观点采择在内隐情绪加工中的调节作用*
2014-02-02夏瑞雪周爱保李世峰徐科朋任德云
夏瑞雪 周爱保 李世峰 徐科朋 任德云 朱 婧
(1西北师范大学心理学院, 兰州 730070)
(2北京师范大学认知神经科学与学习国家重点实验室, 北京 100875)
(3河西学院教师教育学院, 甘肃张掖 734000) (4天水卫生学校, 甘肃天水 741000)
1 引言
观点采择是指个人具有区分自己与他人观点的能力, 并能按照过去或当前情境对他人的观点做出正确的判断(Herold & Akhtar, 2008)。换句话说,就是个人设身处地地理解与感受他人想法与情感的能力(Ku, Wang, & Galinsky, 2010)。情感观点采择, 也称共情。通过观点采择可以从不同的视角深入考察情绪加工的机制。尤其是负性情绪, 因其对人类的生存具有重要的生物学意义, 从不同的视角探讨负性情绪的加工是一种可靠的研究路径, 能更细致地揭示人们在社会活动中的共情特征及规律。已有的相关研究主要围绕负性情绪加工偏向和疼痛共情两个方面展开。
1.1 负性情绪加工偏向
情绪效价作为社会信息的重要特征, 影响着人们对信息的感知与加工。研究表明, 相对于中性刺激, 人们对情绪性刺激(正性和负性)的加工更加迅速, 表现为对情绪刺激的反应时缩短(Bradley,Mogg, Millar, & White, 1995; Dong, Zhou, Zhao, &Lu, 2011), 说明情绪刺激能更快地进入认知加工,信息的情绪特征能更快地调动认知资源。而相对于愉快信息, 人对环境中的威胁性信息具有加工偏向,表现为个体对负性刺激的加工更为敏感, 加工时间更为持续, 这种现象被称为“负性情绪偏向(emotional negativity bias)” (Huang & Luo, 2006;Meng, Yuan & Li, 2009)。Huang 和 Luo (2006)在正性、负性和中性条件下观察了P2、LPC和R-LRP (反应锁定的单侧化准备电位)的变化, 结果表明, 负性刺激引发的 P2显著地大于正性刺激引发的 P2,表明对负性刺激的注意优先性发生在早期的情绪知觉阶段; 负性刺激引发了比正性和中性刺激更大的LPC (late positive component), 说明了对负性刺激加工的优先性也发生在后期的情绪评估阶段; 反应锁定的 LRP的间隔在负性刺激的区组中最小,说明了对负性刺激反应准备的持续时间最短, 表明负性刺激引发了反应启动效应。因此, 情绪的负性加工偏向发生在对刺激注意、评估和反应准备等各个阶段(Delplanque, Lavoie, Hot, Silvert, & Sequeira,2004; Delplanque, Silvert, Hot, & Sequeira, 2005;Huang & Luo, 2006)。这类研究要求被试对刺激的效价进行判断, 或者给被试以无关的内隐实验任务。然而, 负性情绪加工偏向的研究只是解释了人们加工不同情绪效价信息时的差异, 由于缺乏观点采择的介入, 研究结果并不能解释人们在社会活动中如何理解自己的情绪和他人情绪。是否能以他人的视角理解他人的情绪, 对成功的社会互动具有重要意义。
1.2 观点采择对疼痛共情的调节作用
当前共情的神经科学研究主要采用疼痛共情范式(潘彦谷, 刘衍玲, 马建苓, 冉光明, 雷浩,2012)。疼痛共情分为早期自下而上的情绪共情和后期自上而下的认知共情两个方面(Fan & Han,2008)。Li和Han (2010)的研究发现, 观点采择调节疼痛共情只出现在后期的控制加工阶段, 不出现在早期的情绪分享阶段。相关影像研究表明, 分别在自我和他人视角条件下判断脚部是否疼痛, 激活了类似但不完全相同的脑区。在他人条件下, 顶叶、前扣带回和前脑岛被激活, 而在自我视角条件下,出现了更广泛的次级感知皮层和整个脑岛的激活(Jackson, Brunet, Meltzoff, & Decety, 2006), 脑岛对负性情绪加工起着非常重要的作用(Vuilleumler,Armony, Driver, & Dolan, 2001; Herbert, Herbert, &Paul, 2011)。这些相关研究结果提示, 人类对自己疼痛和他人疼痛的表征既相似又不同。一方面, 体现了个体与他人共享情绪, 是情绪观察学习的基础,能减少人类对伤害和负性体验的学习, 有利于人类的正常生存。另一方面, 不同视角条件下加工疼痛任务时激活的脑区存在差异, 这提示了人们以不同的立场感知疼痛时其加工机制是不同的。在疼痛共情的研究中, 均要求被试判断看到的身体部位是否疼痛, 这是一种外显的实验任务, 人们在加工刺激时是任务驱动的有意识加工。因此, 该研究结果仅仅反映了观点采择对控制加工阶段的调节效应, 不能对人们无意识加工负性情绪进行有效地解释。
疼痛是一种特殊的负性心理状态。采用疼痛共情范式的研究, 只说明了与疼痛共情相关的生理心理机制。而在日常生活中, 人们要面对大量不同效价的生活事件, 区分这些事件的情绪效价, 判断这些事件是否具有伤害性意义是重要的。成功的社会互动依赖于对他人情绪和感受的理解, 但意识到自我和他人之间的区别也是必需的(Fan & Han,2008)。因此, 通过观点采择研究内隐情绪加工能更清楚地揭示人们在日常生活中面对一般负性情绪时的共情特点及机制。
相关时频分析的研究表明, 情绪性刺激比中性刺激引发更大的theta频带的能量(Bekkedal, Rossi,& Panksepp, 2011); 面孔特定的视觉自我表征引发更大的内侧额叶的事件相关 theta频谱扰动(event-related spectral perturbation) (Miyakoshi,Kanayama, Nomura, Iidaka, & Ohira, 2008); 自我参照加工引发更大的theta节律的同步性(Mu & Han,2010); 自我相关物主代词“我的”引发更大的theta频带的能量活动(周爱保等, 2013)。很显然, theta频带的活动既参与情绪加工, 又参与自我相关加工,那么, theta节律的变化也可能是观点采择调节一般负性情绪加工的指标。
基于上述考虑, 当前研究考察观点采择在内隐情绪加工中的作用。具体思路为:第一, 采用无关的实验任务, 要求被试对情绪图片进行形状判断。被试在执行无关任务时, 不自觉地对刺激的情绪内容进行加工, 是一种内隐情绪任务, 这种任务设置可以减少被试由于猜测实验目的而造成的任务相关效应(黄宇霞, 罗跃嘉, 2009); 第二, 以人们加工负性情绪时神经变化的两个方面:不同的脑电成分和theta节律为指标; 第三, 采用发生在皮肤上的负性事件为刺激材料。结合负性情绪加工偏向和疼痛共情两个方面的研究, 当前研究假设, 观点采择对内隐情绪任务加工的整个过程都具有调节作用。
2 方法
2.1 被试
大学生18名, 均为右利手, 裸视或矫正视力正常。实验前知情同意, 实验后获得一件纪念品。2名被试因脑电伪迹严重, 分析数据时被剔除, 因此有效被试为 16名, 年龄 21~24岁, 平均年龄 22.6岁, 其中男性7名, 女性9名。
2.2 材料
刺激材料包括两大类, 负性图片和中性图片。负性图片选取意义明确且发生在日常生活中的威胁性事件, 如皮肤破损和针灸等。中性图片包括自然风景和人工建筑等。每类选择出 50张, 共 100张图片, 请30名被试对图片的代表性在9点量表上进行评定。依据《中国情绪图片系统的编制》(白露, 马慧, 黄宇霞, 罗跃嘉, 2005), 评定的内容有3个维度:图片的愉悦性, 兴奋性和支配性。评定形式为自我评价模式, 每个等级评定量表中的等级从1到9, 1代表每个维度都是低值(低愉悦度, 低唤醒度, 低优势度), 9代表每个维度都是高值(高愉悦度,高唤醒度, 高优势度)。图片评定时使用E-prime 2.0软件随机呈现, 图片大小为4.55 cm × 5.55 cm, 分辨率为 100像素/英寸, 要求被试按照自己的感受对图片进行评分, 每一维度为一个区组, 所有图片随机呈现, 一个维度评定结束后, 进行下一个维度,3个维度的顺序在被试间平衡。数据使用SPSS 16.0进行统计分析。威胁性图片选择愉悦度较低(M
<=3), 唤醒度较高(M
>= 5)和优势度较低(M
<= 3)的图片, 共20张; 中性图片选择在3个维度上均处于中等水平(3 <=M
<= 6)的图片, 共20张。最终的实验材料确定为 40张图片。图片分别用长方形和正方形两种形状呈现, 长方形图片4.55 cm × 5.55 cm,刺激视角 3.06° × 3.74° (长×宽)。正方形图片 4.55 cm × 4.55 cm, 刺激视角 3.74°× 3.74°。2.3 实验设计
采用2(观点采择:自我视角, 他人视角) × 2(情绪效价:中性图片, 负性图片)的组内设计。其中他人视角中的“他人”确定为一般的, 不熟悉他人。已有研究在呈现刺激时多采用block设计(Li & Han,2010; Cheng, Yang, Lin, Lee, & Decety, 2008), 一个block完成自我判断, 另一个完成他人判断, 需要被试在 block之间转移注意, 而不是试次之间, 不能区分线索启动与情绪加工的过程(Mu & Han,2013)。因此, 当前研究中, 自我视角和他人视角在每个 block中随机呈现, 这就需要被试在试次之间转换自我视角和他人视角。
2.4 实验程序
E-prime实验程序包含以下几个阶段:①指导语阶段, 告知被试实验的目的和注意事项, 确定他人是一般他人, 即一个不熟悉的人。②正式实验阶段, 电脑屏幕上依次呈现注视点“+” (200 ms)、空屏(200 ms)、我在经历/他在经历(500 ms)、空屏(200~300 ms)、刺激图片(1000 ms)和空屏(1400 ms),线索和刺激图片均随机呈现。要求被试对刺激图片进行形状的判断, 在刺激结束后的空屏上做出反应。按键在被试间和被试内平衡。
2.5 数据记录及离线处理
2.5.1 数据记录
使用美国EGI公司256导脑电系统采集被试的EEG。被试离电脑屏幕 85 cm, 眼睛舒适注视屏幕正中, 在线记录数据时以 Cz作为参照电极, 带通为0.1~100 Hz, 采样率为250 Hz, 所有电极的电阻保持在50 kΩ以下。
2.5.2 ERP数据分析
数据离线分析以0.1 Hz为高通, 45 Hz为低通对EEG数据进行滤波; 以刺激呈现之前200 ms、之后1000 ms为一个分段; 若某通导的信号变化超过200 μV, 该通导被标记为伪迹; 一个trial中有超过10个通导被标为伪迹, 剔除该trial数据; 水平眼电信号变化超过140 μV或垂直眼电信号变化超过55 μV, 该 trial数据也被剔除; 坏导替代采用内差值算法, 用临近通导的数据替代坏导; 并对每个trial的数据进行平均参考。然后对每个条件下的trials进行叠加从而获得每个被试在每个条件下的ERPs, 并以刺激前200 ms为基线对ERP波形进行基线校正。根据每种ERP成分的分布特点, 按照国际10-10系统选择分析的电极。P150和N200在额部、中央以及靠近额中央的8, 43和197等导联上均出现了不同条件下ERP的变化。因此, 选择了9个电极(F3, F4, Fz, 8, 43, 197, C3, C4, Cz) (256导联及分析所选电极的分布如图1所示)对P150和N200进行分析。LPC (late positive component)的分布出现了从中央到顶区的区域性变化, 选择中央(C3,C1, Cz, C2, C4)、中央顶(CP3, CP1, CPz, CP2, CP4)和顶区(P3, P1, Pz, P2, P4)共15个电极进行分区统计分析。

3 结果
图1 256网状导联分布图
实验要求被试在刺激消失之后反应, 且实验任务为无关任务, 因此, 没有对反应时和正确率进行分析。
3.1 成分分析
P150 (123 ~179 ms)
对 P150的潜伏期进行 2(观点采择:自我, 他人)× 2(情绪效价:中性, 负性) × 9(电极:F3, F4, Fz,8, 43, 197, C3, C4, Cz)的重复测量方差分析, 结果表明, 条件、效价和电极主效应以及二阶、三阶交互效应均不显著,ps >
0.05; 对P150的波幅进行同样的重复测量的方差分析, 结果发现, 条件、效价和电极主效应以及二阶、三阶交互效应均不显著,ps
> 0.05。N200 (180 ~220 ms)
对N200的潜伏期进行2(观点采择:自我, 他人) × 2(情绪效价:中性, 负性)× 9(电极:F3, F4, Fz,8, 43, 197, C3, C4, Cz)的重复测量方差分析, 结果表明, 效价主效应显著,F
(1, 15)=23.93,p <
0.001,η=0.571, 人们对负性信息的反应显著地短于中性信息; 条件和电极主效应以及二阶、三阶交互效应均不显著,ps >
0.05。对N200的波幅进行同样的重复测量方差分析, 结果表明, 条件和电极的主效应均不显著,ps
> 0.05; 效价主效应显著, F
(1, 15)=13.252,p
= 0.03, η= 0.486, 事后比较发现, 负性刺激引发了比中性刺激更大的波幅。条件和效价之间交互效应显著,F
(1, 15)= 4.655,p
= 0.049, η=0.250, 简单效应分析表明, 对于负性信息, 自我条件与他人条件之间差异显著,F
(1, 15)= 18.006,p <
0.001, η= 0.563。而对于中性信息, 自我和他人之间差异不显著,F
(1, 15)= 1.418,p
> 0.05; 其余二阶和三阶交互效应均不显著,ps
> 0.05。如图2 (选择中线上3个点Fz, 8, Cz点)所示。LPC (267 ~567 ms)

图2 不同加工条件下的总平均波幅
按照中央、中央顶和顶区首先计算不同区域所选电极的平均潜伏期和平均波幅。对平均潜伏期进行 2(观点采择:自我, 他人) × 2(情绪效价:中性, 负性) × 3(分区:中央、中央顶、顶区)重复测量方差分析, 结果表明, 效价主效应显著,F
(2, 30)= 7.606,p
= 0.013, η= 0.297, 负性信息的潜伏期显著地短于中性信息; 分区主效应显著F
(2, 30)= 7.325,p
=0.01, η= 0.289, 配对比较结果显示, 顶区的潜伏期最短(M
= 373.98,SD
= 9.77), 显著短于中央顶(M
= 398.60,SD
= 9.94)和中央(M
= 409.21,SD
=11.63), 而中央顶与中央之间差异不显著; 效价和分区交互效应显著,F
(2, 30)= 3.400,p
= 0.044, η=0.159, 简单效应分析表明, 对于中性信息, 不同区域的潜伏期差异未见显著; 而对于负性信息, 顶区潜伏期最短(M
= 374.84,SD
= 11.16), 其次是中央顶(M
= 405.12,SD
= 11.23), 最长的是中央区(M
=422.32,SD
= 14.30), 且3个区域之间分别存在差异显著。其他二阶及三阶交互效应均不显著,ps >0.05
。对LPC的平均波幅也进行同样的重复测量方差分析, 结果表明, 条件和效价主效应不显著,ps
>0.05; 分区的主效应显著,F
(2, 30)= 18.862,p
=0.001, η= 0.557; 条件和效价的交互效应显著,F
(1,15)=7.852,p
= 0.013, η= 0.344, 简单效应分析表明, 在自我条件上, 中性和负性之间差异临界显著,p
= 0.05; 条件、效价和分区的交互效应显著,F
(2,30)= 5.731,p
= 0.008, η= 0.276, 简单效应分析表明, 在顶区, 自我负性的LPC波幅显著大于他人负性的LPC波幅,p
= 0.042; 其他二阶及三阶交互效应均不显著,ps
> 0.05。3.2 时频分析
相位锁定的时频分析结果显示, 在中央及顶区,theta频带(4~8 Hz)上出现了自我负性比他人负性相对增大的频谱能量, 这种能量的增加大约从刺激呈现开始一直持续到300 ms。为了分析这种效应, 选择9个电极计算了在0~300 ms时间段内每个被试在每种条件下theta (4~8 Hz)频带的能量值。对提取的theta能量值进行2(观点采择:我在经历、他在经历) × 2(情绪效价:中性, 负性) × 9(电极:C3, Cz,C4, CP3, CP4, CPz, P3, Pz, p4)的重复测量方差分析。结果表明, 电极的主效应显著,F
(8, 120)= 7.475,p <
0.001, η=0.318, 条件、效价和电极三阶交互效应显著,F
(8, 120)= 2.421,p
= 0.018, η=0.141, 简单效应分析表明, 在 Cz和 CPz两个电极上, 自我负性引发的能量值显著大于他人负性引发的能量值(如图3所示)。其他主效应及二阶交互效应均不显著,ps
> 0.05。4 讨论
与先前负性情绪的研究结果一致, 当前研究同样发现了人们的负性情绪加工偏向。与中性信息相比, 加工负性信息时N200、LPC潜伏期更短, 波幅更大。N200成分上, 以额部及中央区域为代表,LPC成分上, 以顶区最为典型, 表现了人们对外界不同情绪效价信息的区分能力, 人们能迅速辨别,负性信息, 负性信息拥有一种加工的优先权和反应启动效应。从进化的角度而言, 当危险情境出现时机体需要作出迅速激活战斗/逃跑的行为, 这对于人们在自然界中的生存具有重要意义(黄宇霞,罗跃嘉, 2005; Campanella et al., 2002)。
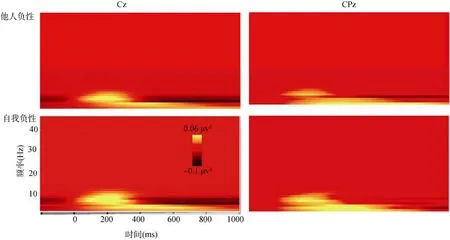
图3 自我负性和他人负性在Cz、CPz上相位锁定的时频能量分布(彩图见电子版)
观点采择调节人们加工内隐情绪任务的整个过程。当前研究结果显示, 与他人视角相比, 在自我视角条件下人们加工负性信息时投入了更多的心理资源, 从早期的N200到晚期的LPC都表现了相同的效应。N200对探测刺激物理属性的变化(Folstein & Pette, 2008)、对探测刺激社会属性的变化(Sui, Hong, Liu, Humphreys, & Han, 2012)以及对负性刺激加工都非常敏感(Yuan et al., 2007)。N200也是注意朝向具有生物学意义事件的标志(Campanella et al., 2002)。当前研究与已有研究相一致(Nagy, Potts, & Loveland, 2003; Yuan et al., 2007),从前额到中央均出现了自动化加工负性信息的偏向。尤其对于自我负性信息, 因其对个体具有重要的适应性价值和意义, 引发了比他人负性条件下显著增大的 N200波幅, 表明自我负性信息在早期自下而上的无意识加工阶段具有优先性; LPC是持续注意、情绪刺激精细加工的指标(Hajcak & Olvet,2008; Huang & Luo, 2006), 也是自我相关加工的重要指标(Zhou et al., 2010; Shi et al., 2011; Miyakoshi,Nomura, & Ohira, 2007)。顶区显著增大的LPC反应了在后期自上而下的控制加工阶段, 自我负性信息加工依然具有优先性。因此, 观点采择对人们无意识加工负性情绪具有稳定的调节作用。
相位锁定的神经振荡反映了连续的 EEG神经节律的变化, 这种变化对于刺激的呈现既锁时又锁相。当前相位锁定的时频分析结果显示, 相对于他人负性条件, 自我负性条件下出现了显著增大的theta节律。反映了 theta节律不仅与情感加工(Putman, van Peer, Maimari, & van der Werff, 2010)、疼痛共情(Mu, Fan, Mao, & Han, 2008)有关, 更重要的是它同时参与情绪加工(Bekkedal et al., 2011)和自我相关加工(Mu & Han, 2010; 周爱保等,2013)。面对负性信息, 在自我视角条件下个体的自传体记忆迅速检索和提取负性经验, 引发了大范围theta节律的同步性(Steinvorth, Wang, Ulbert,Schomer, & Halgren, 2010)。因此, theta节律显著地区分了不同视角条件下对负性情绪的加工。该时频分析结果与成分分析的结果一致地反应了观点采择对负性情绪加工的调节效应。
当前研究结果与 Li和 Han (2010)的研究结果不完全一致, 这说明了观点采择对外显疼痛加工和内隐负性情绪加工的调节存在差异, 机体内隐地加工负性情绪会更稳定地受到观点采择的调节。可以从两个方面去解释这种差异。
第一, 从相关影像研究结果看, 在不同视角条件下加工负性信息有不同脑区的参与。他人视角条件下加工负性信息时更多是认知相关的脑区参与,这些脑区的主要功能是对他人情绪状态的理解和评估。而在自我视角条件下主要是情绪相关脑区的活动, 比如杏仁核和脑岛(Jackson et al., 2006;Lamm, Batson, & Decety, 2007), 以及自我相关加工脑区, 尤其有腹内侧前额叶(vMPFC)的参与。这些脑区是一个人自己的情绪经验表征与提取的重要基础(Herbert, Herbert, & Paul, 2011), 说明自我视角条件下加工负性信息出现了反应启动效应。心理理论的研究表明, 对非相似他人的心理状态的推论激活了背内侧前额叶区域(dMPFC), 该区域主要掌管对信息的认知评价(Mitchell, Macrae, & Banaji,2006), 当前研究中的他人视角采用一个不熟悉的“一般他人”,是与自我非相似的他人。可以断定,他人视角条件下对威胁信息更多是认知相关的理解和评估, 而非情绪经验相关的体验与提取。说明他人视角条件下对负性信息出现了反应抑制效应。这与已有疼痛共情的研究结果相似。他人视角下判断是否疼痛会抑制神经震荡(Cheng et al., 2008),也会降低对疼痛的神经反应(Jackson et al., 2006; Li& Han, 2010)。结合当前研究结果, N200、LPC和theata频带的能量变化都反应了自我视角条件下对负性信息的加工投入了更多的心理资源, 使得自我负性信息会更敏感和精细地被机体加工, 而且这种加工始终得到观点采择的调节。
第二, 实验任务不同所致。不同的实验任务会导致不同的实验结果(Lombardo et al., 2010)。Li和Han (2010)的疼痛共情研究采用外显任务, 是任务驱动的控制性加工。疼痛共情时激活的脑区既有相似, 又有不同, 相似的脑区是早期情绪分享的基础。而在后期的认知评估中, 外显的实验任务使得在不同视角条件下感知疼痛的认知评估出现差异,因此, 观点采择只调节了后期的认知评估阶段(Li& Han, 2010; Lamm et al., 2007; Fan & Han, 2008)。而当前研究采用内隐实验任务, 内隐任务是材料驱动的, 反应机体对材料情绪效价的无意识加工, 在负性情绪加工早期的自动化阶段就已经表现出了观点采择的调节效应。与他人负性信息相比, 自我负性信息在整个加工过程中表现了稳定的加工优势, 体现了自我视角采择可以提高自我监控的能力(Zhou et al., 2013)。
观点采择不仅体现了个体的自我对他人状态的感知和理解, 通过当前研究更加明确地反映了对自我和他人的区分。区分自我与他人是人类生活中最重要的分类, 也是自我意识的核心(David et al.,2006)。自我与他人特异表征的观点认为机体自我表征和他人表征的脑区域是相对独立的, 两者都有各自特定的表征方式和结构(孔繁昌, 张妍, 陈红,2010)。自我观点和行为的表征与内侧前额叶脑区的激活有关(Zhu, Zhang, Fan, & Han, 2007; David et al., 2006), 他人观点和行为的表征与枕颞沟、运动前区、额叶下部及后顶叶相连区域的激活有关(David et al., 2006)。也有研究表明, 机体表征自我心理状态和他人心理状态都出现大脑右半球内侧前额叶区域的激活,但自我条件下的激活程度显著大于他人条件下的激活程度(Lombardo et al.,2010)。内侧前额叶(mPFC)功能性地分离了自我与他人(Mitchell et al., 2006)。这些研究说明人们对自我-他人表征的内在认知机制既重叠又有差异, 重叠使得个体可以通过自己的思想和观点推断他人思想和观点, 而差异则体现了自我的特异性和自我与他人的分离。当前研究与这些结果相呼应, 分别从自我-他人表征的空间特征和时间特征两个方面证实了自我与他人的分离。
综上所述, 人们不仅能区分环境中潜在的威胁信息, 这种能力还受到观点采择的调节。对于一般负性情绪的加工, 观点采择同时调节早期的自动化加工和后期的控制加工, 反应了在威胁情境下自我区别于他人的能力。人通常是自我中心的, 超越自己的立场很难理解别人的心理(Royzman, Cassidy,& Baron, 2003)。依据阿米巴自我理论, 人的皮肤能有力地防御外来的侵害和感染, 是人区分自我与外部世界的界限(Burris & Rempel, 2004, 2010;Legrand, 2007), 当前研究采用发生在皮肤上的负性事件, 研究结果反映了人类的皮肤作为自我与外界的物质界限, 对环境中不同效价信息的区分以及对自我与他人的区分功能, 这对身体自我边界功能的研究具有启示意义。
Bai, L., Ma, H., Huang, Y. X., & Luo, Y. J. (2005). The development of native Chinese affective picture system—a pretest in 46 college students.Chinese Mental Health Journal, 19
, 719–722.[白露, 马慧, 黄宇霞, 罗跃嘉. (2005). 中国情绪图片系统的编制—在46名中国大学生中的试用.中国心理卫生杂志, 19
, 719–722.]Bekkedal, M. Y. V., Rossi, J., & Panksepp, J. (2011). Human brain EEG indices of emotions: Delineating responses to affective vocalizations by measuring frontal theta eventrelated synchronization.Neuroscience and Biobehavioral Reviews, 35
, 1959–1970.Bradley, B. P., Mogg, K., Millar, N., & White, J. (1995).Selective processing of negative information: Effects of clinical anxiety, concurrent depression, and awareness.Journal of Abnormal Psychology, 104
, 532–536.Burris, C. T., & Rempel, J. K. (2004). “It’s the end of the world as we know it”: Threat and the spatial-symbolic self.Journal of Personality and Social Psychology, 86
, 19–42.Burris, C. T., & Rempel, J. K. (2010). If I only had a membrane: A review of amoebic self theory.Social and Personality Psychology Compass, 4
, 756–766.Campanella, S., Gaspard, C., Debatisse, D., Bruyer, R.,Crommelinck, M., & Guerit, J. M. (2002). Discrimination of emotional facial expressions in a visual oddball task: An ERP study.Biological Psychology, 59
, 171–186.Cheng, Y. W., Yang, C. Y., Lin, C. P., Lee, P. L., & Decety, J.(2008). The perception of pain in others suppresses somatosensory oscillations: A magnetoencephalography study.Neuroimage, 40
, 1833–1840.David, N., Bewernick, B. H., Cohen, M. X., Newen, A., Lux,S., Fink, G. R., … Vogeley, K. (2006). Neural representations of self versus other: Visual–spatial perspective taking and agency in a virtual ball-tossing game.Journal of Cognitive Neuroscience, 18
, 898−910.Delplanque, S., Lavoie, M. E., Hot, P., Silvert, L., & Sequeira,H. (2004). Modulation of cognitive processing by emotional valence studied through event-related potentials in humans.Neuroscience Letters, 356
, 1–4.Delplanque, S., Silvert, L., Hot, P., & Sequeira, H. (2005).Event-related P3a and P3b in response to unpredictable emotional stimuli.Biological Psychology, 68
, 107–120.Dong, G. H., Zhou, H., Zhao, X., & Lu, Q. L. (2011). Early negativity bias occurring prior to experiencing of emotion.Journal of Psychophysiology, 25
, 9–17.Fan Y., & Han S. H. (2008). Temporal dynamic of neural mechanisms involved in empathy for pain: An event-related brain potential study.Neuropsychologia, 46
, 160–173.Folstein, J. R., & Petten, C. V. (2008). Influence of cognitive control and mismatch on the Ncomponent of the ERP: A review.Psychophysiology, 45
, 152–170.Hajcak, G., & Olvet, D. M. (2008). The persistence of attention to emotion: Brain potentials during and after picture presentation.Emotion, 8
, 250–255.Herbert, C., Herbert, B. M., & Paul, P. (2011). Emotional self-reference: Brain structures involved in the processing of words describing one’s own emotions.Neuropsychologia,49
, 2947–2956.Herold, K. H., & Akhtar, N. (2008). Imitative learning from a third-party interaction: Relations with self-recognition and perspective taking.Journal of Experimental Child Psychology, 101
, 114–123.Huang, Y. X., & Luo, Y. J. (2005). The reaction priming effect of emotionally negative stimuli: An ERP study.Chinese Journal of Rehabilitation Medicine, 20
, 648–651.[黄宇霞, 罗跃嘉. (2005). 负性情绪刺激的反应启动效应事件相关电位的实验研究.中国康复医学杂志, 20
, 648–651.]Huang, Y. X., & Luo, Y. J. (2006). Temporal course of emotional negativity bias: An ERP study.Neuroscience Letters, 398
, 91–96.Huang, Y. X., & Luo, Y. J. (2009). Can negative stimuli always have the processing superiority?Acta Psychologica Sinica,41
, 822–831.[黄宇霞, 罗跃嘉. (2009). 负性情绪刺激是否总是优先得到加工: ERP 研究.心理学报, 41
, 822–831.]Jackson, P. L., Brunet, E., Meltzoff, A. N., & Decety, J. (2006).Empathy examined through the neural mechanisms involved in imagining how I feel versus how you feel pain.Neuropsychologia, 44
, 752–761.Kong, F. Ch., Zhang, Y., & Chen, H. (2010). Self-with-other representation: Shared representation or special representation?Advances in Psychological Science, 18
,1263–1268.[孔繁昌, 张妍, 陈红. (2010). 自我-他人表征: 共享表征还是特异表征.心理科学进展, 18
, 1263–1268.]Ku, G., Wang, C. S., & Galinsky, A. D. (2010). Perception through a perspective-taking lens: Differential effects on judgment and behavior.Journal of Experimental Social Psychology, 46
, 792–798.Lamm, C., Batson, C. D., & Decety, J. (2007). The neural substrate of human empathy: Effects of perspective-taking and cognitive appraisal.Journal of Cognitive Neuroscience,19
, 42–58.Legrand, D. (2007). Subjectivity and the body: Introducing basic forms of self-consciousness.Consciousness and Cognition, 16
, 577–582.Li, W., & Han, S. H. (2010). Perspective taking modulates event-related potentials to perceived pain.Neuroscience Letters, 469
, 328–332.Lombardo, M. V., Chakrabarti, B., Bullmore, E. T.,Wheelwright, S. J., Sadek, S. A., Suckling, J., … Cohen, S.B. (2010). Shared neural circuits for mentalizing about the self and others.Journal of Cognitive Neuroscience, 22
,1623–1635.Meng, X. X., Yuan, J. J., & Li, H. (2009). Automatic processing of valence differences in emotionally negative stimuli: Evidence from an ERP study.Neuroscience Letters,464
, 228–232.Mitchell, J. P., Macrae, C. N., & Banaji, M. R. (2006).Dissociable medial prefrontal contributions to judgments of similar and dissimilar others.Neuron, 50
, 655–663.Miyakoshi, M., Kanayama, N., Nomura, M., Iidaka, T., &Ohira, H. (2008). ERP study of viewpoint-independence in familiar-face recognition.International
Journal
of Psychophysiology, 69
, 119–126.Miyakoshi, M., Nomura, M., & Ohira, H. (2007). An ERP study on self-relevant object recognition.Brain and Cognition, 63
, 182–189.Mu, Y., & Han, S. H. (2010). Neural oscillations involved in self-referential processing.Neuroimage, 53
, 757–768.Mu, Y., & Han, S. H. (2013). Neural oscillations dissociate between self-related attentional orientation versus evaluation.Neuroimage, 67
, 247–256.Mu, Y., Fan, Y., Mao, L. H., & Han, S. H. (2008).Event-related theta and alpha oscillations mediate empathy for pain.Brain Research, 1234
, 128–136.Nagy, E., Potts, G. F., & Loveland, K. A. (2003). Sex-related ERP differences in deviance detection.International Journal of Psychophysiology, 48
, 285–292.Pan, Y. G., Liu, Y. L., Ma, J. L., Ran, G. M., & Lei, H. (2012).Neurobiological underpinnings of empathy.Advances in Psychological Science, 20
, 2011–2021.[潘彦谷, 刘衍玲, 马建苓, 冉光明, 雷浩. (2012). 共情的神经生物基础.心理科学进展, 20
, 2011–2021.]Putman, P., van Peer, J., Maimari, L., & van der Werff, S.(2010). EEG theta/beta ratio in relation to fear-modulated response-inhibition, attentional control, and affective traits.Biological Psychology, 83
, 73–78.Royzman, E. B., Cassidy, K. W., & Baron, J. (2003). “I know,you know”: Epistemic egocentrism in children and adults.Review of General Psychology, 7
, 38–65.Shi, Z., Zhou, A. B., Liu, P. R., Zhang, P. Y., & Han, W. (2011).An EEG study on the effect of self-relevant possessive pronoun: Self-referential content and first-person perspective.Neuroscience Letters, 494
, 174–179.Steinvorth, S., Wang, C. M., Ulbert, I., Schomer, D., &Halgren, E. (2010). Human entorhinal gamma and theta oscillations selective for remote autobiographical memory.Hippocampus, 20
, 166–173.Sui, J., Hong, Y. Y., Liu, C. H., Humphreys, G. W., & Han, S.H. (2012). Dynamic cultural modulation of neural responses to one’s own and friend’s faces.Social Cognitive and Affective Neuroscience, 8
, 326–332.Vuilleumler, P., Armony, J. L., Driver, J., & Dolan, R. J.(2001). Effects of attention and emotion on face processing in the human brain: An event-related fMRI study.Neuron,30
, 829–841.Yuan, J. J., Zhang, Q. L., Chen, A. T., Li, H., Wang, Q. H.,Zhuang, Z. C. X., & Jia, S. W. (2007). Are we sensitive to valence differences in emotionally negative stimuli?Electrophysiological evidence from an ERP study.Neuropsychologia, 45
, 2764–2771.Zhou, A. B., Li, S. F., Herbert, C., Xia, R. X., Xu, K. P., Xu, Q.Y., … Ren, D. Y. (2013). Perspective taking modulates positivity bias in self-appraisals: Behavioral and event-related potential evidence.Social Neuroscience, 8
, 326–333.Zhou, A. B., Shi, Z., Zhang, P. Y., Liu, P. R., Han, W., Wu, H.F., … Xia, R. X. (2010). An ERP study on the effect of self-relevant possessive pronoun.Neuroscience Letters,480
, 162–166.Zhou, A. B., Li, S. F., Shi, Z. H., Liu, P. R., Xia, R. X., Xu, K.P., … Ren, D. Y. (2013). Searching the self: Encoding self-relevant possessive pronoun and theta activity.Acta Psychologica Sinica, 45
, 790–796.[周爱保, 李世峰, 史战, 刘沛汝, 夏瑞雪, 徐科朋, …任德云. (2013). 寻找自我: 自我相关物主代词的编码与theta节律的活动.心理学报, 45
, 790–796.]Zhu, Y., Zhang, L., Fan, J., & Han, S. H. (2007). Neural basis of cultural influence on self-representation.Neuroimage, 34
,1310–1316.