青少年暴力行为与多巴胺β羟化酶基因多态性的关联分析
2014-02-03邹志礼蒙华庆马中锐张洪银
邹志礼,蒙华庆△,胡 华▲,周 波,马中锐,张洪银
(1.重庆医科大学附属第一医院心理卫生中心 400016;2.四川省人民医院心身医学中心,成都 610072)
近年来,青少年暴力攻击行为已成为世界性的公共卫生问题,它不仅影响青少年本身的学习和社会功能,而且还影响家庭及社会的和谐稳定。引起学者们对此的广泛关注,目前,对其进行了多方面的研究。在遗传的研究中涉及到一系列的基因多态性,如:5-HT受体、5-HT转运体(5-HTT)、多巴胺受体、儿茶酚胺氧化甲基转移酶(COMT)、单胺氧化酶A(MOMA)等相关基因[1-6],但结果却不尽相同。多巴胺β羟化酶(dopamine-β-hydroxylase,DβH)是DA合成去甲肾上腺素的限速酶,其活性对儿茶酚胺的代谢产生重要的影响。先前学者发现DβH基因5′侧翼区的-1021C→T多态性与血浆中的DβH活性相关[7]。同时有学者报道有行为问题的儿童血清中DβH水平较对照组低[8]。由此可以推测DβH基因结构或表达的差异可能与暴力行为的易感性有关。因此,本研究选取DβH基因与青少年暴力行为进行了关联分析,现报道如下。
1 资料与方法
1.1一般资料 根据官方记录将新入的男性青少年服教人员分为暴力罪犯和非暴力罪犯。再根据修订版外显攻击行为量表[9](MOAS)筛查,将MOAS加权分大于或等于4分,体力攻击大于或等于1分定为有暴力行为,MOAS加权分小于4分,体力攻击为0分定为无暴力行为。入组标准:所有受试者均为男性,汉族,年龄16~18岁。排除标准:根据DSM-IV排除青少年期抑郁症、精神分裂症等、家属中有精神疾病史、严重躯体疾病及脑外伤、物质或药物依赖史。(1)暴力组:官方记录为暴力型违法乱纪的青少年,共107例,MOAS加权分大于或等于4分,且体力攻击大于或等于1分,年龄(16.5±0.6)岁。(2)非暴力组:官方记录为非暴力型违法乱纪的青少年,共107例,

表1 3组DβH基因rs1611115多态性基因型和等位基因频率的比较[n(%)]
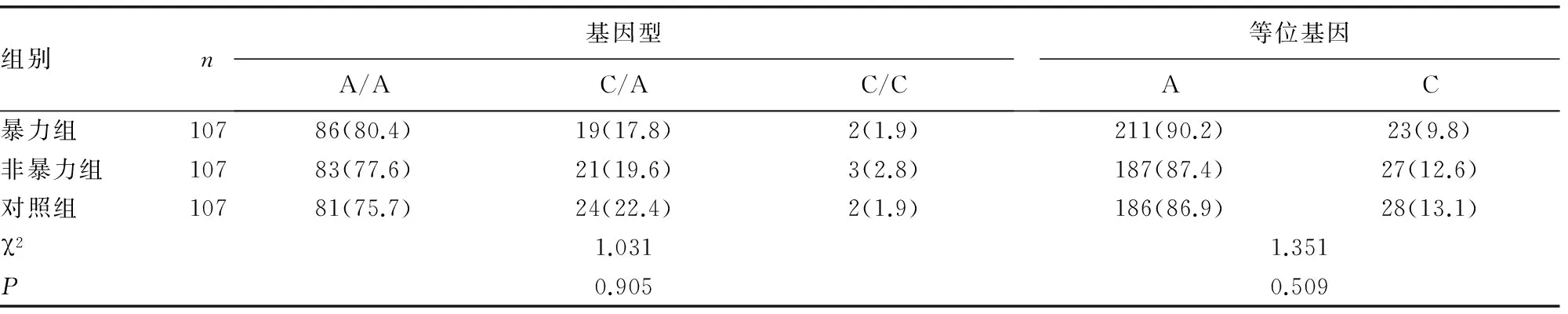
表2 3组DβH基因rs739398多态性基因型和等位基因频率的比较[n(%)]
MOAS加权分小于4分,且体力攻击为0分,年龄(16.6±0.6)岁。(3)对照组:为重庆市渝中区高级职业学校的学生,共107例,MOAS加权分小于4分,且体力攻击为0分,既往无犯罪记录,满足上述入组标准和排除标准,年龄(16.5±0.6)岁。3组在年龄上差异无统计学意义(P>0.05),本研究通过重庆医科大学附属第一医院伦理委员会批准,所有受试者对本研究知情并签署知情同意书。
1.2方法
1.2.1DNA的提取 所有受试者于清晨采集肘静脉血3 mL置于乙二胺四乙酸(EDTA)抗凝管中,使用美国Omega生物技术公司的DNA试剂盒提取DNA。
1.2.2单核苷酸多态性(SNP)分型 所有样本在上海天昊生物公司完成。本研究采用SNaPshot SNP分型技术对所有样本进行SNP位点分型。引物通过在线Primer3 软件设计。
1.2.3PCR循环程序 先95 ℃预变性2 min,在第一相循环94 ℃变性20 s,起始退火温度65 ℃ 40 s,后每增加1个循环下降0.5 ℃,72 ℃延伸90 s,共11个循环,终退火温度为59 ℃。第2相循环94 ℃变性20 s,59 ℃退火30 s,72 ℃延伸1.5 min,共24个循环,72 ℃总延伸2 min,4 ℃保存。 PCR产物通过HotStarTaq进行多重PCR获得,PCR产物通过虾碱酶(SAP)和外切酶Ⅰ(EXOⅠ)纯化后再使用SNaPshot Multiplex试剂盒进行延伸反应。
1.2.4延伸循环程序 先96 ℃预变性1 min,在第一相循环96 ℃变性10 s,退火温度52 ℃ 5 s,60 ℃延伸30 s,共28个循环,4 ℃保存。延伸产物用SAP纯化后在ABI3130XL上样。SNP分型、ABI3130XL测序仪上收集的原始数据均采用GeneMapper4.0(美国Appliedbiosystems公司)来分析。
1.2.5Hardy-Weinberg(H-W)平衡吻合度检验 为了保证本研究样本能代表重庆地区的人群分布,采用χ2检验分别对3组DβH基因rs739398和rs1611115多态性的各基因型分布进行了H-W遗传平衡吻合度检验。

2 结 果
2.1H-W平衡吻合度检验结果 结果发现3组中2个SNP位点的χ2值为0.164~2.439(P均>0.05),符合H-W平衡法则。
2.23组DβH基因rs1611115基因型及等位基因频率的比较 χ2检验显示3组在rs1611115位点基因型及等位基因频率差异均有统计学意义(P<0.05)。两两比较显示暴力组与非暴力组在基因型及等位基因频率上差异均无统计学意义(χ2=0.931、0.000;P>0.05);而与对照组比较,在基因型上差异均有统计学意义 (χ2=8.025、10.224;P<0.05),在等位基因频率上差异也均有统计学意义 (χ2=7.754、7.754;P<0.05),见表1。
2.33组DβH基因rs739398多态性的基因型及等位基因频率的比较 χ2检验显示3组在rs739398位点基因型和基因频率差异均无统计学意义(P>0.05)。两两比较显示暴力组与非暴力组在基因型及等位基因频率差异均无统计学意义(χ2=0.353、0.876;P>0.05);与对照组比较,在基因型上差异均无统计学意义(χ2=0.731、0.541;P均>0.05);在等位基因频率上差异也均无统计学意义(χ2=1.174、0.021;P>0.05),见表2。
3 讨 论
DβH是多巴胺合成去甲肾上腺素的限速酶,对儿茶酚胺的代谢产生重要的影响。2003年白平等[10]发现46例暴力型罪犯的血浆去甲肾上腺素水平明显增高,这些研究表明DβH活性的改变可能与冲动和攻击行为相关。编码DβH的基因定位于9q34,长度接近23 kb,有12个内含子。本研究发现DβH基因rs1611115多态性无论是暴力组还是非暴力组青少年的CC基因型均少于对照组,提示这一多态性位点与暴力行为无关,但提示与暴力和非暴力青少年犯罪均有关。Hess等[11]研究发现,低活性的DβH-1021TT基因型可能与冲动、攻击行为等相关的人格特征有关。DβH对去甲肾上腺素的合成具有限速作用,这可能使脑内多巴胺和去甲肾上腺素的水平发生改变,从而增加一些异常行为及精神疾病发生的风险。但本研究未发现暴力组与非暴力组青少年存在差异,对于青少年罪犯这一特殊群体来说,无论是暴力罪犯还是非暴力罪犯都可能存在着冲动、攻击等个性特征,既往的研究也证实了男性青少年罪犯较对照组具有较高的冲动行为[12]。因此,未来可以在普通人群中尽可能将暴力攻击行为纯化做进一步研究。
本研究未发现DβH基因rs739398多态性与青少年暴力行为存在关联。这可能与DβH-1021C/T或与之连锁的其他多态性是影响DβH活性最主要的多态性有关[13]。Grigorenko等[14]将179名青少年罪犯分为暴力罪犯和非暴力罪犯组,并选取了COMT、DβH、MAOA 及MAOB基因共12个多态性位点进行关联分析,结果未发现单个基因多态性位点在组别中存在差异。同时既往在MAOA、COMT、5-HTT的研究中结合不同的环境因素进行了分析[15-17],并发现环境因素在其中起着重要的作用,尤其是童年有过受虐经历的成年人,携带低活性的MAOA等位基因比携带高活性MAOA等位基因者更容易发生暴力攻击行为。因此,暴力攻击行为可能与基因与环境共同作用有关,未来需进一步联合多基因或环境因素进行分析研究。
总之,本研究发现青少年暴力和非暴力犯罪都与DβH基因rs1611115多态性存在关联,未发现DβH基因rs739398多态性与青少年暴力行为有关。未来将尽可能扩大样本量,将暴力行为纯化,结合环境因素、多基因、多位点进行交互分析。
[1]Conway CC,Keenan-Miller D,Hammen C,et al. Coaction of stress and serotonin transporter genotype in predicting aggression at the transition to adulthood[J]. J Clin Child Adolesc Psychol,2012,41(1):53-63.
[2]于跃,刘祥,杨振兴,等.5-羟色胺转运体基因多态性与青少年暴力犯罪行为的相关性研究[J].中华医学遗传学杂志, 2012,29(4):468-473.
[3]Benis AM,Hobgood DK.Dopamine receptor DRD3 codes for trait aggression as Mendelian recessive[J].Med Hypotheses,2011,77(6):1108-1110.
[4]Hirata Y,Zai CC,Nowrouzi B,et al.Study of the catechol-o-methyltransferase (COMT) gene with high aggression in children[J].Aggress Behav,2013,39(1):45-51.
[5]Vevera J,Stopkova R,Bes M,et al.COMT polymorphisms in impulsively violent offenders with antisocial personality disorder[J].Neuro Endocrinol Lett,2009,30(6):753-756.
[6]黄雄,姜南,林振强,等.COMT基因多态性与精神分裂症暴力攻击行为的关联性[J].广东医学,2010,31(1):82-83.
[7]Zabetian CP,Anderson GM,Buxbaum SG,et al.A quantitative-trait analysis of human plasma dopamine-beta-hydroxylase activity:evidence for a major functional polymorphism at the DβH locus[J].Am J Hum Genet,2001,68(2):515-522.
[8]Rogeness GA,Hernandez JM,Macedo CA,et al.Biochemical differences in children with conduct disorder socialized and undersocialized[J].Am J Psychiatry,1982,139(3):307-311.
[9]谢斌,郑瞻培.修订版外显攻击行为量表//中国行为医学科学编辑委员会.行为医学量表手册[M].北京:中华医学音像出版社,2001:352-353.
[10]白平,傅晓晴,林久茂,等.暴力行为人群性激素水平的异常[J].第四军医大学学报,2003,24(13):1208-1209.
[11]Hess C,Reif A,Strobel A,et al.A functional dopamine-beta-hydroxylase gene promoter polymorphism is associated with impulsive personality styles,but not with a Vective disorders[J].J Neural Transm,2009,116(2):121-130.
[12] 吴岩峰,张建秋,谭钊安,等.245名男性罪犯冲动行为特征分析[J].中华行为医学与脑科学杂志,2009,18(10):927-929.
[13]汤宜朗,Cubells JF,Zabetian CP.多巴胺β-羟化酶与精神疾病[J].上海精神医学,2005,17(1):47-49.
[14]Grigorenko EL,De Young CG,Eastmanl M,et al.Aggressive behavior,related conduct problems,and variation in genes affecting dopamine turnover[J].Aggressive behavior,2010,36(3):158-176.
[15]Frazzetto G,Di Lorenzo G,Carola V,et al.Early trauma and increased risk for physical aggression during adulthood:the moderating role of MAOA genotypen[J].PLoS One,2007,2(5):486-491.
[16]Brennan PA,Hammen C,Sylvers P,et al.Interactions Between the COMT Val108/158Met Polymorphism and Maternal Prenatal Smoking Predict Aggressive Behavior Outcomes[J].Biol Psychol,2011,87(1):99-105.
[17]Reif A,Rosler M,Freitag CM,et al.Nature and nurture predispose to violent behavior:serotonergic genes and adverse childhood environment[J].Neuropsychopharmacology,2007,32(11):2375-2383.
