干物质和蛋白质表观消化率的影响见表 8。饲料中添加水解蛋白显著影响饲料的消化率, 摄食UF、FPH及RF组的大菱鲆饲料干物质和蛋白质表观消化率都显著高于 PP组(P>0.05), 但显著低于 FM 组(P<0.05), 干物质和蛋白质表观消化率在UF组显著高于FPH和RF组(P<0.05), 但FPH和RF组差异不显著(P>0.05)。
不同饲料处理对大菱鲆幼鱼饲料氨基酸消化率的影响见表9。实验所测得16种饲料氨基酸和牛磺酸的消化率在不同处理间均产生显著影响(P<0.05), FM 组中除天冬氨酸、甘氨酸、半胱氨酸、酪氨酸和精氨酸外, 其余氨基酸的消化率均是所有处理组中最高的; UF组的氨基酸消化率除半胱氨酸外均高于FPH、RF及PP组, 并且UF组天冬氨酸、苏氨酸、丝氨酸、谷氨酸、甘氨酸、丙氨酸、缬氨酸和蛋氨酸消化率与FPH、RF及PP组3组相比差异显著(P<0.05), 甚至天冬氨酸、甘氨酸、酪氨酸和精氨酸的消化率在UF组中高于FM组; FPH组与RF组之间的氨基酸消化率仅有蛋氨酸和半胱氨酸有显著差异(P<0.05); PP组饲料氨基酸消化率除半胱氨酸、蛋氨酸、亮氨酸和酪氨酸外, 12种饲料氨基酸的消化率全都显著低于添加水解蛋白组和 FM 组(P<0.05), 并且PP组中亮氨酸和酪氨酸的消化率也是所有处理组中最低的, 但没有产生显著性差异(P>0.05); 含硫氨基酸半胱氨酸和牛磺酸在含水解鱼蛋白组(UF组、FPH组和RF组)与不含水解鱼蛋白组(FM组和 PP组)的消化率有相反的趋势, 即牛磺酸在FM和PP组消化率显著高于UF、FPH及RF组(P<0.05), 但半胱氨酸的消化率则在FM和PP组显著低于UF、FPH及RF组, 另一含硫氨基酸蛋氨酸在FM和UF组显著高于其他组(P<0.05)。
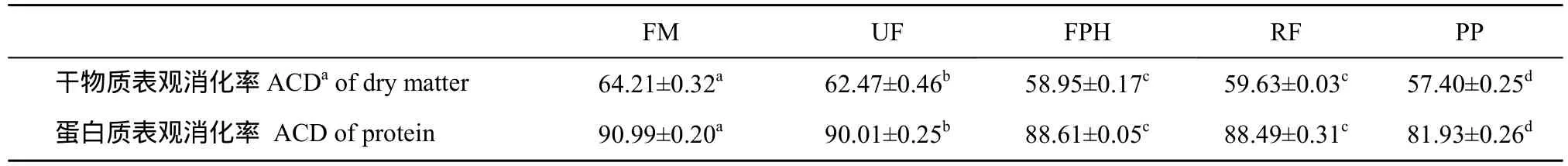
表8 大菱鲆幼鱼对饲料干物质和蛋白质表观消化率的影响(%)(平均值±标准误)Tab. 8 Apparent digestibility coefficients of dietary dry matter and protein in juvenile turbot (%) (mean ± SE)

表9 大菱鲆幼鱼对饲料氨基酸的表观消化率(%)(平均值±标准误)Tab. 9 Apparent digestibility coefficients of dietary amino acids in juvenile turbot (%) (mean ± SE)
3 讨论
3.1 添加不同分子量水解鱼蛋白对大菱鲆幼鱼生长性能的影响
目前, 饲料中添加水解鱼蛋白对肉食性鱼类生长的研究表明, 当饲料中水解鱼蛋白的添加量较低时能够促进鱼类的生长, 但当含量过高时又会对鱼类的生长起抑制作用[8,9,17]。另外, 在水解鱼蛋白对大菱鲆生长作用的研究中, 虽然Oliva-Teles等[18]报道在以标准鱼粉为唯一蛋白源的饲料中添加水解鱼蛋白对大菱鲆幼鱼的生长和饲料利用无促进作用, 但是 Zheng等[19]的研究发现, 在高植物蛋白饲料中适量添加水解鱼蛋白能够提高大菱鲆幼鱼的生长、饲料利用和免疫力。在本实验中, 添加低分子量的UF组和添加中等分子量的FPH组的末重和特定生长率与以鱼粉为唯一蛋白源的 FM 组无显著差异(P<0.05), 而在饲料利用率方面, 添加的水解鱼蛋白, 特别是低分子量水解鱼蛋白UF组, 在摄食率、饲料效率、蛋白质效率和蛋白质沉积率方面也均与FM组无显著差异(P<0.05)。这些表明, 水解鱼蛋白的添加, 特别是经过超滤膜超滤的富含更高含量的(200—500) D(二肽、三肽)低分子量水解鱼蛋白UF,在一定程度上可能促进大菱鲆幼鱼的生长和提高大菱鲆对高植物蛋白饲料的利用效率, 而 Oliva-Teles等[18]以鱼粉为唯一蛋白源, 用5%、10%和15%水解鱼蛋白替代鱼粉的饲料饲喂大菱鲆中没有发现明显的促进作用, 可能的原因一方面是饲料中鱼粉的含量较高, 海洋性蛋白源具有的特定营养成分已经能够满足大菱鲆生长的需要, 因此水解鱼蛋白对大菱鲆的有益作用被掩盖, 另一方面也与添加水解鱼蛋白的分子量有关。后一种原因在本实验中也得到了验证, 添加高分子RF的饲料饲喂大菱鲆幼鱼, 特定生长率甚至低于 PP组, 尽管没产生显著差异(P>0.05)。
在本实验中, 我们还发现PP组鱼粉含量虽然仅有18%, 但是仍然与以鱼粉为唯一蛋白源的FM组特定生长率无显著差异(P>0.05), 这与Fournier等[20]的研究结果相似。将PP组必需氨基酸含量与FM组比较发现, 除赖氨酸和蛋氨酸外, 其余均无明显降低, 且与 Kaushik[21]对大菱鲆所有必需氨基酸需求量的估计相比, PP组饲料也仅有赖氨酸和蛋氨酸含量稍微不足, 低于实际需求量大约 0.1 g/16 g N, 其余均满足需求, 因此含有比较平衡的必需氨基酸,这可能是摄食 PP组饲料的大菱鲆有较高的特定生长率的其中一个原因, 另外本实验的养殖周期仅为68d, 鱼体末重也只有初重的1到2倍, 在短期的养殖中大菱鲆可能对植物蛋白有较高的耐受能力, 这些也都可能造成PP组具有较高的特定生长率。但是PP组的饲料效率、蛋白质效率和蛋白质沉积率不仅显著的低于 FM 组(P<0.05), 而且也低于 UF组和FPH组, 因此如果进行长期的养殖实验, PP组可能会影响大菱鲆的生长。
3.2 添加不同分子量水解鱼蛋白的饲料对大菱鲆鱼体氨基酸组成的影响
本实验中鱼蛋白水解物添加到以高植物蛋白为主要蛋白源的饲料中, EAA/NEAA的比例低于以鱼粉为唯一蛋白源的饲料, 这与Espe等[11]的结果相似,并且进一步分析发现, 与 FM 组饲料相比, 添加水解鱼蛋白的UF、FPH及RF组主要是增加了每100 g饲料蛋白中非必需氨基酸的含量, 每 100 g蛋白所含有的必需氨基酸含量则相近, 并且 UF、FPH及RF组饲料必需氨基酸的含量均高于Kaushik[21]估计的大菱鲆对必需氨基酸的需求量, 另外, 在本实验所测的16种氨基酸中, 仅有精氨酸和组氨酸含量在鱼体内有显著的变化, 其余均无显著变化, 这些表明, 当鱼粉含量为 18%的条件下, 添加总蛋白大约10%的 UF、FPH及RF三种分子量的水解鱼蛋白到饲料中, 大部分的必需氨基酸含量可能已经能满足大菱鲆对饲料中必需氨基酸的需要, 因而在添加水解鱼蛋白的处理组并未引起鱼体大部分氨基酸含量的显著变化。但是发现鱼体牛磺酸含量在所有实验组中显著低于FM组, Qi等[22]报道饲料中的牛磺酸在体重为(6.3±0.01) g和(165.9±5.01) g的大菱鲆的需要量分别为1.15%和0.64%, 在本实验仅FM组牛磺酸含量能满足要求为1.2 g/16 g N, 其他各组只有(0.35—0.47) g/16g N, 因而, 实验组的鱼体牛磺酸含量显著低于FM组可能是饲料中牛磺酸含量较低所致。
3.3 添加不同分子量水解鱼蛋白对大菱鲆幼鱼消化率的影响
在植物蛋白占大约68%的饲料中添加不同分子量水解鱼蛋白后, 一个非常有趣的实验结果是大菱鲆幼鱼对饲料干物质和蛋白质的表观消化率相对于PP组有显著提高(P<0.05), 且含低分子量水解鱼蛋白的UF组消化率显著优于FPH和RF组(P<0.05),这点与本实验的预期相符合, 表明饲料中添加水解鱼蛋白能够改善大菱鲆幼鱼对高植物蛋白饲料的消化, 这一点在对氨基酸消化率的分析中的到进一步验证。
本实验所测得的不同饲料处理的 16种氨基酸和牛磺酸的表观消化率均产生显著的差异(P<0.05),表明在高植物蛋白饲料中添加不同分子量的水解鱼蛋白能够显著影响饲料氨基酸的表观消化率。在以鱼粉为唯一蛋白源的 FM 组中, 除天冬氨酸、甘氨酸、胱氨酸、酪氨酸和精氨酸外, 其余的氨基酸消化率均是各组中含量最高的, 这点证明鱼粉作为肉食性鱼类的蛋白源, 具有很高的消化率。同时, 我们也发现在3种不同分子量的水解鱼蛋白添加到含鱼粉 18%的饲料中后, 饲料氨基酸的消化率, 相对于含相同鱼粉的植物蛋白(PP)组有了显著提高, 甚至天冬氨酸、甘氨酸、酪氨酸和精氨酸的消化率 UF组还高于FM组。此外, 在3种添加水解鱼蛋白的饲料中, UF组的氨基酸消化率明显优于FPH和RF组, 天冬氨酸、苏氨酸、丝氨酸、蛋氨酸、谷氨酸、甘氨酸、亮氨酸和苯丙氨酸这8种氨基酸的消化率均高于FPH和RF组, 并且天冬氨酸、苏氨酸、丝氨酸、蛋氨酸、谷氨酸这5种氨基酸消化率甚至产生显著差异(P<0.05), 由于 UF中包含较高含量的(200—500) D的小肽, 因而可能是提高氨基酸消化率的重要原因之一, 这也部分解释水解蛋白中的低分子成分对促进肉食性鱼类生长起主要作用[9,10,23,24,]的原因, 而本实验的生长结果反映UF组和FPH与PP组生长性能无显著差异, 可能是由于一方面大菱鲆对植物蛋白的耐受力较高, 另一方面 68d的养殖时间较短, 低分子水解鱼蛋白添加到高植物蛋白饲料后在消化率上的优势作用没有充分体现出来, 可能随着养殖时间的增加, UF和FPH的添加会对大菱鲆幼鱼产生明显的促进作用。
本实验还发现, 添加水解鱼蛋白后饲料中牛磺酸的消化率显著低于不添加水解鱼蛋白饲料的牛磺酸消化率(P<0.05), 并且这与饲料中的植物蛋白含量高低无关, 而与牛磺酸合成密切相关的半胱氨酸的消化率正好与之相反, 在添加水解鱼蛋白的UF、FPH及RF组半胱氨酸的的消化率显著高于不添加水解鱼蛋白的FM和PP组(P<0.05)。以前的研究表明, 半胱氨酸是牛磺酸合成的前体物质, 主要来源于食物或者 β-丙氨酸和蛋氨酸, 受半胱胺双加氧酶的调节[25—27], 因此, 半胱氨酸在添加水解鱼蛋白的实验组中消化率提高可能是鱼体通过提高半胱氨酸的消化率增加对半胱氨酸的吸收, 以便更多的半胱氨酸转化成牛磺酸, 从而弥补牛磺酸在添加水解鱼蛋白饲料组中消化率过低导致的机体中牛磺酸需要量的不足, 但是在高植物蛋白中添加水解鱼蛋白为什么会导致牛磺酸消化率降低, 原因尚不清楚。
4 结论
在高植物蛋白饲料中添加低分子水解鱼蛋白(UF组), 大菱鲆幼鱼的生长和饲料利用有升高的趋势, 鱼体氨基酸组成无显著影响, 但UF、FPH以及RF的添加都显著促进大菱鲆对饲料干物质、蛋白质和氨基酸的表观消化率, 且低分子 UF的作用优于FPH和高分子RF。此外, 大菱鲆似乎对含不同分子量水解鱼蛋白的高植物蛋白饲料中牛磺酸消化率都有降低的趋势, 其原因还有待进一步研究。
[1] FAO. The State of World Fisheries and Aquaculture [M]. Rome: FAO Press. 2012, 24
[2] Tacon A G J, Metian M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects [J]. Aquaculture, 2008, 285(1—4): 146—158
[3] Collins S A, Desai A R, Mansfield G S, et al. The effect of increasing inclusion rates of soybean, pea and canola meals and their protein concentrates on the growth of rainbow trout: Concepts in diet formulation and experimental design for ingredient evaluation [J]. Aquaculture, 2012, 344—349: 90—99
[4] Snyder G S, Gaylord T G, Frederic T, et al. Effects of carnosine supplementation to an all-plant protein diet for rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 2012, 338—341: 72—81
[5] Hardy R W, Higgs D A, Lall S P, et al. Alternative Dietary Protein and Lipid Sources for Sustainable Production of Salmonids [M]. Havforskningsinstituttet. 2001, 44
[6] Liaset B, Lied E, Espe M. Enzymatic hydrolysis of by-products from the fish-filleting industry; chemical characterization and nutritional evaluation [J]. Journal of the Science of Food and Agriculture, 2000, 80(5): 581—589
[7] Mach D T N, Nortvedt R. Free amino acid distribution in plasma and liver of juvenile cabia (Rachycentron canadum) fed increased levels of lizardfish silage [J]. Aquaculture Nutrition, 2011, 17(2): 644—656
[8] Espe M, Sveier H, Hogoy I, et al. Nutrient absorption andgrowth of Atlantic salmon (Salmo salar L.) fed fish protein concentrate [J]. Aquaculture, 1999, 174(1—2): 119—137
[9] Zheng K, Liang M, Yao H, et al. Effect of dietary fish protein hydrolysate on growth, feed utilization and IGF-I levels of Japanese flounder (Paralichthys olivaceus) [J]. Aquaculture Nutrition, 2012, 18(3): 297—303
[10] Kousoulaki K, Albrektsen S, Langmyhr E, et al. The water soluble fraction in fish meal (stickwater) stimulates growth in Atlantic salmon (Salmo salar L.) given high plant protein diets [J]. Aquaculture, 2009, 289(1): 74—83
[11] Espe M, Ruohonen K, El-Mowafi A. Hydrolysed fish protein concentrate (FPC) reduces viscera mass in Atlantic salmon (Salmo salar) fed plant-protein-based diets [J]. Aquaculture Nutrition, 2012, 18(6): 599—609
[12] Kousoulaki K, Olsen H J, Albrektsen S, et al. High growth rates in Atlantic salmon (Salmo salar L.) fed 7.5% fish meal in the diet. Micro-, ultra- and nano-filtration of stickwater and effects of different fractions and compounds on pellet quality and fish performance [J]. Aquaculture, 2012, 338—341: 134—146
[13] Zheng K K, Liang M Q, Yao H B, et al. Inclusion of size-fractionated fish protein hydrolysate in high plant protein diets for Japanese flounder, Paralichthys olivaceus [J]. Acta Hydrobiologica Sinica, 2011, 35(5): 829—834 [郑珂珂, 梁萌青, 姚洪波, 等. 在高植物蛋白饲料中添加水解鱼蛋白对牙鲆幼鱼的影响. 水生生物学报, 2011, 35(5): 829—834]
[14] Luo Y W, Ai Q H, Mai K S, et al. Effects of dietary rapeseed meal on growth performance, digestion and protein metabolism in relation to gene expression of juvenile cobia (Rachycentron canadum) [J]. Aquaculture, 2012, 368—369: 109—116
[15] Sugiura S H, Dong F M, Rathbone C K, et al. Apparent protein digestibility and mineral availabilities in various feed ingredients for salmonid feeds [J]. Aquaculture, 1998, 159(3—4): 177—202
[16] Adler-Nissen J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid [J]. Journal of Agricultural and Food Chemistry, 1979, 27(6): 1256—1262
[17] Liu X D, Liang M Q, Zhang L M, et al. Effect of fish protein hydrolysate levels on growth performance and biological and physiological parameters in tongue sole post-larvae [J]. Acta Hydrobiologica Sinica, 2010, 34(2): 242—249 [柳旭东, 梁萌青, 张利民, 等. 饲料中添加水解鱼蛋白对半滑舌鳎稚鱼生长及生理生化指标的影响. 水生生物学报, 2010, 34(2): 242—249]
[18] Olivia-Teles A, Cerqueira A L, Goncalves P. The utilization of diets containing fish protein hydrolysate by turbot (Scophtalmus maximus) juveniles [J]. Aquaculture, 1999, 179(1): 195—201
[19] Zheng, K, Liang M, Yao H, et al. Effect of size-fractionated fish protein hydrolysate on growth and feed utilization of turbot (Scophthalmus maximus L.) [J]. Aquaculture Research, 2013, 44(6): 895—902
[20] Fournier V, Huelvan C, Desbruyeres E. Incorporation of a mixture of plant feedstuffs as substitute for fish meal in diets of juvenile turbot (Psetta maxima) [J]. Aquaculture, 2004, 236(1): 451—465
[21] Kaushik S J. Whole body amino acid composition of European seabass (Dicentrarchus labrax), gilthead seabream (Sparus aurata) and turbot (Psetta maxima) with an estimation of their IAA requirement profiles [J]. Aquatic Living Resources, 1998, 11: 355—358.
[22] Qi G, Ai Q H, Mai K S, et al. Effects of dietary taurine supplementation to a casein-based diet on growth performance and taurine distribution in two sizes of juvenile turbot (Scophthalmus maximus L.) [J]. Aquaculture, 2012, 358—359: 122—128
[23] Aksnes A, Hope B, Høstmark Ø, et al. Inclusion level of size fractionated fish hydrolysate in high plant protein diets for Atlantic cod, Gadus morhua [J]. Aquaculture, 2006b, 261(3): 1102—1110
[24] Aksnes A, Hope B, Jönsson E, et al. Size-fractionated fish hydrolysate as feed ingredient for rainbow trout (Oncorhynchus mykiss) fed high plant protein diets. I: growth, growth regulation and feed utilization [J]. Aquaculture, 2006c, 261(1): 305—317
[25] Goto T, Tiba K, Sakurada Y, et al. Determination of hepatic cysteinesulfinate decarboxylase activity in fish by means of OPA-prelabeling and reverse-phase high-performance liquid chromatographic separation [J]. Fisheries Science, 2001, 67(3): 553—555
[26] Goto T, Matsumoto T, Murakami S. Conversion of cysteate into taurine in liver of fish [J]. Fisheries Science, 2003, 69(1): 2l6—2l8
[27] Kuzmina V V, Gavrovskaya L K, Ryzhova O V. Taurine. Effect on exotrophia and metabolism in mammals and fish [J]. Journal of Evolutionary Biochemistry and Physiology, 2012, 46(1): 19—27
THE EFFECTS OF FISH PROTEIN HYDROLYSATE ON THE DIGESTIBILITY OF JUVENILE TURBOT (SCOPHTHALMUS MAXIMUS L.)
WEI Yu-Liang1,2, LIANG Meng-Qing2, ZHENG Ke-Ke2and WANG Xin-Xing2
(1. The Key Laboratory of Mariculture (Education Ministry of China), Ocean University of China, Qingdao 266003, China; 2. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China)
In this study, we investigated whether and how the molecular weight of fish protein hydrolysate in the plant-protein diets would affect the digestive ability of juvenile turbot (Scophthalmus maximus L). The 68-day experiments were conducted on fish with initial weight of (16.05±0.03) g. Five groups of diets (UF, FPH, RF, PP, and FM) were formulated to be isolipidic, isonitrogenous and isoenergetic. UF (ultrafiltered fish protein hydrolysate), FPH (fish protein hydrolysate) and RF (retentate fish protein hydrolysate) contained 5.4%, 5.5% and 5.5% of protein hydrolysates respectively (10% of total dietary protein). PP contained zero protein hydrolysate. Fish meal consisted of 18% of the diets in UF, FPH, RF, and PP, and it was the sole protein source in the control diet (FM). The special growth rates of fish fed with UF (UF fish), FPH and PP were not significantly different from FM fish (P>0.05). There was also no difference in feed efficiency (FE), protein efficiency ratio (PER), or protein productive value (PPV) between UF fish and FM fish (P>0.05). However FPH fish, RF fish, and PP fish displayed significantly lower FE, PER and PPV than FM fish did (P<0.05). The apparent digestibility coefficients (ADC) for the dry matter and protein of UF, FPH and RF fish were significantly higher than that of PP fish (P<0.05), but lower than that of FM fish (P<0.05). Between UF, FPH, and RF fish, the ADC for the dry matter and protein of UF was significantly higher than that of the latter two (P<0.05). The ADCs for 16 amino acids and taurine could be obviously affected by different dietary treatments (P<0.05). For most amino acids the ADC was the highest in FM fish and the lowest in PP fish among all the groups. The ADCs for cysteine and taurine exhibited opposite patterns between the fish protein hydrolysates-containing groups (UF, FPH and RF) and the fish protein hydrolysates-lacking groups (FM and PP). In conclusion, addition of UF and FPH to the high plant protein diets could help increase the growth and feed utilization of the juvenile turbot; the ACD for dry matter, protein, and amino acids could be improved with FPH and UF; compared to FPH and RF, the low molecular weight UF could be more effective to improve the growth. However, it seemed that the fish protein hydrolysate might reduce the ACD for taurine.
Turbot; Fish protein hydrolysate; Growth; Feed utilization; Apparent digestibility coefficient
S963
A
1000-3207(2014)05-0910-11
10.7541/2014.136
2013-05-10;
2013-12-07
公益性行业专项(201303053); 国家自然科学基金项目(31340076, 31172423)资助
卫育良(1989—), 男, 山西省晋城人, 硕士; 主要从事水产动物营养与饲料学研究。E-mail: happyweiyl@sina.cn
梁萌青(1963—), 女, 研究员; Tel: 0532-85822914; E-mail: liangmq@ysfri.ac.cn
