夏季蓝藻水华期间太湖河口区和敞水区纤毛虫群落组成及水平分布
2014-03-17静卢文轩张雷燕陈非洲
李 静卢文轩张雷燕陈非洲
(1. 中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008; 2. 安徽省农业科学院水产研究所, 合肥 230031)
夏季蓝藻水华期间太湖河口区和敞水区纤毛虫群落组成及水平分布
李 静1,2卢文轩2张雷燕1陈非洲1
(1. 中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 南京 210008; 2. 安徽省农业科学院水产研究所, 合肥 230031)
在太湖北部主要河口区及敞水区域采集纤毛虫样品, 并用定量蛋白银法进行分析。河口区设 10个采样点, 敞水区设14个采样点。调查中共检出3纲15目60属的105种纤毛虫, 种类最多的是钩刺目(21种), 其次是寡毛目(20种)。纤毛虫优势种及其在河口区和敞水区占总丰度比例分别为: 寡毛目的双叉弹跳虫Halteria bifurcate Tamar (12.3%、18.1%)、大弹跳虫H. grandinella Dujardin (12.3%、10.9%)、短列裂隙虫Rimostrombidium brachykinetum Krainer (8.0%、13.4%)、圆筒状似铃壳虫Tintinnopsis cylindrata Kofoid & Campbell (11.8%、4.5%)、盾纤目的银灰膜袋虫Cyclidium glaucoma Müller (3.1%、10.8%)。其他常见种类还有: 前口目的趣尾毛虫Urotricha farcta Claparède & Lachmann、寡毛目的杯铃壳虫Codonella cratera Leidy、小裂隙虫R. humile Penard、奇异急游虫Strombidium mirabile Penard、小筒壳虫Tintinnidium pusillum Entz、缘毛目的水生钟虫复合种Vorticella aquadulcis complex和钟形钟虫V. campanula Ehrenberg。河口区和敞水区纤毛虫平均丰度分别为31407 cells/L(范围1 600—80 900 cells/L)和18618 cells/L(范围1225 —36000 cells/L),平均生物量分别为1322.6 μg/L(范围44.6—3119.7 μg/L)和543.6 μg/L(范围44.0—1570.4 μg/L)。两个区域的优势类群相似, 均以寡毛目、前口目、盾纤目和缘毛目为主。但河口区纤毛虫生物多样性(种类丰富度、Simpson多样性指数、Margalef多样性指数、Shannon多样性指数)显著高于敞水区的(P<0.05)。统计分析发现, 食藻种类与叶绿素 a含量之间无显著相关, 而食菌种类的丰度和叶绿素 a含量呈显著正相关关系(P<0.001)。由此推测, 纤毛虫群落结构的空间差异可能与水华暴发程度有较大关系。
蓝藻水华; 纤毛虫; 定量蛋白银法; 太湖; 水平分布
湖泊蓝藻水华频繁暴发, 会影响到生物多样性保育和区域生态系统平衡。自20世纪80年代以来,太湖水体富营养化加剧, 北部湖区频繁出现蓝藻水华[1,2], 如微囊藻属 Microcystis有时高达藻类总生物量的98%。有关水华蓝藻对水生生物类群的影响,已开展多方面研究, 例如鱼、虾等水产品中可能累积藻毒素从而影响食品安全, 且这种风险目前可能被低估; 大存量蓝藻可能导致浮游动物摄食和生长速度缓慢等[3,4]。也有研究表明, 水华期间部分浮游动物类群, 如纤毛虫和鞭毛虫, 数量急剧减少, 微食物网亦非常脆弱[5]。
太湖是大型浅水湖泊, 因水网稠密和风力作用等造成其水质和生态类型空间差异较大[6]。国内外学者对太湖流域水生态系统已开展了大量研究, 对浮游动物、浮游细菌、真核微型浮游生物等重要类群做了很多工作[7—9]。关于太湖浮游纤毛虫的调查主要在 20世纪, 如: 1951年白国栋于五里湖调查,检出纤毛虫 62种(属)[10]; 徐润林和 Nauwerck[11]对梅梁湾和胥口湾的纤毛虫进行染色分析, 共检出 18个种; 蔡后建[12]也对梅梁湾的纤毛虫进行过调查。但是近年来随着太湖营养盐水平的变化, 纤毛虫群落结构可能发生了较大变化。另外, 近河口区和敞水区水体性质差异较大, 而对这两种生境中纤毛虫的群落组成和水平分布差异尚不清楚。本次调查于2009年7月进行, 选择太湖北部主要入湖河口和敞水区采集样品并使用定量蛋白银法对纤毛虫进行分析。
1 材料与方法
1.1 研究区域和采样点位
2009年夏季蓝藻水华期间, 利用快艇在太湖北部进行取样, 采样点位(图1)如下: (1)在靠近重要港口、河流入湖口等区域设置了10个采样点E1—E10,采样点水深1—2 m; (2)在太湖北部敞水区域设置了14个采样点P1—P14, 水深1.5—3.1 m。河口区采样点按从E1到E10顺序依次对应: 鼋头渚、大溪河、望虞河、金墅港、铜坑港、山后尖浜、定跨港、大浦港、沙塘港、殷村港。

图1 太湖北部河口区和敞水区采样点Fig. 1 Sampling stations in estuary and pelagic zones of Northern Lake Taihu
1.2 样品采集及分析鉴定
用2.5 L采水器分别采集表层(水下50 cm)、中层、底层(底泥界面以上50 cm)湖水, 等体积混合后备用。水深不足1.5 m的, 则取表层和底层水等体积混合。取1 L混合水装入白色塑料瓶中, 用波恩氏液固定(终浓度5%), 用于分析纤毛虫。将部分混合水样装入容器中, 用保温箱保存并尽快带回实验室,以测定氮磷营养盐和叶绿素a浓度[13]。水体透明度用Secchi盘现场测定, 水温、pH等指标用YSI 6600水质分析仪现场测定。
纤毛虫样品用定量蛋白银法制片[14], 步骤如下:将固定后的样品静置 48h, 除去上清液浓缩至 50 mL保存; 将浓缩样品混匀并从中吸取 0.5—2 mL不等水样过滤至硝酸纤维膜; 经琼脂包埋、蛋白银染色、异丙醇和二甲苯梯度脱水、中性树胶封片完成制片,历时约6h。将标本风干后用Olympus BX51显微镜观察分析。纤毛虫的鉴定主要参照Foissner等[15]、Lynn[16]及其他相关文献。生物量的计算主要参照Foissner等[15], 个别没有生物量数据的种类则单独计算: 挑出一般不少于10只个体, 测量虫体长度、宽度和厚度, 按照近似几何体体积计算公式计算体积, 假定虫体比重为 1[17], 从而得出该种类单位个体平均生物量值。
1.3 数据分析

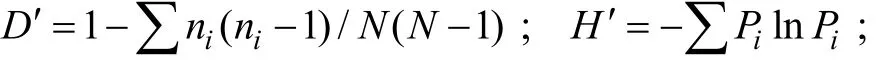
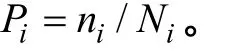
各点纤毛虫丰度和生物量的差异性, 及其与环境因子的相关关系用SPSS 16.0软件分析。
2 结果
2.1 湖区环境特征
采样期间, 水温变化范围为 27.4—30.9 , pH℃变化范围为7.2—9.3。水体平均总氮含量为3.4 mg/L,各点含量相差较大, 如金墅港口 E4采样点总氮为1.5 mg/L, 而梅梁湾北部敞水区 P2采样点则高达9.8 mg/L。总磷平均浓度0.20 mg/L, 其空间分布与总氮有相似格局。叶绿素a变化范围也较大, P9点为8.3 μg/L, 而梅梁湾P2点则高达491.2 μg/L。水体硝酸盐氮、亚硝酸盐氮、铵态氮浓度空间差异也较大(表1)。
2.2 纤毛虫群落组成和空间分布特征
在24个采样点共检出105种纤毛虫, 隶属于3纲、14目、60属。种类数最多的是钩刺目(21种), 其次是寡毛目(20种)(附录1)。河口区和敞水区的优势类群相似, 均以寡毛目(22914和11514 cells/L)、前口目(2342和1179 cells/L)、盾纤目(1976和2086 cells/L)和缘毛目(2395和2407 cells/L)为主, 在所有采样点这四个目纤毛虫丰度占总丰度比例均达 90%以上(表2)。河口区和敞水区的纤毛虫优势种丰度及其占总丰度比例分别为: 寡毛目的双叉弹跳虫 Halteria bifurcate Tamar (3877 cells/L, 12.3%; 3375 cells/L, 18.1%)、大弹跳虫H. grandinella Dujardin (3865 cells/L, 12.3%; 2021 cells/L, 10.9%)、短列裂隙虫 Rimostrom-bidium brachykinetum Krainer(2525 cells/L, 8.0%; 2500 cells/L, 13.4%)、圆筒状似铃壳虫Tintinnopsis cylindrata Kofoid & Campbell(3694 cells/L, 11.8%; 830 cells/L, 4.5%)、盾纤目的银灰膜袋虫Cyclidium glaucoma Müller(988 cells/L, 3.1%; 2017 cells/L, 10.8%), 这5个种共占纤毛虫总丰度的55.2%。其他常见种类还有: 趣尾毛虫Urotricha farcta Claparède & Lachmann、杯铃壳虫Codonella cratera Leidy、奇异急游虫 Strombidium mirabile Penard、小筒壳虫Tintinnidium pusillum Entz、水生钟虫复合种Vorticella aquadulcis complex和钟形钟虫V. campanula Ehrenberg,这些种类检出率均大于 50%。小裂隙虫 R. humile Penard虽然检出率不高, 但是在检出的样品中所占比重(丰度)最高达30%。河口区和敞水区主要食性功能群分布也相似, 均以食菌的最多, 其次是食藻的和营混合营养方式的类群(图2)。
河口区和敞水区纤毛虫平均丰度分别为31407 cells/L (范围 1600—80900 cells/L)和 18618 cells/L(范围1225—36000 cells/L), 平均生物量分别为 1322.6 μg/L (范围44.6—3119.7 μg/L)和543.6 μg/L(范围44.0—1570.4 μg/L)(表 2)。河口区的平均丰度和平均生物量均比敞水区高, 但是差异均不显著(P>0.05)。
虽然种类组成和主要功能群分布相似, 但是河口区的生物多样性指数如种类丰富度、Simpson多样性指数、Margalef多样性指数、Shannon多样性指数均显著高于敞水区(前三者 P<0.05, 后者P<0.01)(表2)。检出种类最多的是靠近定跨港的E7点位, 检出36种, 最少的是敞水区P8点, 仅检出8种。北太湖 Shannon多样性指数范围 1.4—2.9, Simpson多样性指数范围0.6—0.9, Margalef多样性指数范围0.9—4.2。
2.3 纤毛虫群落结构与环境因子的关系
相关性分析发现, 总氮、总磷和叶绿素a含量对纤毛虫群落结构影响较大(表3)。其中叶绿素a含量与纤毛虫总丰度显著相关(P<0.01), 而影响生物多样性指数的主要是硝酸盐氮和铵态氮。主要类群(目)中,缘毛类和盾纤类受总氮、总磷和叶绿素a含量影响较大, 而这些因子对寡毛类和前口类影响不显著。各食性功能群中, 主食细菌的、营混合营养方式的和捕食性的纤毛虫数量均与叶绿素 a含量呈显著正相关(P<0.01), 食藻的和杂食性的则受其影响较小。

表1 2009年7月北太湖敞水区和河口区环境特征(表中数值为平均值及其范围)Tab. 1 Environmental characteristics in the estuary and pelagic zones of Northern Lake Taihu in July 2009 (given as average value and its range)

表2 北太湖敞水区和河口区纤毛虫丰度、生物量和生物多样性(表中数值为平均值及其范围)Tab. 2 Environmental characteristics in the estuary and pelagic zones of Northern Lake Taihu (given as average value and its range)
3 讨论

图2 北太湖纤毛虫群落组成及主要食性功能群分布Fig. 2 Compositions of ciliate community and functional groups in Northern Lake Taihu
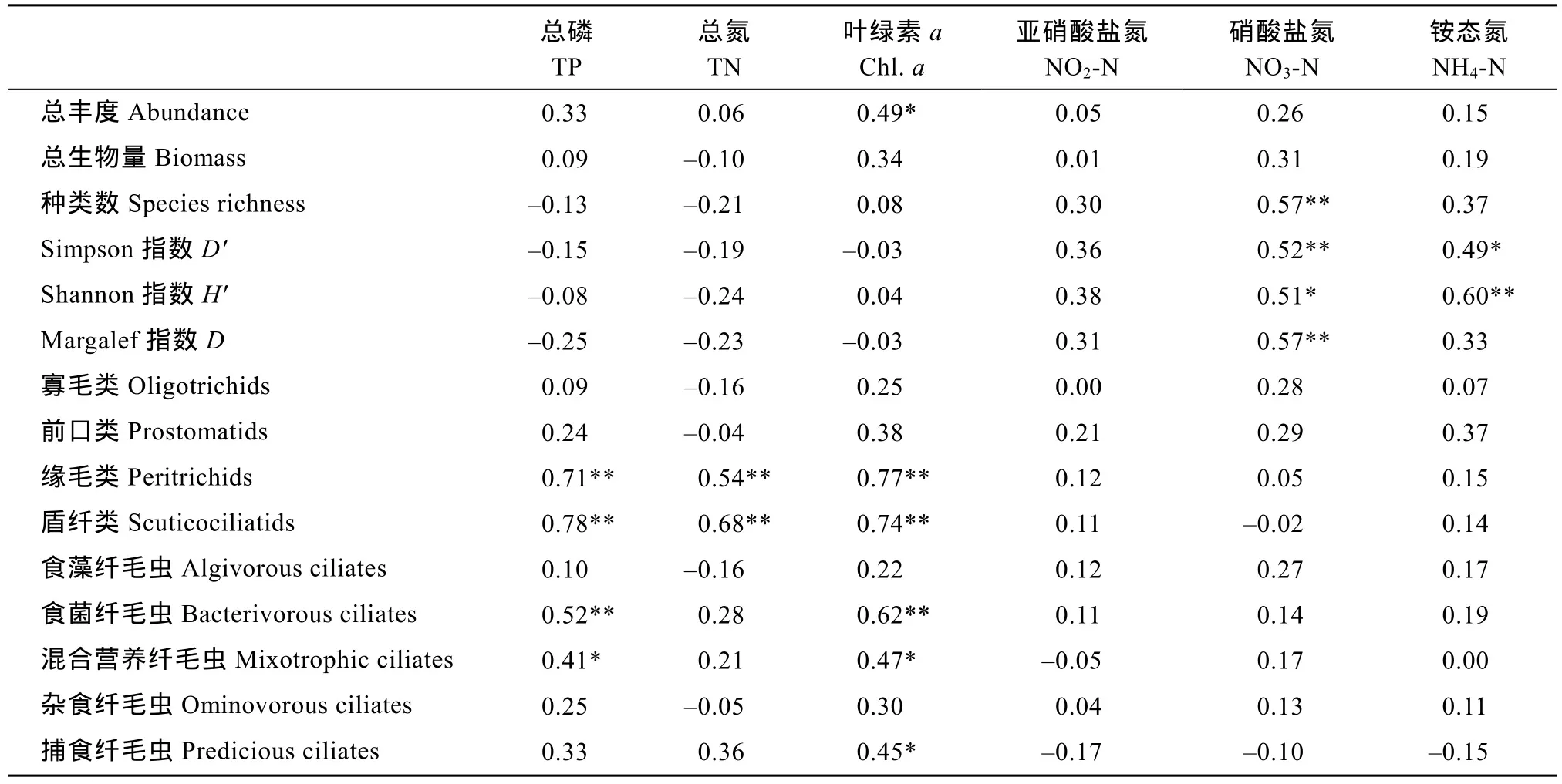
表3 纤毛虫总生物量、各类群丰度及其生物多样性指数和环境因子的相关系数Tab. 3 Correlation between environmental factors and abundance, biomass as well as biodiversity index of ciliates
本文主要目的是阐明大型富营养化湖泊太湖,在夏季水华期间的纤毛虫生物多样性, 及其在河口区和敞水区的群落组成和分布差异。结果显示太湖纤毛虫生物多样性较高, 本次在北太湖共检出超过100种纤毛虫。该结果高于已报道的一些其他湖泊,如位于Finland的温带富营养化湖泊Köyliönjärvi湖,该湖于水华期间检出 27种纤毛虫[18]; 还高于一些常年存在蓝藻水华的亚热带富营养化湖泊, 如滇池和星云湖[19]。这种生物多样性的季节和空间差异,尽管与采样范围、湖泊规模、水体环境差异、分析方法等有关, 但也说明蓝藻水华可能对浮游纤毛虫群落有不容忽视的影响, 而且该影响和水华暴发程度有关[5,20]。
虽然生物多样性差异较大, 但是本次得到的纤毛虫丰度和生物量与以往很多类似湖泊的结果相差不大[21]。本次检出种类中以寡毛目丰度最高, 比例达三分之二; 功能摄食类群上以食菌种类为主, 所占比例近 40%。相关性分析发现, 食菌种类的总丰度与水体叶绿素 a含量呈极显著正相关关系(P<0.01)。这可能是因为水华蓝藻暴发期间, 大量的藻类碎屑为细菌生长提供了有机碳源, 因而细菌大量滋生, 从而世代时间较短的小个体食菌纤毛虫也大量增殖[22]。植食性(食藻者)纤毛虫则没有呈现随叶绿素 a含量升高而增加的趋势, 究其原因可能主要有以下两个方面: 一是采样期间浮游植物以蓝藻为主, 而蓝藻不饱和脂肪酸含量低、常具群体胶鞘、存在化学防御、常产毒等, 因此普遍认为蓝藻不是浮游动物的理想食物来源[23]; 二是食藻纤毛虫还面临着与后生浮游动物竞争食物的情况[24]。
本次调查区域遍布北太湖敞水区和主要河流(出)入湖口, 纤毛虫群落结构存在明显空间差异。首先, 与敞水区相比, 河口区纤毛虫生物多样性和丰度、生物量均较高, 这种现象也存在于海洋中[25];其次, 两个区域的纤毛虫优势类群相似, 但是优势种及其优势度不尽相同。总体来说, 纤毛虫优势种主要是以细菌或藻类为主要食物的寡毛类, 如双叉弹跳虫H. bifurcate、大弹跳虫H. grandinella、短列裂隙虫R. brachykinetum、圆筒状似铃壳虫T. cylindrata等, 这些种类在两个区域均大量存在。但也有些种类空间分布差异明显, 如银灰膜袋虫 C. glaucoma主要是敞水区优势种, 而趣尾毛虫U. farcta、小裂隙虫R. humile、奇异急游虫S. mirabile等则主要出现在河口区水域。因此, 这些种类虽然在全球各种水体中广泛分布, 但其优势度的不同则代表了湖泊水环境的异质性[26]。
致谢:
感谢中国科学院太湖生态系统研究站提供太湖2009年7月理化数据。感谢李宽意、李珂、姚思鹏等协助采集样品。
[1] Duan H, Ma R, Xu X, et al. Two-decade reconstruction of algal blooms in China’s Lake Taihu [J]. Environmental Science & Technology, 2009, 43(10): 3522—3528
[2] Chen Y, Qin B, Teubner K, et al. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China [J]. Journal of Plankton Research, 2003, 25(1): 445—453
[3] Ibelings B W, Chorus I. Accumulation of cyanobacterial toxins in freshwater “seafood” and its consequences for public health: A review [J]. Environmental Pollution, 2007, 150(1): 177—192
[4] Ferrão-Filho A, Kowzlowsky-Suzuki B. Cyanotoxins: bioaccumulation and effects on aquatic animals [J]. Marine Drugs, 2011, 9: 2729—2772
[5] Moustaka-Gouni M, Vardaka E, Michaloudi E, et al. Plankton food web structure in a eutrophic polymictic lake with a history of tocic cyanobacterial blooms [J]. Limnology and Oceanography, 2006, 51(1, part 2): 715—727
[6] Qin B, Xu P, Wu Q, et al. Environmental issues of Lake Taihu, China [J]. Hydroboilogia, 2007, 581: 3—14
[7] Chen M, Chen F, Yu Y, et al. Genetic diversity of eukaryotic microorganisms in Lake Taihu, a large shallow subtropical lake in China [J]. Microbial Ecology, 2008, 56: 572—583
[8] Pang X H, Lü L Y, Niu Y, et al. The spatial Pattem of bacterioplankton community composition in Lake Taihu in Summer [J]. 2014, 38(2): 335—341 [庞兴红, 吕丽媛, 牛远,等. 太湖夏季浮游细菌群落多样性的空间格局. 水生生物学报, 2014, 38(2): 335—341]
[9] Chen F, Chen M, Kong F, et al. Species-dependent effects of crustacean plankton on a microbial community, assessed using an enclosure experiment in Lake Taihu, China [J]. Limnology and Oceanography, 2012, 57(6): 1711—1720
[10] Bai G D. Limnological survey of Wu-Li Lake during 1951 [J]. Acta Hydrobiologica Sinica, 1962, 1: 93—108 [白国栋.五里湖 1951年湖泊学调查. 水生生物学集刊, 1962, 1: 93—108]
[11] Xu R, Nauwerck A. Planktonic ciliates in north region of Taihu Lake, P. R. China [J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 1996, 35(suppl. 2): 100—104 [徐润林, Nauwerck A. 太湖北部水域的浮游纤毛虫. 中山大学学报 (自然科学版), 1996, 35(增刊2): 100—104]
[12] Cai H J. The response of ciliated protozoans to eutrophication in Meiliang Bay of Taihu Lake [J]. Journal of Lake Sciences, 1998, 10(3): 43—48 [蔡后建. 原生动物纤毛虫对太湖梅梁湾水质富营养化的响应. 湖泊科学, 1998, 10(3): 43—48]
[13] Jin X C, Tu Q Y. Survey Criteria of Lake Eutrophication [M]. Beijing: China Environmental Science Press. 1990, 302 [金相灿, 屠清瑛. 湖泊富营养化调查规范. 北京: 中国环境科学出版社. 1990, 302]
[14] Skibbe O. An improved quantitative protargol stain for ciliates and other planktonic protists [J]. Fundamental and Applied Limnology, 1994, 130(3): 339—347
[15] Foissner W, Berger H, Schaumburg J. Identification and Ecology of Limnetic Plankton Ciliates [M]. Informationsberichte des Bayer. Landesamtes für Wasserwirtschaft, Heft 3/99. 1999, 793
[16] Lynn D H. The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature (3nd ed) [M]. Springer. 2008, 605
[17] Zhang Z S, Huang X F. Method for Study on Freshwater Plankton [M]. Beijing: Science Press. 1991, 414 [章宗涉,黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社. 1991, 414]
[18] Wiackowski K, Ventelä A, Moilanen M, et al. What factors control planktonic ciliates during summer in a highly eutrophic lake [J]? Hydrobiologia, 2001, 443: 43—57
[19] Li J, Chen F, Liu Z, et al. Compositional differences among planktonic ciliate communities in four eutrophic subtropical lakes in China [J]. Limnology, 2013, 14: 105—116
[20] Havens K E, East T L. Plankton food web responses to experimental nutrient additions in a subtropical lake [J]. The Scientific World Journal, 2006, 6: 827—833
[21] Pfister G, Auer B, Arndt H. Pelagic ciliates (Protozoa, Ciliophora) of different brackish and freshwater lakes - a community analysis at the species level [J]. Limnologica, 2002, 32: 147—168
[22] Henjes J, Assmy P. Partical availability controls agglutination in pelagic tintinnids in the Southern Ocean [J]. Protist, 2007, 159: 239—250
[23] Combesa A, Dellingera M, Cadel-sixb S, et al. Ciliate Nassula sp. grazing on a microcystin-producing cyanobacterium (Planktothrix agardhii): impact on cell growth and in the microcystin fractions [J]. Aquatic Toxicology, 2013, 126: 435—441
[24] McCarthy M J, Lavrentyev P J, Yang L, et al. Nitrogen dynamics and microbial food web structure during a summer cyanobacterial bloom in a subtropical, shallow, well-mixed, eutrophic lake (Lake Taihu, China) [J]. Hydrobiologia, 2007, 581: 195—207
[25] Montagnes D J S, Allen J, Brown L, et al. Factors controlling the abundance and size distribution of the phototrophic ciliate Myrionecta rubra in open waters of the North Atlantic [J]. Journal of Eukaryotic Microbiology, 2008, 55(5): 457—465
[26] Zingel P, Haberman J. A comparison of zooplankton densities and biomass in Lakes Peipsi and Võrtsjärv (Estonia): rotifers and crustaceans versus ciliates [J]. Hydrobiology, 2008, 599: 153—159
COMMUNITY COMPOSITIONS AND HORIZONTAL DISTRIBUTION OF CILIATES IN LAKE TAIHU DURING THE CYANOBACTERIAL BLOOM IN SUMMER
LI Jing1,2, LU Wen-Xuan2, ZHANG Lei-Yan1and CHEN Fei-Zhou1
(1. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China; 2. Fisheries Research Institute of Anhui Academy of Agricultural Sciences, Hefei 230031, China)
In the present study, ciliate samples were collected from 10 sites in the estuary zones and 14 sites in the pelagic zones in the northern area of Lake Taihu in July 2009. The ciliate species and their horizontal distribution were analyzed with quantitative protargol stain (QPS) method. We also explored the relationship between the ciliate community compositions and the environmental variables. We observed 105 species that represented 60 genera, 15 orders and 3 classes. The numbers of the species in different order were counted and compared. Orders Haptorida (21 species) and Oligotrichida (20 species) ranked the first and second, followed by orders Prostomatida (12 species), Peritrichida (11 species), and Hypotrichida (11 species). These five orders were predominant in the samples, while less species belonged to the other ten orders —— Karyorelictida, Pleurostomatida, Colpodida, Nassulida, Cyrophorida, Suctoria, Hymenostomatida, Synhymeniida, Scuticociliatida, and Heterotrichida. Halteria bifurcate Tamar, H. grandinella Dujardin, Rimostrombidium brachykinetum Krainer, Tintinnopsis cylindrata Kofoid & Campbell and Cyclidium glaucoma Müller were the dominant species in both estuary (3.1%—12.3% in abundance) and pelagic (4.5%—18.1%) zones, followed by Urotricha farcta Claparède & Lachmann, Codonella cratera Leidy, R. humile Penard, Strombidium mirabile Penard, Tintinnidium pusillum Entz, Vorticella aquadulcis complex and V. campanula Ehrenberg. The abundance of ciliate was higher in the estuary zones than that in the pelagic zones, which fell in the range of 1600 to 80900 cells/L (average 31407 cells/L) and 1225 cells/L to 36000 cells/L (average 18618 cells/L), respectively. The biomass of ciliate showedthe same pattern which was 1322.6 μg/L and 543.6 μg/L on average in the estuary and pelagic zones, respectively. Neither the abundance nor the biomass of ciliates showed significant differences (P>0.05). Among the functional feeding groups, bactivorous and algivorous ones were the most abundant in all the sites. With respect to the ciliate horizontal distribution pattern, both zones had similar dominant groups while ciliates in the estuary zones showed significantly higher biodiversity in Simpson’s index, Margalef’s index, Shannon’s index and species richness (P<0.05). Our results showed that the algivorous group was barely affected by the accumulated algae, but bacterivorous assemblage was positively correlated with chlorophyll a content (P<0.001). These results implied that potentially there was a close relationship between the horizontal heterogeneity of ciliate community structures and the extent of cyanobacterial blooms.
Cyanobacterial blooms; Ciliates; Quantitative protargol stain; Lake Taihu; Horizontal distribution
Q145+.2
A
1000-3207(2014)05-0860-08
附录1 夏季北太湖检出纤毛虫种类
Appendix 1 Ciliate taxa found in summer Northern Lake Taihu
★中文名Chinese拉丁文名Latin丰度Abundance中文名Chinese拉丁文名Latin丰度Abundance动基片纲 KINETOFRAGMINOPHORA 023鼻栉毛虫 D. nasutum +核残迹目 Karyorelictida 024天鹅长吻虫 Lacrymaria olor + 001喙纤虫 Loxodes + 025尖顶瓶口虫 Lagynophrya acuminate +前口目 Prostomatida 026瓶口虫属 Lagynophrya spp. + 002浮游藤壶虫 Balanion planctonicum + 027波多长毛虫 Longitricha puytoraci + 003袋座虫属 Bursellopsis spp. + 028小中溢虫 Mesodinium acarus + 004毛板壳虫 Coleps hirtus + 029单环栉毛虫 Monodinium alveolatum + 005*毛板壳虫 C. hirtus (with symbiotic algae) + 030单环栉毛虫 Monodinium balbianii balbianii + 006裸口虫 Holophrya + 031单环栉毛虫 M. balbianii rostratum + 007前管虫属 Prorodon + 032*叶绿单环栉毛虫 *M. chlorelligerum + 008尾毛虫 Urotricha castalia + 033长颈拟多核虫 Paradileptus elephantinus + 009趣尾毛虫 U. farcta ++** 034 *Pelagodileptus trachelioides + 010双叉尾毛虫 U. furcata + 侧口目 Pleurostomatida 011敞水尾毛虫 U. pelagica + 035漫游虫一种 Litonotus sp. + 012 U. simonsbergeri + 036天鹅漫游虫 L. cygnus + 013尾毛虫属 Urotricha spp. + 肾形目 Colpodida钩刺目 Haptorida 037僧帽肾形虫 Colpoda cucullus + 014辐射射纤虫 Actinobolina radians + 038齿脊肾形虫 C. steinii + 015*小射纤虫 A. smalli + 039肾形虫属 Colpoda + 016顶口睥睨虫 Askenasia acrostomia + 040黏液篮环虫 Cyrtolophosis mucicola + 017*叶绿睥睨虫 A. chlorelligera + 041拟斜管虫属 Chilodontopsis + 018团睥睨虫 A. volvox + 篮管目 Nassulida 019 Balantidion pellucidum + 042 Obertrumia aurea + 020 Belonophrya spp. + 管口目 Cyrophorida 021*绿色缨球虫 Cyclotrichium viride + 043钩刺斜管虫 Chilodonella uncinata + 022*叶绿栉毛虫 Didinium chlorelligerum + 044斜管虫属 Chilodonella spp. +

★中文名Chinese拉丁文名Latin丰度Abundance中文名Chinese拉丁文名Latin丰度Abundance 045钟形袋齿虫 Phascolodon vorticella + 074 带核喇叭虫 S. roeseli +吸管目 Suctoria 盾纤目 Scuticociliatida 046 Gajewskajophrya + 寡毛目 Oligotrichida 047足吸管虫 Podophrya + 075杯铃壳虫 Codonella cratera +** 048华丽十字吸管虫 Staurophrya elegans + 076*双叉弹跳虫 Halteria bifurcata ++**寡膜纲 OLIGOHYMENOPHORA 077大弹跳虫 H. grandinella ++**膜口目 Hymenostomatida 078迈色虫属 Meseres + 049 *Disematostoma buetschlii + 079*绿色游跳虫 Pelagohalteria viridis + 050银白前口虫 Frontonia leucas + 080短列裂隙虫 Rimostrombidium brachykinetum ++** 051瞬目虫 Glaucoma + 081小裂隙虫 R. humile +** 052尾草履虫 Paramecium caudatum + 082透明裂隙虫 R. hyalinum +合膜目 Synhymeniida 083湖泊裂隙虫 R. lacustris + 053草履虫属 Paramecium + 084*帽形裂隙虫 R. velox + 054珍珠映毛虫 Cinetochilum margaritaceum + 085拟盗虫 Strombidinopsis chilorhax + 055膜袋虫一种 Cyclidium cf. C. elongatum + 086奇异急游虫 Strombidium mirabile +** 056银灰膜袋虫 C. glaucoma ++** 087浮游急游虫 S. pelagicum + 057膜袋虫一种 Cyclidium cf. C. versatile + 088绿急游虫 S. viride +缘毛目 Peritrichida 089短促筒壳虫 Tintinnidium ephemeridum + 058靴纤虫一种 Cothurnia(cf. C. annulata) + 090淡水筒壳虫 T. fluviatile + 059无秽累枝虫 Epistylis anastatica + 091小筒壳虫 T. pusillum +** 060平铺累枝虫 E. procumbens + 092筒壳虫一种 Tintinnidium sp. + 061累枝虫 E. pygmaeum + 093圆筒状似铃壳虫 Tintinnopsis cylindrata ++** 062后游虫属 Opisthonecta + 094似铃壳虫一种 Tintinnopsis sp. + 063水生钟虫 Vorticella aquadulcis complex +** 腹毛目 Hypotrichida 064钟形钟虫 V. campanula +**095 穆氏鬃毛虫 Chaetospira muelleri + 065*绿钟虫 V. chlorellata + 096盘状游仆虫 Euplotes patella + 066游泳钟虫 V. natans + 097锥状腹毛虫 Hypotrichidium conicum + 067春钟虫 V. vernalis + 098*叶绿尖毛虫 Oxitricha chlorelligera + 068钟虫属 Vorticella spp. + 099尖毛虫属 Oxytricha +多膜纲 POLYHYMENOPHORA 100浮游伪急游虫 Pseudostrombidium planctonticum +异毛目 Heterotrichida 101 Spiretella plancticola + 069 Bursaridium pseudobursaria + 102排毛虫属 Stichotricha + 070 Linostomella vorticella + 103片尾虫属 Urosoma + 071紫晶喇叭虫 Stentor amethystinus + 104尾枝虫属 Urostyla + 072天蓝喇叭虫 S. coeruleus + 105腹毛种1 + 073多态喇叭虫? Stentor cf. S. polymorphus +属数Genus number 60种类数Species number 105
10.7541/2014.129
2013-08-08;
2014-02-27
国家自然科学基金(31170440和41271523); 安徽省农业科学院科技创新团队建设(11C0505); 安徽省农业科学院院长青年创新基金(13B0528)资助
李静(1985—), 女, 河南商丘人; 博士; 主要研究方向为湖泊浮游动物生态学。E-mail: LJLJ278@126.com
陈非洲, E-mail: feizhch@niglas.ac.cn
(续附表)
