亚布力山涧溪流秋季水生昆虫群落特征及其与环境因子的关系1)
2014-03-05阚春梅于洪贤
阚春梅 于洪贤
(东北林业大学,哈尔滨,150040)
水生昆虫作为大型底栖无脊椎动物的重要组成部分,具有生活周期长,生活场所比较固定,以及个体大,易于采集和识别的特征[1-3]。在水生生态系统物质循环和能量流动中具有不可替代的作用[4-5]。由于各类底栖动物对环境条件的适应性以及对污染程度的耐受力和敏感程度不同,可利用底栖动物的种群结构、优势种类、数量等参数来反应环境质量状况,国内外对此已有较多的研究[6]。例如在美国应用大型无脊椎动物科级和属级指示者的耐污值对溪流进行评价[7],在德国利用底栖无脊椎动物发展多元评价系统对溪流进行评价[8]。国内在此方面的研究多集中在南方,如利用水生昆虫评价南昆山溪流的水质[9]、研究西南山地源头溪流附石性水生昆虫群落特征及多样性[10]。北方对溪流水生昆虫群落结构研究较少,仅见李金国等对[11]凉水、帽儿山低级溪流水生昆虫群落结构及李亚俊等[12]利用水生昆虫对水质的敏感性对阿什河上游水质进行生物学评价的相关报道。本研究以北方寒温带亚布力山涧溪流秋季水生昆虫为研究对象,分析溪流中水生昆虫的空间分布及其与环境因素之间的关系,为进一步研究水生昆虫在水生态系统中的作用提供基础数据和理论支持。
1 研究区域概况
亚布力位于黑龙江省尚志市,地处长白山系余脉张广才岭西麓大锅盔山脚下,其旅游资源得天独厚。世界公认的冰雪、森林、海洋三大旅游资源,亚布力占有冰雪、森林两项,是世界十大滑雪场之一。目前,已成为中外驰名的滑雪旅游胜地和国家4A级滑雪旅游度假区。其气候特点属于温带季风气候,年平均气温6 ℃。冬季山上积雪厚达1 m 左右,春季温度升高,冰雪融化汇成许多溪流,溪流是生态系统的重要组成部分。本研究选取锅盔山一条天然溪流为研究对象,海拔为526 m(图1)。
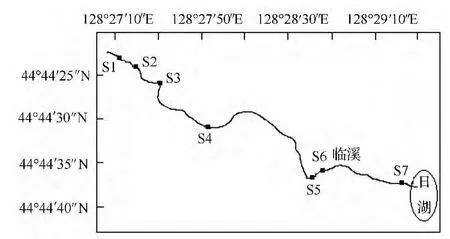
图1 亚布力山涧溪流采样点示意图
2 研究方法
2.1 样品采集与处理
2012年9月对溪流中各个采样点进行水生昆虫采集。样品采集用索伯网(0.3 m×0.3 m)和D 型抄网(直径宽0.3 m),D 型抄网采集时,采集长度为3 m,采样面积约0.9 m2。采样时用脚搅动网前底质,并用刷子刷下粘附在石头上的底栖动物,枯枝落叶及水草也装入样品瓶中,用乙醇固定,带回实验室。在瓷盘中挑出水生昆虫,对水生昆虫样本进行整理归类。物种鉴定根据Morse[13]、Merritt[14]和周长发[15]等文献,大多样本鉴定至种,少数鉴定至科、属、区分到种,并将标本保存于75%的酒精中。
2.2 水体理化指标的测定
采集标本的同时,利用YSI-6600 多功能水质监测仪现场测定溪流的pH、水温、叶绿素a、溶解氧、电导率、氨氮、硝态氮等水体理化指标,并用米尺测量水深和水面宽,再利用LS25-1 型旋桨式流速仪测定流速。用棕色瓶采集500 mL 水样带回实验室进行总氮、总磷测定。总氮采用紫外分光光度法,总磷采用钼蓝法(表1)。

表1 溪流水体理化指标
2.3 数据处理与分析
多维标度分析(MDS)利用SPSS13.0 进行数据统计分析,以Bray-Curtis[16]相似性系数SB为基础进行计算,SB计算公式:
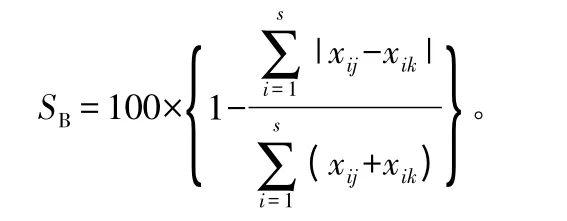
式中:xij是第i 物种在第j 样方的数量;xik是第i 物种在第k 样方中数量。
物种多样性及生物指数采用物种优势度指数(y)[17]、Margalef 种类丰富度指数(d)[18]、Shannon-Wiener 多 样 性 指 数(H')[19]、Pielou 均 匀 度 指 数(J)[20]进行分析。利用典范对应分析(CCA)对水生昆虫与环境因子的相互关系进行研究,在CANOCO4.5 软件上运行。
3 结果与分析
3.1 水生昆虫的种类组成
共采集水生昆虫52 种,录属于6 目23 科(表2)。从科级别来看,由高到低依次为:毛翅目(9种)、双翅目(6 种)、蜉蝣目(4 种)、襀翅目(2 种)、广翅目(1 种)、鞘翅目(1 种)。从物种数来看,由高到低依次为:双翅目、毛翅目、蜉蝣目、襀翅目、广翅目、鞘翅目。双翅目种类最多,为27 种,占总数的51.92%;其次是毛翅目12 种,占23.08%;蜉蝣目6种,占11.54%;襀翅目5 种,占9.62%;广翅目和鞘翅目各1 种,占1.92%。由表2可知,溪流水生昆虫主要由双翅目、毛翅目和蜉蝣目三大类群组成,共占总个体数的87.26%。
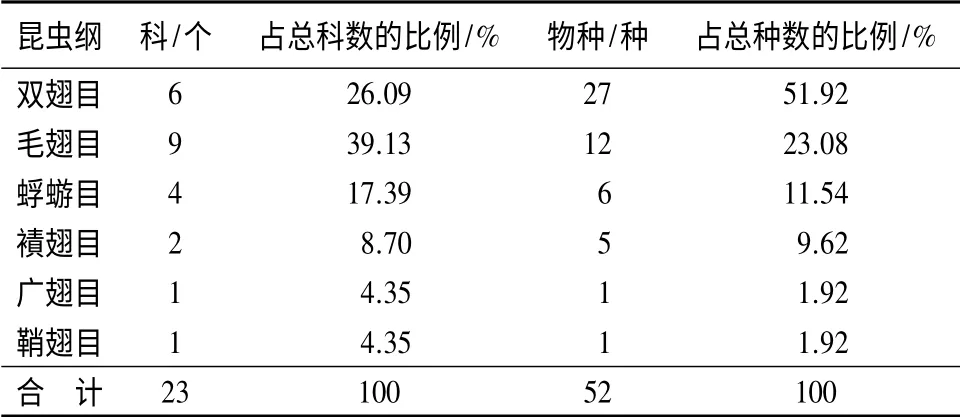
表2 亚布力山涧溪流水生昆虫科及种类的比例
3.2 水生昆虫的密度、生物量及优势度
亚布力溪流中水生昆虫的平均密度和生物量分别为2 249 个/m2和14.83 g/m2。各个采样点密度由大到小为S4、S7、S5、S6、S2、S1、S3,其中S4 密度最大,为3 500 个/m2;S1、S2、S3 都较小。而各个采样点的生物量差异较大,其中S4 最大,为30.17 g/m2;S3 最小,为5.80 g/m2(表3)。S1、S2、S3 海拔较高,水温较低,可能是导致水生昆虫密度和生物量降低的主要原因。根据优势度指数y>0.02,可知溪流水生昆虫优势种为巴瓦特维摇蚊(Tvetenia bavarica)、流环足摇蚊(Rheocricotopus sp.)以及毛蠓(Psychoda sp.)、小蜉(Ephemerella sp.)、四节蜉(Baetis sp.)和Mystrophora inops。
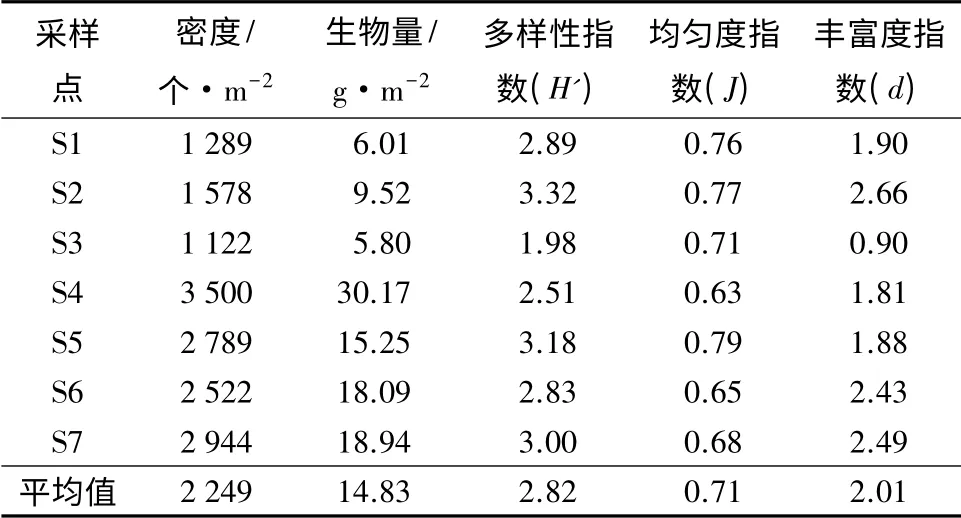
表3 亚布力山涧溪流水生昆虫的群落特征
3.3 溪流水生昆虫的多样性
由表3可知,水生昆虫的多样性指数变化范围为1.98 ~3.32,其中S3 最小。而Margalef 丰富度指数为0.90 ~2.49,其中S3 采样点水生昆虫丰富度指数最低。S3 采样点多样性指数和丰富度指数都较小的原因,可能是由于溪流宽度较窄,底质主要以细砂和基石为主,腐蚀质较少,生境单一,不利于水生昆虫生存。因此造成多样性指数和丰富度指数都最低。
3.4 溪流水生昆虫的群落结构
采用多维排序尺度(MDS)分析亚布力山涧溪流中水生昆虫的群落特征,得到二维标度序列分析图(图2)。从图2可以看出,秋季亚布力山涧溪流水生昆虫的分布主要分为4 个群落区域。S1 采样点一个聚类组,S7 采样点一个聚类组,S2 和S3 采样点一个聚类组,S4、S5 和S6 采样点一个聚类组。秋季的Stress 值为0.050 9(Stress<0.1),说明得到的MDS 图是一个较好的排序,说明该数据利用MDS图能够很好地解释溪流各采样点水生昆虫群落的相似性关系,同时也能显示各采样点水生昆虫群落组成存在的差异。

图2 亚布力山涧各采样点水生昆虫群落的MDS 分析结果
3.5 水生昆虫分布与环境因子的关系
不同种类的底栖动物对外界环境的适应性都存在差异[21]。本研究对出现频率在15%以上的26 个物种与12 个环境因子指标进行典范对应分析,结果见图3。
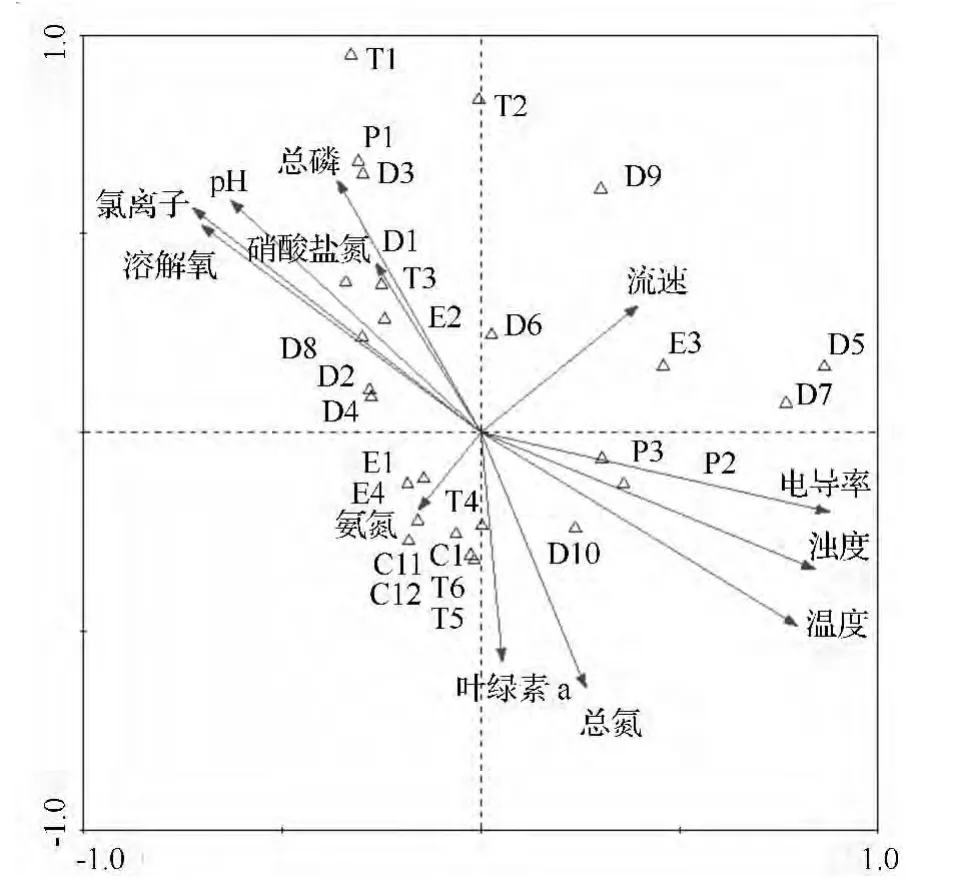
图3 出现频率>15%的水生昆虫与环境因子间的典范对应分析(CCA)排序图
典范对应分析获得前2 个排序轴的特征值为0.654 和0.391,占总特征值1.460 的44.8%(表4)。说明参与分析的12 个环境因子对出现频率在15%以上的水生昆虫分布情况的解释程度为44.8%。前2个排序轴水生昆虫与环境因子的相关系数均为1.000,说明排序结果非常可信。对于水生昆虫来说,第一、第二排序轴之间的相关系数为0,说明这2 个排序轴相互垂直;对于环境因子来说,前2 个排序轴的相关性也为0,表明这2 个排序轴也是垂直的,说明排序图能够反映大型底栖动物种类与环境因素间的关系[22]。从图3可以看出:温度、电导率和浊度与第一轴正相关,溶解氧、pH 和氯离子与第一轴负相关;溶解氧、pH、氯离子和总磷与第二轴正相关,叶绿素a 和总氮与第二轴负相关。在CCA 排序图中,环境因子用带有箭头的线段(矢量)表示,连线的长短表示物种的分布与该环境因子关系的大小[23]。从向量长短揭示出秋季主要有温度、溶解氧、氯离子、电导率和浊度共5 项环境因子与水生昆虫的水平分布密切关系。因此可以得出,水体的温度、溶解氧、氯离子、电导率和浊度是影响秋季溪流水生昆虫分布的主要环境因子。
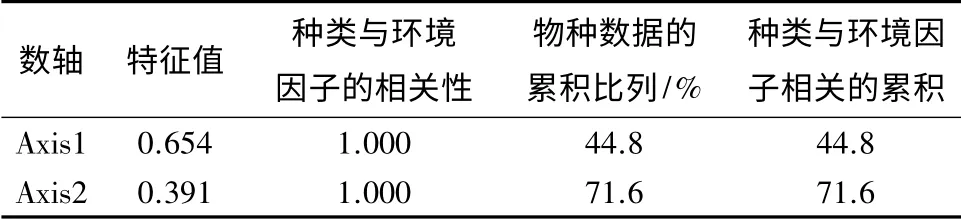
表4 秋季CCA 分析前两个排序轴特征值及种类与环境因子排序轴的相关系数
4 结论与讨论
从调查结果来看,共采集水生昆虫52 种,录属于6 目23 科。各个采样点的水生昆虫多样性都较高,说明亚布力山涧溪流生态环境状态良好,水环境质量优良。其中S3 采样点最少,只有某些毛翅目昆虫以筑巢的形式附着在基石表面,这可能与溪流宽度较窄,底质主要以细砂和基石为主,腐蚀质较少,生境较单一,不利于其他水生昆虫生存有关。
通常毛翅目、襀翅目和蜉蝣目是喜急流和洁净水体并对水质最敏感的水生生物类群[24]。山地溪流水流湍急,水中溶解氧充足,水质洁净,底质多为圆石、漂砾[25]。因此在山地溪流的水生昆虫群落中,EPT 昆虫往往成为优势类群[9,11,25-26]。而本研究中双翅目、毛翅目、蜉蝣目为水生昆虫的三大优势类群,占水生昆虫总数量的86.5%。这与王强等[10]研究西南山地源头溪流附石性水生昆虫群落特征及多样性的研究结果相似。
河流连续理论认为,由源头溪流至下游的各级河流,在地理空间、生物学过程及其物理环境上形成一个连续系统[27-28]。按照该理论,连续变化的河流其生物群落特征是可以预测的[10]。随着海拔的升高,水生昆虫群落多样性常常会逐步降低[29-32]。而在不同的水域中生活的水生昆虫的多样性,主要决定于水的理化条件,如:温度、矿物质、有机质和pH值等[12]。在本次调查中S5、S4、S3 采样点水生昆虫群落多样性随着海拔升高逐渐降低,而S1 和S2 采样点海拔最高,其多样性指数也较高,可能是河岸相对平缓,水面较宽阔,还受水体的理化因素影响,如水中溶解氧较高,浊度低,有利于各种水生昆虫生存。从而导致S1 和S2 水生昆虫多样性较高。
典范对应分析表明,水体的温度、溶解氧、氯离子、电导率和浊度对亚布力山涧溪流中水生昆虫分布的作用比较大。而其他环境因子,如流速和硝酸盐氮等都会对水生昆虫多样性产生重要影响。因此,控制水体中的温度、氯离子、电导率和浊度以及增加溶解氧是保护亚布力山涧溪流水生昆虫多样性的可行性手段之一。
[1] 王备新,杨莲芳.用河流生物指数评价秦淮河上游水质的研究[J].生态学报,2003,23(10):2082-2091.
[2] 戴友芝,唐受印,张建波.洞庭湖底栖动物种类分布及水质生物学评价[J].生态学报,2000,20(2):277-282.
[3] 王备新,杨莲芳.大型底栖无脊椎动物水质快速生物评价的研究进展[J].南京农业大学学报,2001,24(4):107-111.
[4] 渠晓东,曹明,邵美玲,等.雅砻江(锦屏段)及其主要支流的大型底栖动物[J].应用生态学报,2007,18(1):158-162.
[5] 江晶,温芳妮,顾鹏,等.湖北清江流域胡家溪大型底栖动物群落结构及水质评价[J].湖泊科学,2009,21(4):547-555.
[6] 高峰,尹洪斌,胡维平,等.巢湖流域春季大型底栖动物群落生态特征及与环境因子关系[J].应用生态学报,2010,21(8):2132-2139.
[7] Carlisle D M,Meador M R,Moulton S R Ⅱ,et al. Estimation and application of indicator values for common macroinvertebrate genera and families of the United States[J]. Ecological Indicators,2007,7(1):22-33.
[8] Böhmer J,Rawer-Jost C,Zenker A,et al. Assessing streams in Germany with benthic invertebrates:development of a multimetric invertebrate based assessment system[J]. Limnologica-Ecology and Management of Inland Waters,2004,34(4):416-432.
[9] 童晓立,胡慧建,陈思源.利用水生昆虫评价南昆山溪流的水质[J].华南农业大学学报,1995,16(3):6-10.
[10] 王强,袁兴中,刘红.西南山地源头溪流附石性水生昆虫群落特征及多样性:以重庆鱼肚河为例[J].水生生物学报,2011,35(5):887-892.
[11] 李金国,王庆成,严善春,等.凉水、帽儿山低级溪流中水生昆虫的群落特征及水质生物评价[J].生态学报,2007,27(12):5008-5018.
[12] 李亚俊,吴梦婷,张宇,等.利用水生昆虫对阿什河上游秋季水质进行生物学评价[J].东北林业大学学报,2013,41(3):135-139.
[13] Morse J C,Yang Liangfang,Tian Lixin. Aquatic insects of China useful for monitoring water quality[M]. Nanjing:HeHai University Press,1994.
[14] Merritt R W,Cummins K W. An introduction to the aquatic insects of North American[M]. Dubuque:Kendall/Hunt Publishing,1996:521-632.
[15] 周长发.中国大陆蜉蝣目分类研究[D].天津:南开大学,2002:44-121.
[16] Bray J R,Curtis J T. An ordination of the upland forest communities of Southern Wisconsin[J]. Ecological Monographs,1957,27:325-349.
[17] 徐兆礼,王云龙,陈亚瞿,等.长江口最大浑浊带区浮游动物的生态研究[J].中国水产科学,1995,2(1):39-48.
[18] Margalef R. Information theory in ecology[J]. General Systems,1958,3:36-71.
[19] Shannon C E. A mathematical theory of communication[J]. The Bell System Technical Journal,1948,27:379-423.
[20] 吕光俊,熊邦喜,刘敏,等.不同营养类型水库大型底栖动物的群落结构特征及其水质评价[J].生态学报,2009,29(10):5339-5349.
[21] 蒋万祥,贾兴焕,周淑婵,等.香溪河大型底栖动物群落结构季节动态[J].应用生态学报,2009,20(4):923-928.
[22] Ter Braak C J F. Canonical correspondence analysis:a new eigenvector technique for multivariate direct gradient analysis[J]. Ecology,1986,67(5):1167-1179.
[23] 曹同,郭水良,高谦.应用排序分析藓类植物分类群分布与气候因素的关系[J].应用生态学报,2000,11(5):680-686.
[24] 李强,杨莲芳,吴璟,等.西苕溪EPT 昆虫群落分布与环境因子的典范对应分析[J].生态学报,2006,26(11):3817-3825.
[25] 于力,暴学祥,云宝琛.长白山水生昆虫的研究[J].水生生物学报,1997,21(1):31-39.
[26] 渠晓东.香溪河大型底栖动物时空动态、生物完整性及小水电站的影响研究[D].武汉:中国科学院水生生物研究所,2006.
[27] Vannote R L,Minshall G W,Cummins K W,et al. The river continuum concept[J]. Canadian Journal of Fisheries and Aquatic Sciences,1980,37:130-137.
[28] 蔡庆华,唐涛,刘建康.河流生态学研究中的几个热点问题[J].应用生态学报,2003,14(9):1573-1577.
[29] Houghton D C. Biodiversity of Minnesota caddisflies (Insecta:Trichoptera):delineation and characterization of regions[J]. Environmental Monitoring and Assessment,2004,95(1/3):153-181.
[30] Heino J,Parviainen J,Paavola R,et al. Characterizing macroinvertebrate assemblage structure in relation to stream size and tributary position[J]. Hydrobiologia,2005,539(1):121-130.
[31] Paller M H,Specht W L,Dyer S A. Effects of stream size on taxa richness and other commonly used benthic bioassessment metrics[J]. Hydrobiologia,2006,568(1):309-316.
[32] Deván P,Mucinaz L. Structure,zonation,and species diversity of the mayfly communities of the Belá River basin Slovakia[J].Hydrobiologia,1986,135(1/2):155-165.
