高粱原花青素四聚体干预Streptococcus mutans Ingbritt(c)体外黏附途径及机理
2014-01-18玉佳男唐翠娥
黄 曼,蔡 欣,玉佳男,唐翠娥,刘 睿*
(华中农业大学食品科学技术学院,环境食品学教育部重点实验室,湖北 武汉 430070)
高粱原花青素四聚体干预Streptococcus mutans Ingbritt(c)体外黏附途径及机理
黄 曼,蔡 欣,玉佳男,唐翠娥,刘 睿*
(华中农业大学食品科学技术学院,环境食品学教育部重点实验室,湖北 武汉 430070)
以高纯度高粱原花青素(sorghum procyanidins,SPC)混合物和高粱原花青素四聚体(sorghum procyanidins tetramers,SPC-Ⅲ)为原料,采用荧光标记的方法比较其抑制Streptococcus mutans Ingbritt(c)黏附效果,在此基础上结合双向凝胶电泳(two-dimensional electrophoresis,2-DE)以及基质辅助激光解吸电离飞行时间质谱(matrix-associated laser dissociation/ionization time of fl ight mass spectrometry,MALDI-TOF-MS)探究其干预S. mutans Ingbritt(c)黏附的主要途径并期望阐明其作用机理。结果表明:SPC-Ⅲ比SPC混合物抑制S. mutans Ingbritt(c)体外黏附的效果更强,SPC-Ⅲ处理细菌的抑制效果强于处理获得性膜的抑制效果,是其抑制细菌黏附主要途径;类葡萄糖基转移酶(glycosyltransferases,GTFs)活性蛋白(3.10-Ⅱ)经SPC-Ⅲ作用前后的2-DE图有明显差异,差异点均为S. mutans中的不同蛋白酶类,其中A24可能是一种新蛋白。
高粱原花青素;黏附;荧光标记;双向凝胶电泳;基质辅助激光解吸电离飞行时间质谱
龋病为最常见的口腔感染性疾病,主要发生于牙齿硬组织,其患病率高、危害范围广,90%学龄儿童及成年人均不同程度受到龋病的影响。变形链球菌(Streptococcus mutans)作为主要口腔致病菌,其中多样化的毒力因子是其致龋的物质基础,尤其是葡萄糖基转移酶(glycosyltransferases,GTFs)合成的水不溶性多糖,参与介导S.mutans蔗糖依赖性黏附,是龋病发生的始动条件[1-2]。在国内外学者进行的防龋研究中,天然药物因其安全、经济,并能调理口腔菌群平衡而受到众多学者的青睐。目前认为天然植物中有效的防龋成分主要有鞣质、黄酮类化合物、木脂素和多酚类物质等。大量研究表明从多种植物(如越橘汁、乌龙茶、葡萄柚、葡萄等)中分离出的多酚物质对S. mutans中葡聚糖的形成、GTFs的活性等均表现出很强的抑制能力,从而阻碍了S. mutans的蔗糖依赖性黏附[3-6]。原花青素属于多酚类物质,它的药理活性受其单体组成、构型以及聚合度等多种因素的影响[7]。
现有研究多采用体外模拟S. mutans在唾液包被羟基磷灰石(saliva-cuated hydroxylapatite,SHA)上的黏附效果,探究天然产物成分抑制致龋菌的黏附情况[8-10],揭示其抑制能力。研究结果显示同浓度的绿茶提取物(sunphenon)和茶多酚处理细菌的黏附抑制率明显高于处理获得性膜的黏附抑制率[6],在一定程度上揭示了天然活性多酚抑制S. mutans体外黏附的主要途径。由于S. mutans是黏附于唾液在牙齿表面形成的获得性膜上,实验药物或天然活性成分有多种抑制途径,导致实验当中的可变因素多,且目前的研究多采用粗提物或混合物进行,鲜有以组成或结构明确的活性成分为原料,并探求其抑制S. mutans体外黏附主要途径的报道。
大量研究事实证明植物多酚对S. mutans的生物活性有明显影响,若要进一步探讨其抑制机理需找出S. mutans中能与多酚发生作用的受体物质,因此植物多酚对变形链球菌表面黏结素,尤其是对GTFs抑制效果的研究成为主要研究对象。通过测定GTFs经过多酚处理前后性质的改变(如GTFs酶活性)即可判定多酚抑制GTFs活性影响S. mutans的黏附[11-13],在一定程度上揭示了多酚可通过产生对GTFs的影响从而抑制S. mutans的黏附。若要进一步探求多酚与GTFs的内在作用机理,需要从分子层面上进行。随着蛋白质组学的发展,双向凝胶电泳(two-dimensional electrophoresis,2-DE)以及基质辅助激光解吸电离飞行时间质谱(matrix-associated laser dissociation/ionization time of fl ight mass spectrometry,MALDITOF-MS)也成为生物研究的主流技术,被学者们广泛用于分析细菌蛋白的差异性表达以及活性小分子与蛋白质大分子相互作用的研究。但未见将2-DE和MALDI-TOF-MS相结合运用于探究多酚和S. mutans黏结蛋白的相互作用。
因此,本研究以高纯度高粱原花青素 (sorghum procyanidins,SPC)混合物和高粱原花青素四聚体(sorghum procyanidins tetramers,SPC-Ⅲ)(ESI-MS/MS分析其为原花青素四聚体)为原料,采用荧光标记方法模拟体外黏附SHA模型筛选高活性抑制S. mutans黏附的原花青素活性成分,并分别处理细菌和获得性膜探究其干预S. mutans体外黏附的主要作用途径,在此基础上,结合2-DE和MALDI-TOF-MS分析探讨SPC-Ⅲ与S. mutans主要毒力因子GTFs的作用机理。
1 材料与方法
1.1 材料与试剂
羟基磷灰石(hydroxylapatite,HA),四川空腔医学院提供;唾液,20~25岁口腔健康的志愿者提供,低温冷冻保存;S. mutans Ingbritt(c),由武汉大学口腔医学部提供,-80 ℃甘油低温保存;高纯度SPC混合物(Porter’s法测得纯度大于98%)、SPC-Ⅲ[14](ESI-MS/MS分析其为原花青素四聚体)、类GTFs蛋白3.10-Ⅱ,实验室前期制备[7]。
荧光染料BECEF/AM 碧云天生物技术研究所。
1.2 仪器与设备
RF-5301pc荧光分光光度计 日本岛津公司;ABI4700 MALDI-TOF-TOF质谱仪 美国ABI公司。
1.3 方法
1.3.1 Tryptic Soy Broth(TSB)培养基[8]、标准菌液及活性成分溶液的制备
标准菌液:将复苏48 h后的S. mutans Ingbritt(c)接种于TSB液体培养基中,37 ℃条件下厌氧培养18~24 h,涂片检查为纯培养后,离心(3000 ×g,10 min)收集菌体,磷酸盐缓冲液洗涤2次,重新悬浮于磷酸盐缓冲液中,调节菌浓度至OD600nm=1.10,备用。
活性成分溶液:将NaF、SPC混合物和SPC-Ⅲ用0.01 mol磷酸盐缓冲液配制成下列质量浓度梯度:7.812 5、15.625、31.25、62.5、125.0、250.0、500.0 μg/mL,备用。
1.3.2 活性成分分别处理S. mutans Ingbritt(c)或获得性膜干预其在SHA上黏附的效果
实验步骤分别见图1、2,每组4份,将磷酸盐缓冲液代替药液的孔作为阴性对照组,测得的荧光值作为总荧光值。采用荧光分光光度计测定荧光强度(激发波长488 nm,发射波长535 nm)。按下式计算细菌黏附抑制率。
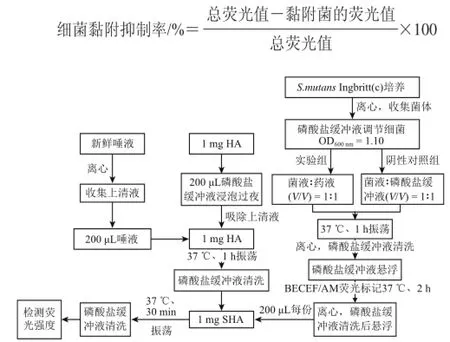
图1 活性成分处理S. mutans Ingbritt(c)后对其在SHA黏附的影响研究流程图Fig.1 Flow chart of the effect of bioactive constituent treatments on the S. mutans Ingbritt (c) adhesion to SHA

图2 活性成分处理获得性膜对S. mutans Ingbritt(c)在SHA上黏附的影响研究流程图Fig.2 Flow chart for studying the effect of bioactive constituents-treated salivary acquired pellicle on S. mutans Ingbritt (c) adhesion to SHA
1.3.3 SPC-Ⅲ与类GTFs活性蛋白3.10-Ⅱ的双向电泳分析
准确配制质量浓度为30 mg/mL的3.10-Ⅱ蛋白溶液和质量浓度为5 mg/mL的SPC-Ⅲ溶液。吸取1 mL 3.10-Ⅱ蛋白溶液冻干得到反应前蛋白粉末。再分别吸取1 mL 3.10-Ⅱ蛋白溶液和1 mL SPC-Ⅲ溶液混合,反应5 min后,4℃、10 000×g离心10 min,将上清液冻干,得到反应后蛋白粉末。将得到的两个蛋白粉末进行双向电泳分析。
1.3.4 SPC-Ⅲ与类GTFs活性蛋白3.10-Ⅱ反应前后差异点的MALDI-TOF-MS分析
凝胶显色后经扫描和图像分析,比较与SPC-Ⅲ反应前后的3.10-Ⅱ蛋白的2-DE胶图,得到差异蛋白点。将差异点进行顺序编号,利用美国ABI4700 MALDI-TOF-MS质谱仪进行分析并输入数据库检索。
1.4 数据处理
采用SPSS11.5统计软件,选用单因素方差分析的Dunnett双侧检验,比较各实验组与对照组之间荧光强度的统计学差异。
2 结果与分析
2.1 活性成分分别处理S. mutans Ingbritt(c)或获得性膜干预其在SHA上黏附的效果
设置NaF为阳性对照,SPC混合物和SPC-Ⅲ 3种活性成分先处理S. mutans Ingbritt(c),菌被荧光标记后再黏附于形成了获得性膜的SHA上,通过检测荧光强度计算其黏附抑制率。
由表1可知,随着活性成分质量浓度的升高,荧光强度逐渐减弱,随着质量浓度的上升,≥62.5 μg/mL时,各活性成分的每个质量浓度与对照组比较均有极显著差异(P<0.01),并且SPC-Ⅲ的荧光强度明显最低,提示S. mutans Ingbritt(c)经过SPC-Ⅲ处理后在SHA上黏附最少。由图3可知,随着3种活性成分质量浓度的升高,它们对S. mutans Ingbritt(c)在SHA上的黏附抑制率呈上升趋势;最高质量浓度时,SPC-Ⅲ的抑制率为67.76%明显高于另外两种成分综上所述,可推测SPC-Ⅲ处理S. mutans Ingbritt(c)后干预其在SHA上的黏附能力最强。

表1 不同活性成分处理S. mutans Ingbritt(c)干预其在SHA上黏附的荧光强度Table 1 Fluorescence intensity of S. mutans Ingbritt (c) adhesion to SHA during intervention with different bioactive constituents

图3 不同活性成分处理S. mutans Ingbritt(c)干预其在SHA上黏附抑制率变化Fig.3 Inhibitory change of S. mutans Ingbritt (c) adhesion to SHA during intervention with different bioactive constituents treatments

表2 不同活性成分处理获得性膜后干预S. mutans Ingbritt(c)在SHA上黏附的荧光强度Table 2 Fluorescence intensity of S. mutans Ingbritt (c) adhesion to SHA during intervention with salivary acquired pellicle treated by different bioactive constituents
由表2可知,当3种活性成分处理获得性膜时,随着质量浓度的升高,荧光强度逐渐减弱,SPC-Ⅲ在较高的3个质量浓度(>62.5 μg/mL)的荧光强度与对照组比较有极显著性差异(P<0.01),阳性对照NaF只有在质量浓度125.0 μg/mL处的荧光强度与对照组比较才有显著性差异(P<0.05)。由图4可知,随着3种成分质量浓度升高,对S. mutans Ingbritt(c)在SHA上的黏附抑制率呈先上升较快后平缓的趋势。最高质量浓度时,3种成分的抑制率均低于35%,但是SPC-Ⅲ对菌在SHA表面黏附的抑制率仍然表现最高。综上所述,可推测活性成分处理获得性膜后干预S. mutans Ingbritt(c)在SHA上的黏附效果较弱,但SPC-Ⅲ抑制黏附能力仍然比其他活性成分效果明显。

图4 不同活性成分处理获得性膜干预S. mutans Ingbritt(c)在SHA上黏附抑制率变化Fig.4 Inhibitory change of S. mutans Ingbritt (c) adhesion to SHA during intervention with salivary acquired pellicle treated by different bioactive constituents
根据上述结果有如下推测:活性成分处理S. mutans Ingbritt(c)比处理获得性膜干预菌在SHA上黏附的抑制能力强,且SPC-Ⅲ的抑制能力表现为最强。此结果与Otake等[15]和肖悦等[20]研究结果相似,两位学者同样采用唾液包被HA形成实验性膜的体外模式,研究结果表明,同质量浓度的实验活性成分处理致龋菌后对其黏附抑制率高于先处理获得性膜后对菌黏附抑制率。
以上结果显示活性物质处理龋齿菌抑制其黏附的效果明显好于处理获得性膜的效果。这可能是因为S. mutans的表面蛋白(黏附受体)结构中有一段富含脯氨酸并跨越细胞壁的肽段[16],多酚物质能与富含脯氨酸的蛋白形成稳定复杂的化合物,从而干扰细胞表面受体参与黏结。以前的实验多采用含有单宁等多种多酚类物质的粗提物进行实验,无法明确产生作用的具体成分[15],而本实验采用纯度高且组成成分明确的四聚体进行检测,最大程度降低了干扰现象。在唾液包被羟磷灰石的体外黏附模式中,生物膜对抗菌药物的耐受性,其中细菌的组成以及对环境的适应性等在其形成过程的不同阶段均会发生变化[17]。与浮游细菌相比,牙菌斑生物膜对于药物的敏感性因其特殊的结构和生理特性而降低[18]。研究表明茶多酚可改变获得性膜表面的性质,从而减少细菌对茶多酚处理过的SHA的黏附:首先,茶多酚能与唾液获得性膜中的富脯蛋白形成稳定的化合物,改变其构型,抑制其活性,阻碍细菌与获得性膜上的受体结合,减少细菌的黏附。其次,由于富脯蛋白和唾液淀粉酶是S. mutans在唾液获得性膜中的受体,多种茶多酚均能抑制唾液淀粉酶活性[19],由此推测,本研究中S. mutans Ingbritt(c)和获得性膜经3种活性成分处理后,其中某些具有黏附功能的因子减少或是构象、功能发生改变,致使细菌在SHA上黏附的减少。当成分质量浓度相同时,两种处理方式导致黏附抑制率出现差别,处理细菌的黏附抑制率高于处理获得性膜的黏附抑制率,可推测活性成分作用于细菌是抑制致龋菌黏附的主要作用途径。
2.2 2-DE和MALDI-TOF-MS分析与SPC-Ⅲ作用前后的类GTFs活性蛋白3.10-Ⅱ

图5 类GTFs活性蛋白3.10-Ⅱ的双向电泳图Fig.5 2-DE of 3.10-Ⅱ

图6 SPC-Ⅲ处理类GTFs活性蛋白3.10-Ⅱ的双向电泳图Fig.6 2-DE of 3.10-Ⅱ treated by SPC-Ⅲ
实验前期分离得到S. mutans Ingbritt(c)中的主要黏结蛋白——GTFs粗提物,在此基础上通过凝胶进一步纯化和分离获得类GTFs活性蛋白3.10-Ⅱ,将其作为研究目标蛋白。采用2-DE对作用前后的3.10-Ⅱ蛋白进行分析,以便更直观深入地探究两者作用前后3.10-Ⅱ蛋白的变化。双向电泳图分别见图5、6。经软件分析后将与SPC-Ⅲ作用前后灰度值明显降低的点分别编为A01~A51。其中灰度值最大的6个点分别为:A02、A08、A24、A26、A41和A51,证明它们是类GTFs活性蛋白3.10-Ⅱ与SPC-Ⅲ作用前后差异最大的点。

表3 双向电泳A24和A51蛋白点通过MALDI-TOF-MS分析和数据库检索得到的蛋白成分Table 3 Protein composition of A24 and A51 electrophoresis spot in two-dimensional electrophoresis
经数据库检索,选取点的MALDI-TOF-MS分析结果显示,以上6个点的蛋白均为S. mutans家族中所含有的具有不同功能的蛋白酶类。A24和A51蛋白点通过MALDITOF-MS分析和数据库检索得到的蛋白成分信息如表3所示。A51为二甲基腺苷转移酶。A24蛋白点信息未收录在S. mutans细菌库中,但在整个细菌库中检索出,为外膜蛋白1,是一种孔蛋白,主要存在于细胞膜上,可推测该蛋白是一种同源蛋白存在于多数菌中,也可能是以前未在S. mutans中检测出的一种新蛋白。
3 结 论
以NaF为阳性对照,荧光检测SPC混合物、SPC-Ⅲ分别处理S. mutans Ingbritt(c)和获得性膜干预其在SHA上的黏附,结果表明:3种研究材料均表现出对S. mutans Ingbritt(c)黏附于SHA的抑制作用,存在明显的浓度效应关系,其中SPC-Ⅲ的抑制效果明显优于NaF和SPC混合物;并且其处理细菌是干预体外黏附的主要途径。
从GTFs粗提物中分离得到的类GTFs活性蛋白(3.10-Ⅱ)与SPC-Ⅲ作用前后的2-DE图显示有多个蛋白点均发生不同程度的颜色变化,灰度值变化相对大的蛋白点经MALDI-TOF-MS分析后,检索得到所有点均为变形链球菌家族中的不同蛋白酶类,其中A24为外膜蛋白,在全细菌库中检索出,推测该蛋白可能是一种同源蛋白或是以前未在变形链球菌中检测出的一种新蛋白。
[1] NAKAI M, OKAHASHI N, OHTA H, et al. Saliva-binding region of Streptococcus mutans surface protein antigen[J]. Infection and Immunity, 1993, 61(10): 4344-4349.
[2] TSUMORI H, KURAMITSU H. The role of the Streptococcus mutans glucosyltransferases in the sucrose-dependent attachment to smooth surfaces: essential role of the GtfC enzyme[J]. Oral Microbiology and Immunology, 1997, 12(5): 274-280.
[3] THIMOTHE J, BONSI I A, PADILLA-ZAKOUR O I, et al. Chemical characterization of red wine grape (Vitis vinifera and Vitis interspecific hybrids) and pomace phenolic extracts and their biological activity against Streptococcus mutans[J]. Journal of Agricultural and Food Chemistry, 2007, 55(25): 10200-10207.
[4] 王进. 绿茶中EGCG提取纯化方法及其与牛血清蛋白相互作用的研究[D]. 泰安: 山东农业大学, 2009.
[5] GREGOIRE S, SINGH A P, VORSA N, et al. In uence of cranberry phenolics on glucan synthesis by glucosyltransferases and Streptococcus mutans acidogenicity[J]. Journal of Applied Microbiology, 2007, 103(5): 1960-1968.
[6] OTAKE S, MAKIMURA M, KUROKI T, et al. Anticaries effect of polyphenolic compounds from Japanese green tea[J]. Caries Research, 1991, 25(6): 438-443.
[7] 黄曼, 徐丽嫚, 陈静, 等. 响应面法优化高粱外种皮中原花青素的超声波辅助提取工艺[J]. 食品科学, 2012, 33(24): 26-30.
[8] 徐丽嫚. 高粱原花青素干预龋齿的作用机制[D]. 武汉: 华中农业大学, 2012.
[9] 闫文娟. C-di-GMP信号通路在变形链球菌致龋过程中作用的初步研究[D]. 西安: 第四军医大学, 2009.
[10] 熊炎斌, 李德玲, 刘持平, 等. 几种植物药对变形链球菌生长、产酸、黏附影响的体外实验[J]. 中华口腔医学杂志, 1997(9): 261-267.
[11] HIDEAKI T, SHIMAMURA A, MUKASA H. Comparative study of Streptococcus mutans extracellular glycosyltransferases by isoelectric focusing[J]. Journal of General Microbiology, 1983, 129(10): 3261-3269.
[12] TAKUSHI F, TOSHIHIKO K, TOSIKI N, et al. Purification and characterization of glucosyltransferases from Streptococcus mutans 6715[J]. Journal of General Microbiology, 1985, 131(2): 285-293.
[13] MUKASA H, TSUMORI H, SHIMAMURA A. Isolation and characterization of an extracellular glucosyltransferase synthesizing insoluble glucan from Streptococcus mutans serotypec[J]. Infection and Immunity, 1985, 49(3): 790-796.
[14] 徐丽嫚, 黄曼, 涂世, 等. 高粱原花青素的 ESI-MS 分析及其低聚体的RP-HPLC-MS/MS 法分离鉴定[J]. 食品科学, 2011, 32(20): 221-225.
[15] OTAKE S, MAKIMURA M, KUROKI T, et al. Anticaries effect of polyphenolic compounds from Japanese green tea[J]. Caries Research, 1991, 25(6): 438-443.
[16] BRADY L J, van TILBURG M L, ALFORD C E, et al. Monoclonal antibody-mediated modulation of the humoral immune response against mucosally applied Streptococcus mutans[J]. Infection and Immunity, 2000, 68(4): 1796-1805.
[17] TAKAHASHI N, NYVAD B. The role of bacteria in the caries process: ecological perspectives[J]. Journal of Dental Research, 2011, 90(3): 294-303.
[18] TEN CATE J M. Biofilms, a new approach to the microbiology of dental plaque[J]. Odontology, 2006, 94: 1-9.
[19] 李鸣宇, 刘正, 朱彩莲. 人工合成变形链球菌表面蛋白多肽体外抗黏附实验[J]. 口腔医学纵横杂志, 2001, 17(3): 176-178.
[20] 肖悦, 刘天佳, 黄正蔚, 等. 茶多酚对口腔细菌致龋力影响的实验研究[J]. 广东牙病防治, 2002, 10(1): 426-429.
in vitro Adhesion Pathways and Mechanisms of Streptococcus mutans Ingbritt (c) during Intervention with Proanthocyanidins Tetramers
HUANG Man, CAI Xin, YU Jia-nan, TANG Cui-e, LIU Rui*
(Key Laboratory of Environment Correlative Dietology, Ministry of Education, College of Food Science and Technology, Huazhong Agricultural University, Wuhan 430070, China)
In this study, high purity proanthocyanidins (SPC) and proanthocyanidins tetramers (SPC-III) from sorghum episperm were compared for their inhibitory effects on the adhesion of Streptococcus mutans Ingbritt (c) through fluorescence labeling. By combining two-dimensional electrophoresis (2-DE) and matrix-associated laser dissociation/ ionization time of flight mass spectrometry (MALDI-TOF-MS), the major intervening pathways were investigated, and the underlying mechanisms were explored. The results showed that SPC-III (tetramers) had better inhibitory effect than SPC and had stronger inhibitory effect than SPC-III-treated salivary acquired pellicle on the adhesion of S. mutans Ingbritt (c), which was the major pathway to inhibit bacterial adhesion. Two-dimensional electrophoretograms of 3.10-II treated by SPC-III or not showed significant differences. All different points were different proteases derived from S. mutans, in which A24 was suggested to be a new protein that has never been detected in S. mutans previously.
sorghum proanthocyanidins; adhesion; fluorescence labeling; two-dimensional electrophoresis; matrixassociated laser dissociation/ionization time of fl ight mass spectrometry
TS201.9
A
1002-6630(2014)03-0001-05
10.7506/spkx1002-6630-201403001
2013-07-19
国家自然科学基金项目(31271939)
黄曼(1988—),女,硕士研究生,研究方向为天然产物化学与功能食品。E-mail:huangman_880103@163.com
*通信作者:刘睿(1969—),男,副教授,博士,研究方向为天然产物化学与功能食品。E-mail:liurui@mail.hzau.edu.cn
