花青素抗肿瘤作用机制研究进展
2014-01-18李晓岩
崔 建,李晓岩*
(东北林业大学生命科学学院,黑龙江 哈尔滨 150040)
花青素抗肿瘤作用机制研究进展
崔 建,李晓岩*
(东北林业大学生命科学学院,黑龙江 哈尔滨 150040)
花青素是一类广泛存在于植物中的水溶性色素,属于黄酮类化合物,具有保肝,抗肿瘤等多种药理活性。花青素的抗肿瘤机制主要有:抗突变,抗氧化,抗炎,诱导转化,调节信号转导通路抑制肿瘤细胞增殖,诱导细胞周期阻滞,促进肿瘤细胞凋亡,诱导自噬,抗肿瘤侵袭及转移,逆转肿瘤细胞耐药性及增加对化疗的敏感性等。本文就花青素抗肿瘤分子机制的最新研究进展进行综述。
花青素;抗肿瘤;信号传导;分子靶点
花青素是构成花瓣和果实的主要色素之一,普遍存在于植物器官的细胞液中。它是一种天然食用色素,广泛存在于蔬菜和水果中,如紫薯、蓝莓、越橘、葡萄、黑树莓等。
花青素是一类易溶于水的黄酮类化合物,其基本结构是2-苯基苯并吡喃型阳离子(花色基元),在天然物中主要以与葡萄糖、半乳糖、鼠李糖等配体结合成花色苷的形式存在。根据B环上取代基的不同可把植物中常见的花青素分为6 种:天竺葵色素(pelargonidin)、矢车菊色素(cyanidin)、飞燕草色素(delphinidin)、芍药色素(peonidin)、牵牛花色素(petunidin)、锦葵花色素(malvidin)(结构如图1所示)。实验研究表明,花青素B环上的邻二苯酚结构是花青素发挥抑制肿瘤生长与转移作用的活性结构[1-3]。由于花青素具有来源广泛、细胞毒 性小、食用安全等特点,其抗肿瘤作用已成为近年来人们关注的热点[4-6]。本文综述了近10 年来花青素的抗肿瘤分子机制(图2)。

图1 花青素的结构Fig.1 Structure of anthocyanins
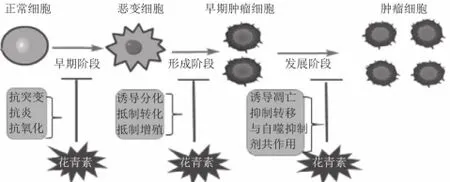
图2 花青素抗肿瘤作用机制Fig.2 Anti-tumor mechanisms of anthocyanins
1 起始阶段
在正常细胞演变为癌细胞的过程中,体细胞多突变的发生会造成基因的不稳定性,进而导致癌症的发生。Yoshimoto等[7]在以4 种不同的甘薯根作为实验材料研究其抗突变作用时发现:鼠伤寒沙门氏菌TA98在杂环诱变剂(3-氨基-1,4-二甲基-5H-吡啶并(4,3-b)吲哚,3-氨基-1-甲基-5H-吡啶并(4,3-b)吲哚,2-氨基-3-甲基咪唑(4,5-f)喹啉等)作用下发生回复突变,而加入4 种不同的甘薯根(其主要成分是矢车菊素糖苷(YGM-3)和芍药花青素糖苷(Y G M-6))能抑制TA 9 8发生回复突变,且抑制效果呈剂量 依赖性。可见,矢车菊素糖苷(YGM-3)和芍药花青素糖苷(YGM-6)具有抑制诱变剂诱导正常细胞发生回复突变的作用。自由基异常引起的氧化应激会导致DNA损伤,会引起相关癌基因的突变,引发癌症。花青素能够作用于抗氧化系统,清除自由基,避免氧化应激引起正常细胞基因组损伤,降低由基因突变导致的细胞恶性转化,预防肿瘤发生[4,8]。Yi Long等[8]研究发现其抗氧化作用是由B环上的3’,4’,5’羟基和C环上的3’羟基决定的。Shih等[4]发现花青素(矢车菊素,飞燕草色素,锦葵色素)能通过能通过Kelch样ECH联合蛋白1-核转录相关因子(NF-E2-related factor 2,Nrf2)途径,作用于抗氧化反应元件(antioxidant response element,ARE),通过调节二相抗氧化酶(谷胱甘肽还原酶,谷胱甘肽过氧化酶,谷胱甘肽转移酶,醌氧化还原酶)的表达,抑制半胱天冬酶-3(cysteinyl aspartate specific proteinase-3,Caspase-3)的活性,从而发挥其抗氧化保护能力。此外,炎症细胞是肿瘤发生的必不可少的参与者。研究表明,花青素能通过多种途径抑制核因子活化B细胞κ轻链增强子(nuclear factor kappa-light-chain-enhancer of activated B cells,NF-κB)的作用实现抗炎。花青素(矢车菊素-3-葡萄糖苷、飞燕草色素-3-葡萄糖苷、牵牛花色素-3-葡萄糖苷等)能通过作用磷脂酰肌醇-3激酶(phosphoinositide 3-kinase,PI3K)/蛋白激酶B(protein kinase,PKB)和丝裂原激活的蛋白激酶(mitogen-activated protein kinases,MAPKs)途径抑制外界刺激(如脂多糖,干扰素-γ等)诱导的NF-κB的活化,抑制环氧合酶(cyclooxygenase 2,COX-2)和诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS)的表达及它们的产物前列腺素E(prostagland in E,PGE2)和一氧化氮的产生[9-10]。Miyake等[11]发现富含花青素越橘提取物能阻止信号转导与转录激活因子3(signal transducer and activator of transcription,STAT3)激活,抑制NF-κB表达。
2 形成阶段
2.1 诱导分化
诱导分化是指恶性肿瘤在诱导分化剂的作用下重新分化向正常成熟方向逆转的现象。恶性肿瘤中存在大量有丝分裂现象,而且分化程度较低,花青素能够诱导肿瘤细胞向终端分化,阻断肿瘤发生。Fimognari等[5]通过检测细胞分化过程中的标记性产物及激酶抑制剂等发现矢车菊素-3-O-葡萄糖苷(cyanidin-3-O-βglucopyranoside,Cy-g)能够通过激活PI3K和蛋白激酶C(protein kinase C,PKC)剂量依赖性诱导人早幼粒急性白血病细胞系HL-60的分化。用Cy-g(质量浓度为200 mg/mL)处理HL-60细胞,发现细胞表现出黏连性增加,酯酶活性增强等分化特征,且细胞内癌基因c-Myc的表达下降;而用PI3K和PKC抑制剂处理细胞后,Cy-g诱导HL-60分化效果明显降低。Serafino等[6]用高效液相色谱法,蛋白免疫印迹法发现矢车菊素-3-O-β-吡喃葡萄糖苷能够通过上调环腺苷酸(cyclic adenosine monophosphate,cAMP)和酪氨酸酶活性来诱导黑色素瘤细胞TVM-A12的分化。
2.2 抑制细胞致瘤性转化
细胞发生致瘤性转化是肿瘤发生的重要原因之一。研究表明,一些致癌剂如对苯二甲酸(terephthalic acid,TPA),表皮生长因子(epidermal growth factor,EGF)会通过上游激活蛋白(Raf)-MAPK/ERK激酶(MEK)-胞外调节蛋白激酶(extracellular regulated protein kinases,ERK)和PI3K/Akt途径细胞致瘤转化关键的活化蛋白转录因子(ectivator protein transcription factor-1,AP-1)和NF-κB来实现诱导各种细胞系的转化。此外,炎症发生与细胞致瘤转化也有重要联系,COX-2和PGE2高表达会增强致癌作用[1]。花青素可以作用于上游激活蛋白(Ras)-ERK和PI3K/Akt途径,抑制AP-1的表达从而抑制细胞的致瘤性转化。Hou等[1]发现飞燕草素、矢车菊素、牵牛色素能够抑制TPA诱导的小鼠皮肤细胞JB6P+的转化。Kang等[12]研究发现,飞燕草素能以三磷酸腺苷(adenosine triphosphate,ATP)非竞争性方式与Raf1和MEK1结合,抑制TPA诱导的JB6P+细胞中AP-1和NF-κB的表达,进 而抑制COX-2表达和PEG2的产生。实验检测了MEK,ERK,核糖体蛋白S6激酶,促分裂原应力激活蛋白激酶的磷酸化水平,发现飞燕草素通过Ras/Raf/ MEK/ERK途径弱化TPA诱导细胞的致瘤性转化作用。早期实验表明,这与花青素能清除超氧自由基的抗氧化能力有关[1]。最近Song等[13]研究发现矢车菊素和矢车菊素-3-葡萄糖苷(cyanidin-3-glucoside,C-3-G)的1,1-二苯基-2-三硝基苯肼和2,2-联氮-二(3-乙基-苯并噻唑-6-磺酸)二胺盐清除能力相差不大,但是抑制致瘤性转化能力有很大差别:矢车菊素具有强烈抑制转化的能力,而C-3-G几乎没有抑制转化的能力,这说明矢车菊素的抑制转化作用与其抗氧化能力无关。此研究中还发现,矢车菊素能以ATP竞争性方式直接与PI3K结合,通过PI3K/ Akt/p70S6激酶信号途径抑制AP-1和NF-κB的表达,抑制EGF诱导的JB6P+的致瘤性转化。
2.3 抑制细胞增殖
癌细胞的一个显著特点就是细胞周期调控异常,能够持续分裂与增殖。研究发现,花青素能够选择性抑制癌细胞的增殖,而对正常细胞的增殖影响较小[14]。花青素能够抑制癌细胞生长和增殖主要表现在以下几方面。
2.3.1 抑制酪氨酸蛋白激酶受体途径阻断信号传导
2.3.1.1 肝细胞生长因子受体
Syed等[15]研究发现,飞燕草色素能抑制肝细胞生长因子诱导的人正常乳腺细胞MCF-10A肝细胞生长因子受体的磷酸化和激活,阻断Ras-ERK MAPK途径和PI3K/ Akt信号途径。
2.3.1.2 表皮生长因子受体
花青素能普遍抑制癌细胞中受体酪氨酸激酶(receptor tyrosine kinases,RTKs)自身磷酸化,且对癌基因ErbB3抑制最有效[16]。
2.3.1.3 抑制cAMP-PDE活性
花青素尤其是锦葵色素能够有效抑制人结肠癌HT29细胞内磷酸二酯酶(phosphodiesterase,PDE)的活性和cAMP的水解,进而抑制MAPK信号途径[2]。
2.3.2 调节癌基因及相关蛋白的表达
Malik[14]和Yun[17]等发现花青素上调结肠癌细胞内p53活化DNA修复系统同时启动p21和p27的转录,p21能与多种细胞周期蛋白-细胞周期蛋白依赖性激酶(cyclincyclin-dependent kinase,cyclin-CDK)结合并抑制其活性,诱导癌细胞周期停滞。花青素能够下调CDK-1、CDK-2表达,抑制cyclin-B、-A、-E的表达,促进周期蛋白依赖性激酶抑制因子表达,诱导癌细胞停滞在G0/G1期和G2/M期[14,17-18]。
2.3.3 其他信号途径
Kausar等[19]发现飞燕草色素能作用于Wnt途径和Notch途径及其下游靶蛋白,抑制细胞生长增殖。
3 发展阶段
3.1 诱导肿瘤细胞凋亡
恶性转化的细胞由于获得了生长失控的特性,从而过度增殖形成肿瘤。从细胞凋亡角度来看,是肿瘤细胞的凋亡过程受到抑制,不能正常清除死亡细胞的结果。花青素能够通过内部线粒体途径和外部死亡受体途径诱导癌细胞的凋亡。
3.1.1 死亡受体途径
Huang Huipei等[20]用p38抑制剂SB203580处理检测p38、p53、蛋白Fas、Fasl的磷酸化水平,证实桑葚花青素能够通过外在受体p38/Fas/Fasl/Caspase-8途径诱导胃癌细胞凋亡。Chang等[21]研究发现,飞燕草素能激活p38-Fasl和促细胞凋亡因子Bid蛋白途径,以时间和剂量依赖的方式诱导HL-60细胞的凋亡。
3.1.2 线粒体信号途径
线粒体介导细胞凋亡的途径分为Caspase依赖途径和非Caspase依赖途径。
3.1.2.1 Caspase依赖途径
Lee[22]和Shin[23]等发现花青素作用于B淋巴细胞瘤(B-cell lymphoma,Bcl)家族和细胞凋亡抑制蛋白家族,激活Caspase依赖的级联凋亡反应。
3.1.2.2 非Caspase依赖途径
Reddivari等[24]发现从马铃薯中提取的花青素成分能够通过c-Jun氨基末端激酶(c-Jun amino-terminal kinase,JNK)途径(不引起氧化应激反应),诱导线粒体释放核酸内切酶G和凋亡诱导因子蛋白,引发前列腺癌LNCaP和PC-3细胞系不依赖Caspase的凋亡。
3.2 与自噬抑制剂共作用
自噬是细胞内的物质成分利用溶酶体被降解过程,在癌症的发生过程中起到双重作用:一方面,自噬缺陷会促进恶性转化和癌症发生;另一方面,自噬能限制肿瘤坏死和炎症,减轻了肿瘤细胞内的染色体损伤代谢性应激反应。研究发现,花青素能够诱导细胞以自噬相关蛋白5(autophagy related protein 5,Atg5)依赖的方式发生自噬[25]。Longo等[26]发现用花青素处理人肝癌细胞PLC/PRF/5后,引起下游Bcl-2家族和雷帕霉素靶蛋白的表达下调,真核细胞翻译起始因子2α表达上调,最终导致自噬相关基因LC3-Ⅱ表达上调,这些现象都表明花青素能诱导人肝癌细胞HCC细胞自噬的发生。用小干涉RNA(small interfering RNA,siRNA)干扰Atg5基因沉默或者用自噬抑制剂3-甲基腺嘌呤作用花青素处理过的肝癌细胞会引起凋亡蛋白Bax转移到线粒体中,释放细胞色素c,引起Caspase-3裂解,激发级联凋亡反应。Feng Rentian等[27]发现矢车菊素-3-芸香糖苷和飞燕草素不能诱导HCC细胞的凋亡,只能引起细胞生长迟缓,出现细胞液泡化现象,这种现象可以被Ⅲ型PI3K抑制剂3-甲基腺嘌呤和阻断溶酶体降解的质子泵抑制剂抑制。在用3-MA和siRNA干扰自噬标志产物LC3后,用飞燕草素处理人肝癌SMMC7721细胞,发现飞燕草素能够诱导SMMC7721细胞发生自噬,而且3-MA和飞燕草素共同作用SMMC7721细胞会导致细胞大量死亡。
3.3 抑制肿瘤细胞的转移
在肿瘤转移这个过程,肿瘤细胞的增殖和侵袭是最为重要的两个部分。研究发现,花青素能作用于血管内皮生长因子及其受体(vascular endothelial growth factor/vascular endothelial growth factor receptor,VEGF/ VEGFR) 和细胞外基质(extra cellular matrix,ECM)的降解来有效抑制肿瘤的侵袭,因抑制肿瘤细胞增殖的机制在上面已经阐述过,在此不再赘述。
3.3.1 抑制肿瘤血管的形成
肿瘤的血管生成是恶性肿瘤生长和转移的限制性条件,其过程受到多种细胞因子的调控,其中最重要的正向调控因子是血管内皮生长因子(VEGF)。因此抑制促血管发生的受体——VEGFR可以有效抑制肿瘤的转移。研究表明,花青素能够广泛抑制RTK,对VEGFR-3抑制作用尤为明显[18,28]。Oak等[29]研究发现,飞燕草色素和矢车菊色素能通过阻断p38-MAPK和JNK途径强烈抑制血小板源性生长因子(platelet derived growth factor,PDGFAB)诱导血管平滑肌细胞内VEGF的表达。Lamy等[30]发现飞燕草色素能够抑制血管的生成不仅与抑制VEGF有关,还和其抑制PDGFR-β的作用有关。飞燕草色素能以时间和剂量依赖性抑制PDGF诱导的肺动脉平滑肌细胞中PDGFR-β的磷酸化,抑制VEGF。
诱导的人类脐静脉内皮细胞HUVEC的微血管的形成。低氧环境是实体瘤普遍具有的病理生理特征,低氧容易诱导肿瘤血管的生成,而这一过程主要由低氧诱导因子(hypoxia inducible factor-1α,HIF-1α)介导的VEGF信号途径完成。Huang Lili等[31]发现抑制HIF-1α的蛋白水平会导致包括VEGF在内的HIF-1α的靶基因的转录活性下降。Wang Lishu等[32]发现冷冻干燥的黑莓花青素能够降低N-甲基苄基亚硝胺诱导的F344大鼠食道瘤细胞内HIF-1α和VEGF的表达,抑制食道瘤血管的生成。
3.3.2 抑制肿瘤的侵袭与转移
侵袭和转移是癌症威胁患者健康乃至生命的主要原因。Xu Mei等[3]发现矢车菊素通过阻断ErbB2/原癌基因cSrc/黏着斑激酶 (focal adhesion kinase,FAK)途径,可以有效抑制ErbB2高表达的乳腺癌细胞(BT474、MDA-MB231、MCF-7)的侵袭和转移。Syed等[16]发现飞燕草色素能够降低肝细胞生长因子介导的MCF-10A细胞系内PKCα的膜转位和STAT3的磷酸化水平,抑制NF-κB/ p65的核转位,从而抑制该细胞的侵袭。侵袭转移过程涉及黏附、降解、运动3 个主要过程。花青素能够作用于肿瘤细胞的一些黏附分子和蛋白水解酶来抑制细胞的黏附和降解过程。
3.3.2.1 黏附
Chen Chunye等[33]研究发现飞燕草色素通过活性氧(reactive oxygen species,ROS)/p38-MAPK/NF-κB途径,剂量依赖性降低氧化低密度脂蛋白诱导的细胞黏附分子-1和P-选择素的表达,从而抑制单核细胞对内皮细胞的黏附。
3.3.2.2 降解
尿激酶型纤溶酶原激活剂(uridylyl phosphate adenosine,uPA)和基质金属蛋白酶(matrix metallo preteinases,MMPs)是细胞外基质降解的两个关键组分,它们可以促进肿瘤组织侵袭,增加迁移能力。花青素靶向作用于uPA和MMPs,从而起到抑制肿瘤转移的作用。
1)尿激酶型纤溶酶原激活剂(uPA)
Lamy等[34]发现飞燕草素能够通过干扰恶性胶质瘤U-87细胞内uPA-纤溶酶激活因子抑制剂复合物的清除,影响uPA 受体和低密度脂蛋白受体相关蛋白的表达及纤溶酶的产生,发挥其抑制U-87细胞侵袭的能力。Chen Peini等[35]发现黑米花青素能够通过抑制肝癌细胞AP-1和NF-κB的表达来抑制uPA的表达,从而发挥抑制癌细胞侵袭和转移的功能。Ho等[36]发现芍药色素能通过抑制ERK1/2的磷酸化,从而降低肺癌细胞内uPA的表达,发挥其抑制癌细胞侵袭的作用。
2)ECM消化酶(MMPs)
Lim等[37]发现飞燕草色素能以还原型烟酰胺腺嘌呤二核苷酸磷酸氧化酶为靶点,抑制MAPK激酶-JNK1/2、MKK3/6-p38和MEK-ERK1/2的磷酸化,从而抑制紫外线B辐射诱导的人表皮纤维母细胞内MMP-1的表达。Shin[38]和Huang Huipei[39]等发现矢车菊素及天竺葵色素等通过抑制癌细胞内PI3K/Akt途径来抑制MMP-2、MMP-9的表达,从而发挥其抑制癌细胞侵袭的作用。研究还发现,紫葛花青素能通过抑制癌细胞中NF-κB的激活来抑制MMP-2、MMP-9的表达[40]。此外,研究发现,花青素(飞燕草素-3-葡萄糖苷、矢车菊素-3-葡萄糖苷、锦葵色素-3-葡萄糖苷等)能够通过激活人结肠癌细胞HCT-116内p38-MAPK途径,抑制紧密连接蛋白Claudin-1和Claudin-3的表达,进一步抑制HCT-116的侵袭[38]。
4 逆转多药耐药性
化疗是通过干扰肿瘤细胞复制DNA的能力而抑制其增生或诱导其凋亡来达到疗效,而肿瘤细胞的多药耐药性(multidrug resistance)是导致化疗失败的常见原因。引起肿瘤细胞多药耐药性的经典途径是由ATP结合盒式蛋白(ATP-binding cassette,ABC)跨膜蛋白超家族引起的,主要包括P-糖蛋白(P-glycoprotein,P-gp),多药耐药相关蛋白,乳腺癌耐药蛋白(breast cancer resistance protein,BCRP)引起的。ABC超家族在药理学屏障中高度表达。因此,以这些蛋白为靶点逆转多药耐药性可以辅助化疗从而达到治疗癌症的目的。实验表明,花青素具有小分子结构,能够透过血脑屏障[41]。进一步研究发现,氯化矢车菊素能够抑制过度表达P-gp的人表皮癌细胞KB-C2中P-gp的表达[42]。Dreiseitel等[43]发现花青素对BCRP具有高亲和力,所测花青素中,7 种(锦葵色素、牵牛花色素、锦葵色素-3-半乳糖苷、锦葵花苷、矢车菊素-3-半乳糖苷、芍药素-3-葡萄糖苷、矢车菊素-3-葡萄糖苷)表现为BCRP的潜在底物,12 种(矢车菊素、芍药花色素、矢车菊素-3,5-二葡萄糖苷、锦葵色素、天竺葵色素、飞燕草色素、牵牛花色素、飞燕草素-3-葡萄糖苷、矢车菊素-3-芸香糖苷、锦葵色素-3-葡萄糖苷、天竺葵色素-3,5-二葡萄糖苷、锦葵色素-3-半乳糖苷)表现出BCRP抑制剂特性。还有3 种(锦葵色素、锦葵色素-3-半乳糖苷、牵牛花色素)表现出双重生物活性。Esselen等[44]在体外实验中发现黑米花青素能够抑制结肠癌HT29细胞内的拓扑异构酶的活性以及拓扑异构酶-DNA复合物的形成。这些结果表明,花青素可能具有改变药物代谢动力学,逆转多药耐药性的作用。
5 结 语
花青素B环上的取代基不同,抗癌作用不同,B环上有邻二苯酚结构的花青素显示出最强的抗肿瘤活性[1,8]。大量研究表明,花青素抗癌的主要机制是以RTKs(EGFR、PDGFR、VEGF/VEGFR等)为靶点,作用于Ras-MAPK和PI3K/Akt信号级联途径,进而抑制肿瘤的生长与转移。在起始阶段:花青素作用于PI3K/Akt和NF-κB途径,抑制COX-2和iNOS的表达,实现抗炎作用和通过Nrf2/ARE信号系统,通过调节二相抗氧化酶的表达实现抗氧化,最终实现预防癌变的作用;在形成阶段:花青素靶向MAPK途径和AP-1因子,抑制细胞的致瘤性转化,还能抑制RTK活性及其信号级联通路调节癌基因表达,引起细胞周期阻滞,修复DNA,防止癌症的发生;在发展阶段:花青素能通过ROS和JNK/p38-MAPK介导的Caspase活化诱导肿瘤细胞凋亡,另外花青素还靶向VEGF信号通路和ECM的降解实现抑制肿瘤的转移。此外,花青素还能逆转肿瘤细胞的多药抗药性,提高化疗敏感性。
现有的大量实验数据表明,花青素可以干扰多种信号通路,具有抗癌活性。然而必须指出的是这些实验大多都是体外实验,而且在人流行病学研究中并没有显示花青素的摄取与罹患癌症风险率的必要联系[45]。不过,建立的动物模型以及体外实验具有指示性,表明了花青素潜在的预防和治疗性抗癌作用。因此需要更多的临床实验证明花青素可以应用于人体的抗癌活性。此外,实验显示花青素的人体利用度低于体外起作用的花青素浓度[46],所以如何提高花青素的体内生物利用度以及稳定性也是进一步需要研究的问题。
[1] HOU D X, KAI K, LI J J, et al. Anthocyanidins inhibit activator protein 1 activity and cell transformation: structure-activity relationship and molecular mechanisms[J]. Carcinogenesis, 2004, 25(1): 29-36.
[2] MARKO D, PUPPEL N, TJADEN Z, et al. The substitution pattern of anthocyanidins affects different cellular signaling cascades regulating cell proliferation[J]. Molecular Nutrition & Food Research, 2004, 48(4): 318-325.
[3] XU Mei, BOWER K A, WANG Siying, et al. Cyanidin-3-glucoside inhibits ethanol-induced invasion of breast cancer cells overexpressing ErbB2[J]. Molecular Cancer, 2010, 9(1): 285.
[4] SHIH P H, YEH C T, YEN G C. Anthocyanins induce the activation of phase II enzymes through the antioxidant response element pathway against oxidative stress-induced apoptosis[J]. Journal of Agricultural and Food Chemistry, 2007, 55(23): 9427-9435.
[5] FIMOGNARI C, BERTI F, NÜSSE M, et al. Induction of apoptosis in two human leukemia cell lines as well as differentiation in human promyelocytic cells by cyanidin-3-O-beta-glucopyranoside[J]. Biochemical Pharmacology, 2004, 67: 2047-2056.
[6] SERAFINO A, SINIBALDI-VALLEBONA P, LAZZARINO G, et al. Differentiation of human melanoma cells induced by cyanidin-3-O-βglucopyranoside[J]. The FASEB Journal, 2004, 18(15): 1940-1942.
[7] YOSHIMOTO M, OKUNO S, YOSHINAGA M, et al. Antimutagenicity of sweet potato (Ipomoea batatas) roots[J]. Bioscience, Biotechnology, and Biochemistry, 1999, 63(3): 537-541.
[8] YI Long, CHEN Chunye, JIN Xin, et al. Structural requirements of anthocyanins in relation to inhibition of endothelial injury induced by oxidized low-density lipoprotein and correlation with radical scavenging activity[J]. FEBS Letters, 2010, 584(3): 583-590.
[9] JEONG J W, LEE W S, SHIN S C, et al. Anthocyanins downregulate lipopolysaccharide-induced inflammatory responses in BV2 microglial cells by suppressing the NF-κB and Akt/MAPKs signaling pathways[J]. International Journal of Molecular Sciences, 2013, 14(1): 1502-1515.
[10] HASEEB A, CHEN D, HAQQI T M. Delphinidin inhibits IL-1βinduced activation of NF-κB by modulating the phosphorylation of IRAK-1(Ser376) in human articular chondrocytes[J]. Rheumatology, 2013, 52(6): 998-1008.
[11] MIYAKE S, TAKAHASHI N, SASAKI M, et al. Vision preservation during retinal inflammation by anthocyanin-rich bilberry extract: cellular and molecular mechanism[J]. Laboratory Investigation, 2012, 92(1): 102-109.
[12] KANG N J, LEE K W, KWON J Y, et al. Delphinidin attenuates neoplastic transformation in JB6 Cl41 mouse epidermal cells by blocking Raf/mitogen-activated protein kinase kinase/extracellular signal-regulated kinase signaling[J]. Cancer Prevention Research, 2008, 1(7): 522-531.
[13] SONG N R, YANG H, PARK J, et al. Cyanidin suppresses neoplastic cell transformation by directly targeting phosphatidylinositol 3-kinase[J]. Food Chemistry, 2012, 133(3): 658-664.
[14] MALIK M, ZHAO Cuiwei, SCHOENE N, et al. Anthocyanin-rich extract from Aronia meloncarpa E. induces a cell cycle block in colon cancer but not normal colonic cells[J]. Nutrition and Cancer, 2003, 46(2): 186-196.
[15] SYED D N, AFAQ F, SARFARAZ S, et al. Delphinidin inhibits cell proliferation and invasion via modulation of Met receptor phosphorylation[J]. Toxicology and Applied Pharmacology, 2008, 231(1): 52-60.
[16] TELLER N, THIELE W, MARCZYLO T H, et al. Suppression of the kinase activity of receptor tyrosine kinases by anthocyaninrich mixtures extracted from bilberries and grapes[J]. Journal ofAgricultural and Food Chemistry, 2009, 57(8): 3094-3101.
[17] YUN J M, AFAQ F, KHAN N, et al. Delphinidin, an anthocyanidin in pigmented fruits and vegetables, induces apoptosis and cell cycle arrest in human colon cancer HCT116 cells[J]. Molecular Carcinogenesis, 2009, 48(3): 260-270.
[18] CHEN Peini, CHU Shuchen, CHIOU H L, et al. Cyanidin 3-glucoside and peonidin 3-glucoside inhibit tumor cell growth and induce apoptosis in vitro and suppress tumor growth in vivo[J]. Nutrition Cancer, 2005, 53(2): 232-243.
[19] KAUSAR H, JEYABALAN J, AQIL F, et al. Berry anthocyanidins synergistically suppress growth and invasive potential of human nonsmall-cell lung cancer cells[J]. Cancer Letter, 2012, 325(1): 54-62.
[20] HUANG Huipei, CHANG Y C, WU C H, et al. Anthocyanin-rich mulberry extract inhibit the gastric cancer cell growth in vitro and xenograft mice by inducing signals of p38/p53 and c-jun[J]. Food Chemistry, 2011, 129(4): 1703-1709.
[21] CHANG Y C, HUANG Huipei, HSU J D, et al. Hibiscus anthocyanins rich extract-induced apoptotic cell death in human promyelocytic leukemia cells[J]. Toxicology and Applied Pharmacology, 2005, 205(3): 201-212.
[22] LEE S H, PARK S M, PARK S M, et al. Induction of apoptosis in human leukemia U937 cells by anthocyanins through down-regulation of Bcl-2 and activation of caspases[J]. International Journal of Oncology, 2009, 34(4): 1077-1083.
[23] SHIN D Y, LEE W S, LU Jingnan, et al. Induction of apoptosis in human colon cancer HCT-116 cells by anthocyanins through suppression of Akt and activation of p38-MAPK[J]. International Journal of Oncology, 2009, 35(6): 1499-1504.
[24] REDDIVARI L, VANAMALA J, CHINTHARLAPALLI S, et al. Anthocyanin fraction from potato extracts is cytotoxic to prostate cancer cells through activation of caspase-dependent and caspaseindependent pathways[J]. Carcinogenesis, 2007, 28(10): 2227-2235.
[25] TSUYUKI S, FUKUI S, WATANABE A, et al. Delphinidin induces autolysosome as well as autophagosome formation and delphinidininduced autophagy exerts a cell protective role[J]. Journal of Biochemical and Molecular Toxicology, 2012, 26(11): 445-453.
[26] LONGO L, PLATINI F, SCARDINO A, et al. Autophagy inhibition enhances anthocyanin-induced apoptosis in hepatocellular carcinoma[J]. Molecular Cancer Therapeutics, 2008, 7(8): 2476-2485.
[27] FENG Rentian, WANG S Y, SHI Yinghong, et al. Delphinidin induces necrosis in hepatocellular carcinoma cells in the presence of 3-methyladenine, an autophagy inhibitor[J]. Journal of Agricultural and Food Chem istry, 2010, 58(7): 3957-3964.
[28] TELLER N, THIELE W, BOETTLER U, et al. Delphinidin inhibits a broad spectrum of receptor tyrosine kinases of the ErbB and VEGFR family[J]. Molecular Nutrition & Food Research, 2009, 53(9): 1075-1083.
[29] OAK M H, BEDOUI J E, MADERIRA S V F, et al. Delphinidin and cyanidin inhibit PDGFAB-induced VEGF release in vascular smooth muscle cells by preventing activation of p38 MAPK and JNK[J]. British Journal of Pharmacology, 2006, 149(3): 283-290.
[30] LAMY S, BEAULIEU É, LABBÉ D, et al. Delphinidin, a dietary anthocyanidin, inhibits platelet-derived growth factor ligand/receptor (PDGF/PDGFR) signaling[J]. Carcinogenesis, 2008, 29(5): 1033-1041.
[31] HUANG Lili, ZHANG Zefu, ZHANG Shen, et al. Inhibitory action of celastrol on hypoxia-mediated angiogenesis and metastasis via the HIF-1α pathway[J]. International Journal of Molecular Medicine, 2011, 27(3): 407-415.
[32] WANG Lishu, HECHT S S, CARMELLA S G, et al. Anthocyanins in black raspberries prevent esophageal tumors in rats[J]. Cancer Prevention Research, 2009, 2(1): 84-93.
[33] CHEN Chunye, YI Long, JIN Xin, et al. Inhibitory effect of delphinidin on monocyte-endothelial cell adhesion induced by oxidized low-density lipoprotein via ROS/p38MAPK/NF-κB pathway[J]. Cell Biochemistry and Biophysics, 2011, 61(2): 337-348.
[34] LAMY S, LAFLEUR R, BÉDARD V, et al. Anthocyanidins inhibit migration of glioblastoma cells: structure-activity relationship and involvement of the plasminolytic system[J]. Journal of Cellular Biochemistry, 2007, 100(1): 100-111.
[35] CHEN Peini, KUO W H, CHIANG C L, et al. Black rice anthocyanins inhibit cancer cells invasion via repressions of MMPs and u-PA expression[J]. Chemico-Biological Interactions, 2006, 163(3): 218-229.
[36] HO M L, CHEN Peini, CHU Shunchen, et al. Peonidin 3-glucoside inhibits lung cancer metastasis by downregulation of proteinases activities and MAPK pathway[J]. Nutrition and Cancer, 2010, 62(4): 505-516.
[37] LIM T G, JUNG S K, KIM J, et al. NADPH oxidase is a novel target of delphinidin for the inhibition of UVB-induced MMP-1 expression in human dermal fibroblasts[J]. Experimental Dermatology, 2013, 22(6): 428-430.
[38] SHIN D Y, LU Jingnan, KIM G Y, et al. Anti-invasive activities of anthocyanins through modulation of tight junctions and suppression of matrix metalloproteinase activities in HCT-116 human colon carcinoma cells[J]. Oncology Reports, 2011, 25(2): 567-572.
[39] HUANG Huipei, SHIH Y W, CHANG Y C, et al. Chemoinhibitory effect of mulberry anthocyanins on melanoma metastasis involved in the Ras/PI3K pathway[J]. Journal of Agricultural and Food Chemistry, 2008, 56(19): 9286-9293.
[40] YUN J W, LEE W S, KIM M J, et al. Characterization of a profile of the anthocyanins isolated from Vitis coignetiae Pulliat and their antiinvasive activity on HT-29 human colon cancer cells[J]. Food and Chemical Toxicology, 2010, 48(3): 903-909.
[41] ANDRES-LACUEVA C, SHUKITT-HALE B, GALLI R L, et al. Anthocyanins in aged blueberry-fed rats are found centrally and may enhance memory[J]. Nutritional Neuroscience, 2005, 8(2): 111-120.
[42] KITAGAWA S. Inhibitory effects of polyphenols on p-glycoprotein-mediated transport[J]. Biological and Pharmaceutical Bulletin, 2006, 29(1): 1-6.
[43] DREISEITEL A, OOSTERHUIS B, VUKMAN K V, et al. Berry anthocyanins and anthocyanidins exhibit distinct affinities for the efflux transporters BCRP and MDR1[J] . Biological and Pharmaceutical Bulletin, 2009, 158(8): 1942-1950.
[44] ESSELEN M, BOETTLER U, TELLER N, et al. Anthocyaninrich blackberry extract suppresses the DNA-damaging properties of topoisomerase I and II poisons in colon carcinoma cells[J]. Journal of Agricultural and Food Chemistry, 2011, 59(13): 6966-6973.
[45] WANG Lishu, STONER G D. Anthocyanins and their role in cancer prevention[J]. Cancer Letter, 2008, 269(2): 281-290.
[46] COOKE D, STEWARD W P, GESCHER A J, et al. Anthocyans from fruits and vegetables-does bright colour signal cancer chemopreventive activity?[J]. European Journal of Cancer, 2005, 41(13): 1931-1940.
Progress on Anti-tumor Mechanisms of Anthocyanins
CUI Jian, LI Xiao-yan*
(College of Life Sciences, Northeast Forestry University, Harbin 150040, China)
Anthocyanins are a class of water soluble flavonoids and show some pharmacological activities such as hepatoprotective activity and anti-tumor activity. The potential anti-tumor mechanism mainly include anti-mutagenesis, antioxidant, anti-inflammatory, induction of differentiation, inhibiting proliferation by modulating signal transduction pathways, inducing cell cycle arrest and stimulating apoptosis of tumor cells, inducing autophagy, anti-invasion and metastasis, reversing drug resistance of tumor cells as well as increasing sensitivity to chemotherapy. In this review, the latest progresses on the anti-tumor activities of anthocyanins and the underlying molecular mechanisms are summarized.
anthocyanins; anti-tumor; signal transduction; molecular target
TS209
A
1002-6630(2014)13-0310-06
10.7506/spkx1002-6630-201413060
2014-02-24
中国博士后科学基金项目(2013M541328)
崔建(1993—),女,本科生,主要从事天然抗肿瘤活性产物的研发。E-mail:yiluxiangbei2012@live.com
*通信作者:李晓岩(1982—),女,讲师,博士,主要从事花青素保健品的开发。E-mail:xyli821187@163.com
