草鱼呼肠孤病毒抗原捕获ELISA检测方法的建立
2014-02-24景宏丽林祥梅张利峰吴绍强
景宏丽,曹 欢,张 旻,王 娜,林祥梅,张利峰,吴绍强
(1.中国检验检疫科学研究院动物检疫研究所,北京 100029;
2.北京市水产技术推广站,北京 100021;3.北京出入境检验检疫局检验检疫技术中心,北京 101113)
草鱼呼肠孤病毒抗原捕获ELISA检测方法的建立
景宏丽1,曹 欢2,张 旻1,王 娜1,林祥梅1,张利峰3,吴绍强1
(1.中国检验检疫科学研究院动物检疫研究所,北京 100029;
2.北京市水产技术推广站,北京 100021;3.北京出入境检验检疫局检验检疫技术中心,北京 101113)
研究以纯化的羊抗草鱼呼肠孤病毒多克隆抗体作为捕获抗体,抗草鱼呼肠孤病毒的单克隆抗体为检测抗体,建立草鱼呼肠孤病毒抗原捕获ELISA检测方法,其最佳反应条件是:羊多克隆抗体包埋浓度为2.5μg/ mL,抗草鱼呼肠孤病毒单克隆抗体工作浓度为1∶400稀释,以3%牛血清白蛋白作为封闭液。该方法的检测限为5×105pfu/mL,且能够特异性检出发病草鱼内脏组织中的草鱼呼肠孤病毒。特异性试验结果表明该方法具有较高的特异性,与病毒性出血性败血症病毒、传染性造血器官坏死病毒和鲤春血症病毒等水产动物病毒株均不反应。该方法还具有较好的稳定性。
草鱼呼肠孤病毒;抗原捕获ELISA;单克隆抗体
草鱼呼肠孤病毒(grass carp reovirus,GCRV)是中国分离到的第一株鱼类病毒,是草鱼出血病的主要病原。草鱼出血病发病季节长,流行范围广,防治方法有限。据不完全统计,每年由于草鱼出血病导致的经济损失达21.36亿元[1-2]。
该病毒为20面体的球形颗粒,直径为70-80nm,具双层衣壳,无囊膜。病毒基因组为双股RNA,由11条片段组成[3]。据数据,目前在患有草鱼出血病病鱼中已经分离到了多种草鱼呼肠孤病毒,包括GCRV873,GCRV875,GCRV HZ08,
GCRVD108和AGCRV等。有些学者认为这些草鱼呼肠孤病毒株在毒力水平、细胞培养特性和免疫抗原性都具有较大的差异性[4-8]。
鱼类的病毒病通常是以“预防”为主,只有建立快速、敏感、特异的草鱼出血病诊断技术并应用到实际防治工作中去才能真正做到这一点。目前基于草鱼呼肠孤病毒的多样性,只能先建立一种研究最为成熟的草鱼呼肠孤病毒的检测方法,然后推广,并比较病毒株之间差异,以期能够建立一种能够真正预防草鱼出血病的检测方法。
GCRV873株,曾是我国南方草鱼养殖地区草鱼出血病的主要病原,也是我国研究最早的一株病毒株,检测方法也较为全面,主要包括电镜观察法、各种PCR法[9-10]、细胞分离法和逆转录环介导等温扩增(RT-LAMP)法[11-12]。这些方法都各有一定的不足,如检测时间长、操作复杂和依赖贵重仪器等。本研究旨在建立一种抗原捕获ELISA检测法,并初步应用该ELISA方法检测草鱼出血病病患鱼的组织。
1 材料与方法
1.1 病毒株和细胞系
草鱼呼肠孤病毒湖南株(GCRV873)由武汉病毒所提供;对照毒株:鲤春血症病毒(spring viraemia of carp virus,SVCV)和传染性造血器官坏死病毒(infectious haematopoietic necrosis virus,IHNV)由英国Weymouth的OIE参考实验室惠赠;传染性胰脏坏死病毒(infectious pancreatic necrosis virus,IPNV)由丹麦Ahus的OIE参考实验室确认;流行性造血器官坏死病毒( epizootic haematopoietic necrosis virus,EHNV)由澳大利亚OIE参考实验室惠赠;大马哈鱼病毒(Chum salmon reovirus,CSV)和病毒性出血性败血症病毒(viral haemorrhagic septicemia virus,VHSV)由本实验室保存;试验中所用细胞系有蓝鳃太阳鱼细胞(BF-2)、草鱼卵巢细胞(CO)、草鱼肾细胞(CIK)、鲑囊胚细胞(CHSE)、胖头鱥肌肉细胞(FHM)、鲤上皮瘤细胞(EPC)、虹鳟肝细胞(R1)、虹鳟性腺细胞(RTG-2),均用含10%胎牛血清的199培养液培养。
1.2 抗体和其他试剂
实验中应用的抗草鱼呼肠孤病毒的单克隆抗体由本实验室制备[13];硫酸盐(TMB),购自Sigma公司;辣根过氧化物酶(HRP)标记羊抗小鼠免疫球蛋白(Ig),购自SIGMA公司;其他试剂为国产化学试剂。
1.3 草鱼呼肠孤病毒(GCRV)的制备
将贴壁生长状况良好的鲤鱼上皮瘤细胞(EPC)于75cm3细胞培养瓶中传代(每瓶加入约12mL培养液),置25℃生化培养箱培养。取生长24h以内且已经长满单层的细胞接种GCRV病毒悬液,接种量为0.2mL/瓶。然后,置25℃生化培养箱培养。每天观察细胞,直至出现100%细胞病变(CPE),即细胞结构破坏,完全脱落,且悬浮病毒合胞体。收集病毒悬液-80℃保存且标定收集到病毒悬液的TCID50/0.1mL。
1.4 羊抗草鱼呼肠孤病毒多克隆抗体的制备和纯化
上述收集到的病毒悬液,以8000r/min离心35min,收集上清;24000r/min离心2h,收集沉淀作为为抗原,分别多点注射免疫山羊,每周一次,共4次;取血清,分装,-80℃保存。
采用饱和硫酸铵法纯化羊抗GCRV多克隆抗体:羊血清与0.01mol/L的PBS等体积混合4℃不停搅拌,逐渐加入饱和硫酸铵终浓度为35%。4℃下,搅拌1h。9000r/min离心20min。收集沉淀,用0.01mol/L的PBS重悬。4℃不停搅拌,逐渐加入饱和硫酸铵终浓度为45%。4℃下,搅拌1h。9000r/min离心20min。收集沉淀,用0.01mol/L的PBS重悬。把溶液装入到透析袋中,在0.01mol/ L的PBS中透析24h。离心,分装,即得到纯化的山羊IgG。
1.5 ELISA方法的建立和优化
1.5.1 方法基本程序[10]
(1)包埋纯化的羊多克隆抗体,每孔100μL,4℃过夜;
(2)洗涤:用PBST洗涤ELISA板3次,每次2min,甩干;
(3)封闭:加3﹪BSA,置37℃,60min;
(4)加样:待测样品、阳性样品和阴性样品,每孔100μL,37℃反应90min;
(5)洗涤:用PBST洗涤3次,每次2min,甩干;
(6)一抗:每孔加抗GCRV的单克隆抗体100μL,37℃反应90min;
(7)洗涤:用PBST洗涤3次,每次2min,甩干;
(8)去非特异性过氧化物酶:加0.1%H2O2,每孔300μL,37℃反应15min;
(9)洗涤:用PBST洗涤3次,每次2min,甩干;
(10)二抗:每孔加100μL辣根过氧化物酶(HRP)标记小鼠抗山羊IgG作二抗,37℃反应90min;
(11)洗涤:用PBST洗涤3次,每次2min,甩干;
(12)显色:每孔加100μL显色剂,室温暗处避光反应5min,阳性显色,阴性未显示时终止;
(13)终止:每孔加150μL2M H2SO4终止反应;
(14)酶标仪测定:测定波长为450 nm时各孔的吸光度(A)值,计算阳性血清与阴性血清光吸收值之比(P/N),当P/N≥2.1时判定为阳性。
1.5.2 抗体最佳浓度确定
包埋纯化的羊抗IgG(10μg/mL;5μg/ mL;2.5μg/mL;1.25μg/mL),鼠抗GCRV单克隆抗体(1∶400;1∶800;1∶1600),病毒抗原(V0:5×106pfu/mL;V1:5×105pfu/mL;V2:5×104pfu/mL),棋盘滴定方法进行上述ELISA实验。
1.5.3 特异性试验
其他水生动物病毒包括IHNV、SVCV、VHSV、IPNV和CSV(浓度均为5×106pfu/mL)及其增殖用水生动物细胞系作为待检测样品,进行ELISA实验。
1.5.4 敏感性试验
检测抗原GCRV浓度均为5×106pfu/mL,10倍系列稀释,进行ELISA检测。
1.5.5 稳定性试验
所有ELISA中的试剂,在10℃和37℃分别放置3d、6d和9d,进行ELISA检测。
1.6 ELISA方法的应用
在某单位获得患有草鱼出血病的鱼类内脏组织样品3份,分别进行ELISA和PCR检测。PCR方法依照检测草鱼出血病的行业标准(SN/T3584-2013)进行。
2 结果
2.1 抗体最佳浓度确定
包埋纯化羊抗和鼠单克隆抗体最适浓度的确定,根据P/N值最大,且保证病毒检测的敏感度最高,确定包埋羊多克隆抗体浓度为2.5μg/ mL,鼠单克隆抗体浓度为1∶400(表1)。
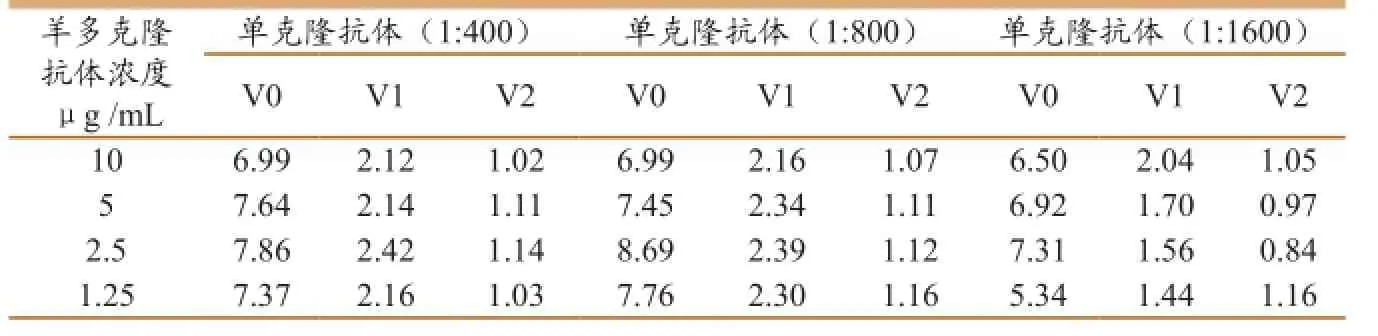
表1抗体最适浓度的确定(P/N值)
2.2 特异性
在特异性实验中,ELISA方法特异性较高,与多株病毒株和细胞株发生反应时,只与GCRV反应,即P/N值为5.18;而与其他病毒株和细胞株均不发生反应(表2)。
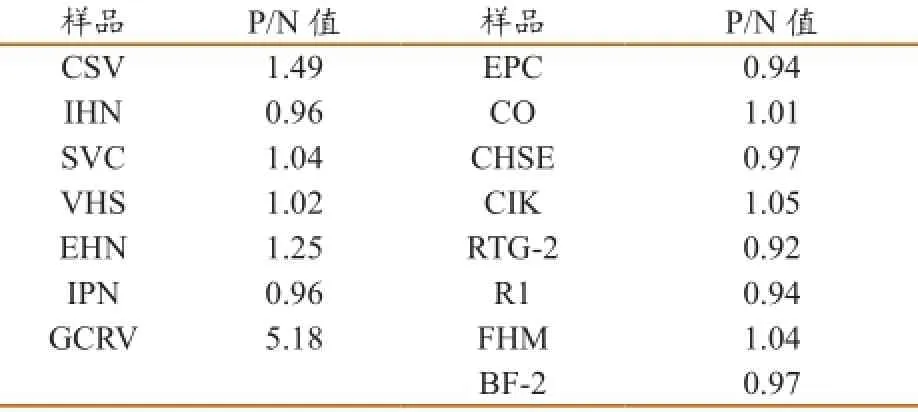
表2特异性实验结果(P/N值)
2.3 敏感性
建立的ELISA方法最低能够检测到5×105pfu/ mL(表3)。

表3敏感性实验结果(P/N值)
2.4 稳定性
在稳定性试验中,ELISA方法中所有试剂在
10℃和37℃稳定性比较好,表4所示,阳性样品和阴性样品的相关数据,其他试剂的数据未列出。在参与国家认监委组织的2013年能力验证项目,19个实验室均能够达到满意标准,也进一步证明了ELISA检测方法的具有较好的稳定性。
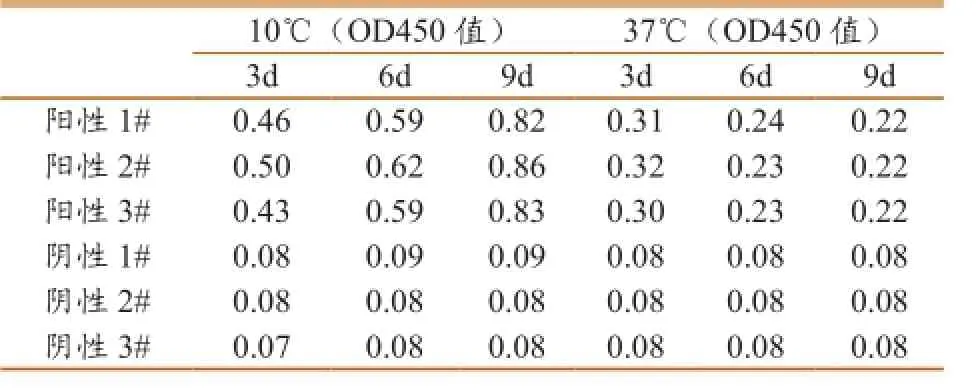
表4稳定性试验结果
2.5 ELISA方法的应用
在某单位获得草鱼出血病样品(分别命名为1、2、3),ELISA结果显示3个样品均为阳性(表5),而根据GCRV行业标准,进行PCR检测,结果显示只有2和3为阳性样品(图1)。

表5 ELISA检测样品结果(P/N值)
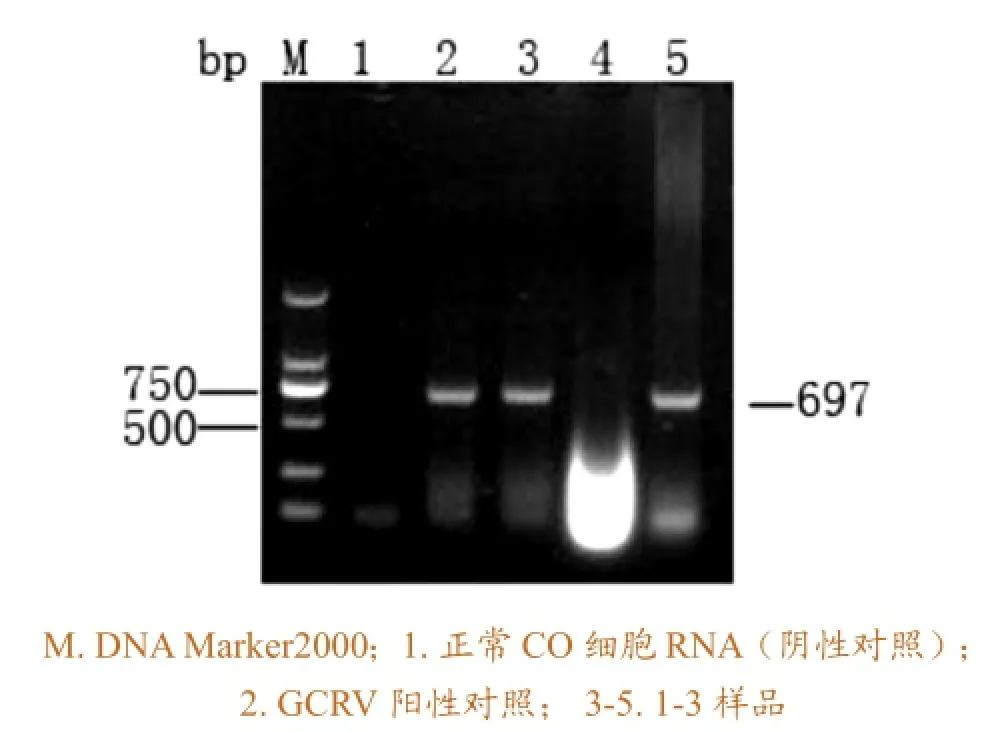
图1 RT-PCR检测结果
3 讨论
本次试验所用的抗GCRV单克隆抗体经用Western-blotting和间接ELISA分析其特性,证明单克隆抗体具有针对GCRV的特异性[13]。试验中采用的两种特异性抗体,形成了“抗GCRV羊多克隆抗体-GCRV抗原-抗GCRV单克隆抗体”的夹心结构,提高了抗原抗体反应特异性,减少了假阳性结果。另外采用方阵滴定法对抗体浓度进行了相关优化,提高了反应灵敏度。
抗原捕获ELISA检测方法中所有试剂在10℃和37℃在9天内稳定性都比较好,但是从结果上可以明显看出放在10℃时的稳定性更好。在后期研究中,本实验室还进行在10℃放置30天的稳定性试验,结果显示仍有较强的阳性反应,表明建立的抗原捕获ELISA方法稳定性较好,更长时间的稳定性研究还在进行中。
试验中应用的抗原捕获ELISA方法对具有明显症状的草鱼,3份肝、脾和肾等内脏混合组织进行检测,结果表明3份样品含有GCRV病毒。在进行ELISA方法的同时本实验室根据检测草鱼出血病的行业标准(SN/T3584-2013)进行了PCR检测,结果只有2份样品为阳性。抗原捕获ELISA方法结果与PCR方法结果有一定的差异。这一差异尽管不能说明在对具有发病症状的草鱼内脏组织进行检测抗原捕获ELISA方法的准确性优于PCR检测方法,但是至少能够说明建立的抗原捕获ELISA方法能够特异性的检测具有明显发病症状草鱼的中携带的GCRV。由于草鱼出血病的爆发是有时间性的,采集发病鱼类的样品就有一定的限制性,因此建立的抗原捕获ELISA方法准确性有待进一步研究,而抗原捕获ELISA方法与PCR方法的比较也需要进一步研究。
[1]刘宝芹,曾伟伟,王庆,等.草鱼呼肠孤病毒株-检测方法的建立及应用[J].中国水产科学,2012,19(2):329-335.
[2]戈贤平.我国大宗淡水鱼养殖现状及产业技术体系建设[J].中国水产,2010(5):5-9.
[3]Rangel AAC,Rockemann DD,Hetrick FM,et al. Identification of grass carp haemorrhage virus as a new genogroup of aquareovirus[J].J Gen Virol,1999,80(9):2399-2402.
[4]曾伟伟,王庆,王英英,等.草鱼呼肠孤病毒株蛋白单克隆抗体的制备及鉴定[J].水产学报,2013,37(3):450-456.
[5]迟妍妍,田园园,叶星,等.南方养殖草鱼呼肠孤病毒的分子特性比较及双重检测方法的建立[J].病毒学报,2011,27(4):358-365.
[6]Ye X,Tian Y Y,et al. Complete genomic sequence of a reovirus isolated from grass carp in China[J].Virus Res,2012,163(1):275-283.
[7]Zeng WW,Wang Q,et al. A one-step molecular biology method for simple and rapid detection of grass carp Ctenopharyngodon idella reovirus(GCRV)HZ08 strain[J].J Fish Biol,2013,82(5):1545-1555
[8]Yan XY,Wang Y,et al. Phylogenetic analysis of newly isolated grass carp reovirus doi:10.1186/2193-1801-3-190.
[9]Zhang L,Luo Q,et al. An improved RT-PCR assay for rapid and sensitive detection of grass carp reovirus[J].J Virol Methods,2010,169(1):28-33.
[10]Guo Z X,Weng S P,et al. Development of an RT-PCR detection method for mud crab reovirus[J].J Virol Methods,2008 ,151(2):237-241.
[11]Zhang QL,Yan Y,et al. Development of a reverse transcription loop-mediated isothermal amplifcation assay for rapid detection of grass carp reovirus[J].J Virol Methods,2013,187(2):384-389.
[12]张金凤,曾令兵,张辉,等.草鱼呼肠孤病毒逆转录环介导等温扩增-检测方法的建立[J].中国水产科学,2013,120(1):129-136.
[13]Hongli J,Lifeng Z,et al. Detection of grass carp reovirus(GCRV)with monoclonal antibodies[[J].Arch Virol,2014,159(4):649-655.
Establishment of Antigen-Capture ELISA for Detection of Grass Carp Reovirus
Jing Hongli1,Cao Huan2,Zhang Min1,Wang Na1,Lin Xiangmei1,Zhang Lifeng3,Wu Shaoqiang1*
(1. Institute of Animal Quarantine,Chinese Academy of Inspection and Quarantine,Beijing 10002;2. Beijing Aquiculture Technology Extension Station,Beijing 100021;3. Research Centre of Aquatic Animal Diseases,Beijing Exit-Entry Inspection and Quarantine Bureau,Beijing 101113)
An antigen-capture enzyme-linked immunoassay(ELISA)was developed for the detection of grass carp reovirus(GCRV)with purifed polyclonal antiserum against GCRV as capture antibody and monoclonal antibodies against GCRV as detection antibody. The 96-well enzyme immunoassay(EIA)plates were coated with 100μL purifed polyclonal antiserum at concentration of 2.5μg /mL and blocked with 3% bovine serum albumin(BSA). The working concentration of monoclonal antibody was diluted to 1:400 with distilled water. GCRV was detected in culture supernatants(5×105pfu/mL)and in the extracts of kidney and spleen of infected grass carp. The assay was highly specifc:viral hemorrhagic septicemia virus,infectious haematopoietic necrosis virus,spring viraemia of carp virus,epizootic haematopoietic necrosis virus,chinook salmon reovirus could not be detected by the developed ELISA and the method was also of good stability.
grass carp reovirus;antigen-capture ELISA;monoclonal antibody
S941.41
:A
:1005-944X(2014)12-0078-05
“十二五”国家科技支撑计划项目《重大外来与新发水生动物疫病识别与监测技术研究及示范》(2013BAD12B02)
吴绍强
