引种鲁梅克斯长叶-20的生殖特征
2014-01-16常缨安玉婷李艳徐永清李凤兰胡宝忠
常缨,安玉婷,李艳,徐永清,李凤兰,胡宝忠
(东北农业大学生命科学学院,哈尔滨 150030)
引种鲁梅克斯长叶-20的生殖特征
常缨,安玉婷,李艳,徐永清,李凤兰,胡宝忠
(东北农业大学生命科学学院,哈尔滨 150030)
运用石蜡切片技术、光学显微镜和扫描电子显微镜技术对鲁梅克斯长叶-20花芽形态分化、大小孢子发生与雌雄配子体发育以及胚后发育等生殖过程进行研究。结果表明,鲁梅克斯长叶-20花芽分化分为花序原基分化期、小花原基分化初期、萼片原基分化期、花瓣原基分化期、雄蕊原基形成期和雌蕊原基形成期;小孢子母细胞经减数分裂形成四分体,属四面体型,成熟花粉粒以2-细胞型为主,兼有3-细胞型,具有3条或4条萌发沟;雌蕊发育过程中双珠被的倒生胚珠、厚珠心发育形成核型胚乳,大孢子母细胞减数分裂后形成的四分体呈线形排列,合点端的一个功能大孢子发育成单核胚囊,经三次连续的有丝分裂形成八核的蓼型胚囊;受精后初生胚乳核先于合子分裂,为胚的发育提供营养,胚经过原胚期、球形胚期、心形胚期直至发育成具有完整子叶的成熟胚。
鲁梅克斯长叶-20;花芽分化;大小孢子发生;雌雄配子体发育
鲁梅克斯长叶-20(Rumexlongleaf-20)是蓼科酸模属多年生宿根草本植物,作为新型高蛋白饲科和优良的防止水土流失、改善生态环境的地被植物引种于俄罗斯[1]。目前对鲁梅克斯品种的研究主要集中于鲁梅克斯K-1,黄光明等研究表明,在畜牧业方面,用鲁梅克斯K-1可代替部分鲜绿饲料,饲喂牛、羊、猪等畜禽[2-3]。在饲料工业方面,可加工成蛋白质丰富的饲料或饲料添加剂,降低饲料成本[4-5]。鲁梅克斯K-1也可作保健食品,弥补人体中VC摄入量较低的缺陷[6-7]。在食品工业方面,鲁梅克斯K-1可用于多种食品制备,开发高品质营养粉和营养液,生产安全无公害保健食品[8-9]。
目前国内对鲁梅克斯品种的营养成分、栽培技术、耐盐性、生物育种及用于食品和叶蛋白提取等方面已有研究。而对鲁梅克斯品种的生物学特性,尤其是其胚胎发育方面鲜见研究报道。本文以鲁梅克斯长叶-20为材料,运用石蜡切片技术、光学显微镜和扫描电子显微镜技术对鲁梅克斯长叶-20花芽形态分化、大小孢子发生与雌雄配子体发育及胚后发育等生殖过程进行研究,全面了解鲁梅克斯长叶-20的生殖生长特性,为其规模化生产和栽培实践提供理论依据。
1 材料与方法
1.1 材料
供试材料鲁梅克斯长叶-20(Rumexlongleaf-20)种子购买于俄罗斯,于2013年6月9日播种在东北农业大学阿城实验实习基地。
1.2 方法
1.2.1 花芽形态分化的试验方法
2013年7月末至2013年9月初每隔5 d取10株鲁梅克斯茎上的第一真叶叶腋处的花芽,花芽外部形态大且饱满,顶端圆钝,抱合紧密,所取材料放在有标记的小瓶中,采用70%FAA固定液固定。采用常规石蜡切片方法,切片厚度为10 μm,番红-固绿双重染色,中性胶封片,在光学显微镜装置(E-200尼康公司)下观察并照相。
1.2.2 雄蕊及雌蕊形态发育的试验方法
2013年9月初至9月末,根据花蕾发育不同时期,每隔3 d取鲁梅克斯长叶-20植株上的花蕾和刚开的完全花,将所取材料放在有标记的小瓶中,采用70%FAA固定液固定,固定一定时间后,逐级酒精脱水,二甲苯透明,熔点60~62℃的石蜡包埋,切片厚度为10 μm,番红-固绿双重染色,中性树胶封片,在光学显微镜装置下观察并照相。
1.2.3 花粉形态的观察方法
①扫描电镜法。将2013年9月中旬盛开的鲁梅克斯长叶-20花的花被片剥离,取下花药,戊二醛固定,常规扫描电子显微镜样品方法,即固定、换样和置换、干燥、粘样、镀膜后置于扫描电子显微镜(S-3400日立公司)下观察并照相。
②透射电镜法。将2013年9月中旬盛开的鲁梅克斯长叶-20花的花被片剥离,取下花药,戊二醛固定,常规透射电子显微镜样品方法,即取材、固定、脱水、浸透、包埋聚合、切片及染色等步骤,然后置于透射电子显微镜(H-7650日立公司)下观察并照相。
1.2.4 胚和胚乳发育的试验方法
2013年10月9日至2013年10月20日,根据从授粉后到种子完全成熟的不同时期,每隔3 d取样1次,将所取材料放在有标记小瓶中,采用70% FAA固定液固定,在固定液中固定一定时间后,逐级酒精脱水,二甲苯透明,熔点60~62℃的石蜡包埋,切片厚为10 μm,番红-固绿双重染色,中性树胶封片,在光学显微镜装置下观察并照相。
2 结果与分析
2.1 花芽形态分化
鲁梅克斯长叶-20的花序为穗状花序,可按花序原基和花原基各部分发生顺序将该植物的花芽分化整个过程分为:花序原基分化期、小花分化初期、萼片原基分化期、花瓣原基分化期、雄蕊原基形成期和雌蕊原基形成期等6个时期。
2.1.1 花序原基分化期
穗状花序的花芽分化顺序为下位花先分化,上位花后分化(见图版Ⅰ-1)。
2.1.2 小花分化初期
花原基及生长锥的体积增大,生长锥先端变得宽而肥大,向上隆起,呈近半球形(见图版Ⅰ-2)。
2.1.3 萼片原基分化初期
生长锥顶端开始变宽而扁平,然后边缘产生小突起,此突起即为萼片原基,并逐渐伸长(见图版Ⅰ-3),最终发育成花萼。
2.1.4 花瓣原基分化期
随着萼片原基的伸长、内弯,在其内侧基部形成新的突起即为花瓣原基,花瓣原基不断伸展(见图版Ⅰ-4,5),最终发育成花瓣。
2.1.5 雄蕊原基形成期
随着萼片原基和花瓣原基的逐渐伸展,在花瓣原基内侧相继出现的突起,即为雄蕊原基(见图版Ⅰ-6,7),顶端分生组织活跃,分化出花丝和花药。
2.1.6 雌蕊原基形成期
在芽内生长锥中心基部向上出现小突起,形成雌蕊心皮原基(见图版Ⅰ-8,9),将花芽纵切后可以看出一朵发育中的花完整的组成部分,由外到内应该依次为萼片原基、花瓣原基、雄蕊原基和雌蕊原基。

图版I 鲁梅克斯长叶-20花芽分化的过程Plate I Process of flower bud differentiation of Rumex longleaf-20
2.2 小孢子发生及雄配子体发育
鲁梅克斯长叶-20的成熟花药具有4个花粉囊,常以药隔分成2个药室,每一药室具2个花粉囊,药隔由薄壁细胞组成,药隔组织延伸至相邻的两个花粉囊之间,每个花粉囊的一半由药隔包围,药隔维管束位于中间。
2.2.1 花药壁发育
幼嫩的花药横切结构显示花药的最外层是表皮,由一层细胞组成。表皮内部是一群分裂极为活跃的分裂细胞,呈不规则排列。后来由于花药的花粉囊处的细胞分裂速度比较快,从而在每个花粉囊的表皮下分裂分化出孢原细胞。孢原细胞进一步发育是经过一次平周分裂形成初生壁细胞和初生造孢细胞。初生壁细胞以后又进行1次或数次平周分裂和多次垂周分裂,形成4层壁细胞。以后4层壁细胞继续生长,并出现分化:外面2层细胞体积增大缓慢,长圆形或纺锤形,而里面的2层,特别是最里面的一层细胞体积增大速度很快,在径向方向上逐渐扩展,原生质体很浓厚。最后,源于最外层的细胞分裂分化为药室内壁;源于第2层的细胞分裂分化为中层细胞;从第3层细胞的增殖和分化中形成中层细胞,但后来被压成扁平状,并逐渐解体消失;从第4层细胞最终分化成绒毡层。在这个时候,花药壁得到充分发育,从外到内分为表皮(1层)、药室内壁(1层)、中间层(2层)、绒毡层(1层)。因此鲁梅克斯长叶-20的花药壁发育属于基本型(见图版Ⅱ-1)。
随着小孢子母细胞减数分裂以及花粉粒的形成,花药壁发生一系列变化。观察结果表明表皮和药室内壁存在时间最长,一直到花粉囊开裂。从小孢子母细胞进行减数分裂开始,中层细胞先被挤压成扁形,后以狭长状存在,直至营养被吸收殆尽而消失。绒毡层细胞,细胞核大而明显,液泡小,细胞质不密集(见图版Ⅱ-2)。花粉粒处于单核期时,绒毡层细胞显著退化,至花粉粒发育成熟时,绒毡层细胞完全解体(见图版Ⅱ-3)。绒毡层细胞一直处在原位置,通过细胞内表面分泌各类物质为小孢子发育提供营养,因此鲁梅克斯长叶-20的绒毡层属于腺质绒毡层(见图版Ⅱ-4)。花粉成熟,两个小孢子囊在花粉囊的同一侧联通,然后在两个小孢子囊的联通处开裂,花药壁开裂只有药室内壁和表皮细胞残留(见图版Ⅱ-5)。
2.2.2 小孢子发生
造孢细胞经数次有丝分裂形成小孢子母细胞,其形态与周围花药壁的细胞有明显的差别。观察结果显示:小孢子母细胞呈大致等径的多边形,细胞体积大,细胞核大,细胞质浓,可见1个核仁,无明显液泡(见图版Ⅱ-6)。第一次减数分裂时,小孢子母细胞被胼胝质壁包围,随后进行的第二次减数分裂形成四分体,四分体时期小孢子没有明显的液泡,四分体包藏在共同的胼胝质壁中,四个小孢子也被胼胝质分开。鲁梅克斯长叶-20减数分裂的胞质分裂为同时型,四分体多数呈四面体形排列。同一药室内的小孢子母细胞在减数分裂时均处于同一时期,具有同步性;同一花药中的4个药室之间,小孢子母细胞的减数分裂不完全同步,但相差不大。
2.2.3 雄配子体发育
刚从四分体释放出来的小孢子被许多胼胝质物质包围,由于不断从周围的绒毡层吸收营养和水分而很快变圆,随后分开。该时期小孢子细胞体积大,核处于中心,核仁很显著,细胞壁逐渐加厚,为单核居中期(见图版Ⅱ-7)。随着小孢子细胞体积逐渐增大,细胞质不断液泡化最终形成体积较大的中央液泡,中央液泡挤压小孢子向花粉壁一侧靠拢,为小孢子单核靠边期(见图版Ⅱ-8)。小孢子在进一步发育中,小孢子核发生不均等有丝分裂形成两个细胞,细胞质多、细胞体积大的为营养细胞;细胞质少、细胞体积小的为生殖细胞,生殖细胞靠近花粉壁一侧,两个细胞被胼胝质壁分开。随着胼胝质壁的溶解,生殖细胞融入营养细胞的细胞质中,形成2-细胞型的花粉或3-细胞型的花粉(见图版Ⅱ-9,10)。扫描电镜下观察到散粉前花粉粒的孢粉壁上有粘稠物质存在和较多成熟花粉粒(见图版Ⅱ-11,12),未观察到花粉败育现象。
2.2.4 花粉形态
扫描电镜观察结果显示鲁梅克斯长叶-20的花粉粒较多,花粉为近球形,赤道面观为圆形,极面观为三裂圆形,大小为(17.3±0.2)μm,具3沟或4沟(见图版Ⅲ-1,2)。可以看出花粉外壁表面有3条或4条萌发沟(见图版Ⅲ-3,4),花粉表面为网纹状结构,具覆盖层和瘤状突起,突起大小不一(见图版Ⅲ-5)。随着花粉粒发育,营养物质逐渐被消耗,逐渐花粉粒出现类似沟状的结构,花粉管从靠近柱头的凹陷处萌发。
透射电子显微镜观察表明,鲁梅克斯长叶-20花粉粒较多,可看到2-细胞型花粉粒和3-细胞型花粉粒内结构,包括核物质和较多淀粉粒(见图版Ⅲ-6,7,9,10)。花粉壁结构由外层(包括覆盖层、柱状层和基层)和内层组成(见图版Ⅲ-8,11)。花药壁在成熟花粉粒时期时,此时花药壁由表皮、药室内壁组成(见图版Ⅲ-11,12)。花粉壁结构常分为外壁层和内壁层两层细胞,本文研究花粉壁形态,主要是研究外壁结构。外壁由外层细胞和内层细胞组成。外层细胞一般由3层组成,最外层为覆盖层细胞,形态发育不完全时则为半个或无覆盖层细胞花药壁;下面一层为柱状层细胞,这层细胞的形状是柱状的或棒状的;最下面一层细胞为基层细胞。

图版Ⅱ 鲁梅克斯长叶-20花药的发育PlateⅡ Anther development of Rumex longleaf-20

图版Ⅲ 鲁梅克斯长叶-20花粉的发育PlateⅢ Pollen development of Rumex longleaf-20
2.3 大孢子发生及雌配子体发育
2.3.1 雌蕊结构
鲁梅克斯长叶-20的单一雌蕊是由柱头、花柱和子房三部分组成。柱头为头状或增加裂为丝状或流苏状,有大量的柱头乳突细胞。子房上位,心皮1,1室1胚珠,一个胚珠着生于心皮腹线上,为具双珠被的倒生胚珠(见图版Ⅳ-1,2)。
2.3.2 胚珠的结构与发育
胚珠是在心皮内侧沿腹缝处的胎座上发育的,最初产生的一团突起是胚珠的珠心细胞,珠心是胚珠中极为重要的部分,以后的胚囊就是由这部分细胞发育而来。在珠心原基的内侧形成内珠被原始细胞,由于内珠被原始细胞进行平周分裂,形成内珠被;接着在内珠被基部以同样方式形成外珠被。随着胚珠珠被的不断发育,胚珠开始向一侧弯曲,形成倒生胚珠,整个胚珠倒转,内珠被形成珠孔。内珠被生长速度先于外珠被,在孢原细胞时期,在大孢子四分体时期,内珠被几乎达到珠心顶端,而外珠被此时才到达珠心的中心。胚珠基部的一部分细胞分裂分化为柄状结构,与心皮直接相连,称珠柄。珠孔与珠柄非常接近,珠被和珠心以及珠柄一起愈合的部分称为合点。
2.3.3 大孢子发生
大孢子发生开始于珠心表皮下的一个特殊细胞即为孢原细胞,与其他珠心细胞相比,体积较大,细胞质浓厚,具显著的细胞核,形状不规则。
大孢子母细胞是由造孢细胞膨大后发育而来的,大孢子母细胞呈椭圆形,随着大孢子母细胞的发育,细胞核明显大于周围的细胞,大孢子母细胞染色质浓缩后,细胞体积增大,细胞核由珠孔端移至中央开始进行减数分裂。大孢子母细胞进行减数分裂分别形成二分体和直线型排列的四分体(见图版Ⅳ-3)。在此时,珠被内表皮细胞开始逐步在径向方向延伸,细胞质浓厚,四分体形成后远离合点端的3个大孢子停止生长和分化并解体,而合点端的大孢子随后发育成功能大孢子。
2.3.4 雌配子体发育
周围组织的营养和水分为功能大孢子所吸收,体积增大为单核胚囊,单核胚囊细胞核较大,位于胚囊中央(见图版Ⅳ-4)。单核胚囊首先有丝分裂形成二核胚囊,其次是三次有丝分裂形成八核胚囊(见图版Ⅳ-5,6)。跟着核分裂,胚囊显著扩大,中央被大液泡占据。八核胚囊进一步发育,最开始从合点端和珠孔端移向胚囊中央的两个核构成中央细胞(见图版Ⅳ-7),珠孔端分化出两个助细胞、一个卵细胞(见图版Ⅳ-8),合点端的三个核则分化成三个反足细胞(见图版Ⅳ-9),各细胞核的核仁都很明显。成熟胚囊含有7个细胞8个核。从上述发育过程来看,胚囊发育属于典型的蓼型胚囊。
成熟胚囊的结构:卵细胞是雌配子,是胚囊中极为重要的成员,与雄配子形成新一代孢子体。成熟的卵细胞是梨形,位于合点端,有一个大的中央液泡,卵细胞极性不明显(见图版Ⅳ-8)。两个助细胞并列在一起,极性明显,细胞质聚集于珠孔端,核位于珠孔端,液泡占据合点端,极性恰与卵细胞相反。成熟雌配子体中的2个助细胞与卵细胞在空间上呈三角形排列构成卵器。助细胞通常生命期较短,其中一个进入花粉管之前就已退化,另一个在受精后不久也退化。中央细胞高度液泡化,中间有两个相互靠近的极核,极核较大,与其他细胞相比,占据胚囊最大空间(见图版Ⅳ-7)。
2.4 胚和胚乳的发育
受精后珠被发育成种皮时发生变化,主要是珠孔瑞的内珠被细胞变形,内珠被前期原生质体开始解体逐渐被吸收,外珠被发育成种皮。外珠被表皮细胞沿轴向伸长,细胞质逐渐消失,其外壁后期形成一层很厚的角质层,约由1~2层细胞组成。最后外层细胞有膨大变化,起保护作用。
2.4.1 胚的发育
种子里的胚是由卵经过受精后的合子发育来的,合子是胚的第一个细胞。合子第一次有丝分裂形成两个核,将来发育成基细胞和顶细胞,靠近珠孔端的细胞,称为基细胞;远离珠孔端的细胞,称为顶细胞。顶细胞将随后发育为胚,而基细胞只具有提供营养物质的功能,不具备发育为胚的特性,以后发育成为胚柄。基细胞和顶细胞之间的联系由两者之间的胞间连丝联通。然后2个细胞各自分别进行一次分裂,成为4个细胞的原胚。之后,4个细胞又各自不断地从横向或纵向进行分裂,胚的总体积逐渐增大。分裂成16~32细胞的胚胎时期,胚呈现棍棒状,上面部分经分裂分化发育成为胚体的前身,下面细长的部分,分裂分化为胚柄(见图版Ⅳ-10)。
2.4.2 胚乳的发育
胚乳是被子植物种子贮藏的营养结构,由2个极核和精子融合形成受精极核,不断发育形成初生胚乳核,是三个核融合的产物。极核受精,无休眠期,直接就发育为胚乳。胚乳的发育一般有核型、细胞型和沼生目型三种,而鲁梅克斯长叶-20的胚乳发育主要以核型为主。
被子植物核型胚乳的发育:2个极核和精子融合的受精核发育中只有核的分裂而没有形成细胞壁,细胞核在同一细胞质中仍是游离状态,所以这段时期称为游离核的生成期。随着游离核数的增加,在中央液泡出现的条件下,核和原生质慢慢地被挤向胚囊的两极边缘,主要分布在珠孔端和合点端。核的分裂主要是以有丝分裂为主,在少数情况下也会出现无丝分裂,特别是在远离珠孔端分布的核。胚乳游离核分裂到一定程度,它直接进入细胞形成时期,此时在各个游离的细胞核之间形成细胞壁,从而完成细胞质分裂形成胚乳细胞,这时整个胚乳细胞组织称为胚乳。在多细胞原胚形成过程中,种子内大量空间被胚乳占据。胚乳细胞不断分裂增生,包围整个棒状结构(见图版Ⅳ-11)。
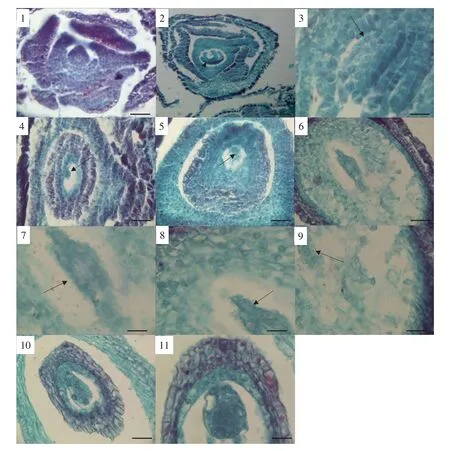
图版Ⅳ 鲁梅克斯长叶-20胚胎发育PlateⅣ Embryonic development of Rumex longleaf-20
3 讨论
3.1 鲁梅克斯长叶-20花芽分化与温度的关系
花芽分化不仅与各种植物的活动特征变化有关;还与外界环境条件和农业技术措施有密切关系,其中温度对花芽分化影响很大,在高温条件下,能促进花芽分化完成,这与本试验中在8月份较高的温度条件下完成花芽分化整个过程一致。总之,掌握外界温度变化规律,且在恰当的农业技术条件下,满足花芽分化对内部和外部条件的需求,使其完成充足的花芽分化过程,对提高鲁梅克斯长叶-20的产量具有重要意义。
3.2 温度变化对鲁梅克斯长叶-20胚胎发育的影响
本试验中,在2013年9月初鲁梅克斯长叶-20开始进入胚胎发育阶段,包括大小孢子发生、雌雄配子体发育、传粉受精、胚和胚乳的发育,接着在10月中旬进入枯萎期,整个胚胎发育时期为1个半月。而这两个月的温度分别为16.0和5.1℃,相对比俄罗斯种植地区同期温度为11.5和6℃,哈尔滨地区9月份温度较高,有利于其胚胎发育。两地温度相差不大,且哈尔滨地区较高的温度更适宜引种鲁梅克斯长叶-20的种植和生长。观察并计算其结籽期的结籽数量,每一穗状花序上的花经发育都能结籽,适宜的温度可提高鲁梅克斯产量。
由于鲁梅克斯长叶-20是第一年在哈尔滨地区种植,虽然结籽率较高,但是抽茎的植株棵树较少,为了解植株未抽茎原因,在10月初植株未枯萎之前,将部分植株移植到温度高于室外的大棚内,但仍未抽茎,究其原因有可能是地域差异、光照条件及种子特性等问题,尚需进一步研究。
3.3 鲁梅克斯长叶-20雌雄蕊发育与结实的关系
本试验中鲁梅克斯长叶-20是雌雄异株植物,雌株和雄株发育到一定阶段同时成熟,同熟可以帮助植物完成正常授粉。植株在异花授粉之后经过一段时间会结籽,其中抽茎的植株中雌株几乎完全结籽,结籽率良高,说明植株的雌蕊和雄蕊发育良好,不会影响该植物的结籽率。抽茎的植株数太少,植株未抽茎就不会从营养生长阶段进入生殖生长阶段,整个植株就不会完成生殖发育过程,也就不会开花结籽,导致鲁梅克斯长叶-20整体结籽量下降。是否可以推断直接影响植株抽茎的因素也会间接影响植株的结籽率,尚待深入验证。
4 结论
本试验以鲁梅克斯长叶-20为材料,运用石蜡切片技术、光学显微镜和扫描电子显微镜等技术对鲁梅克斯长叶-20的花芽形态分化、大小孢子发生与雌雄配子体发育以及胚后发育等生殖生物学过程进行研究,全面了解鲁梅克斯长叶-20的生殖生长特性。试验结果如下:
①鲁梅克斯长叶-20的花序为穗状花序。7月末进入花芽分化时期,到9月初已初具花的形态。按花序和花原基各部分发生的顺序将该植物花芽分化的整个过程分为:花序原基分化期、花分化初期、萼片原基分化初期、花瓣原基分化期、雄蕊原基形成期和雌蕊原基形成期等6个时期,从开始分化到雌蕊原基形成,大概需1个月时间,且穗状花序分化顺序为下位花先分化,上位花后分化。
②在9月末时,鲁梅克斯长叶-20开始其雌雄配子体的发育。成熟花药有4个花粉囊,纵向开裂,花药横切面呈蝶形。花药壁发育为基本型,绒毡层为腺质绒毡层。小孢子在四分体中的排列方式多数为四面体形,少数为十字交叉形。4个小孢子分裂同步,但在同一药室中减数分裂步骤表现为不一定同步。成熟花粉粒为2-细胞型或3-细胞型,以2-细胞型花粉粒为主,圆球形,外壁表面呈网纹状,具瘤状突起,具有3条或4条萌发沟。
③雌蕊胚珠原基珠心表皮下的孢原细胞经过一次分裂形成初生壁细胞和初生造孢细胞,随后初生壁细胞继续分裂形成4层壁细胞,造孢细胞膨大发育成大孢子母细胞。大孢子母细胞减数分裂后形成的四分体呈线形排列,合点端的一个功能大孢子发育成单核胚囊,经三次连续的有丝分裂形成八核的蓼型胚囊,反足细胞液泡化,中央细胞内两个极核核大而明显。子房上位,具双珠被的倒生胚珠,厚珠心。
④受精的合子有短暂的休眠期,初生胚乳核的分裂在合子分裂之前,为胚的发育供给营养成分。胚乳发育类型为核型。
[1]安继忠.高产耐盐碱的新型高蛋白牧草─杂交酸模[J].饲料工业,1999,20(11):26.
[2]黄光明.蛋白质最高的牧草─鲁梅克斯K-1[J].中国农学通报,1999,15(40):73.
[3]石定燧,熊军功.高产优质饲料作物——鲁梅克斯K-1[J].新疆农业大学学报,1996,19(4):54-56.
[4]李翼.新型高蛋白植物鲁梅克斯K-1及其应用前景初析[J].中国食物与营养,1999(2):23-24.
[5]李守德.鲁梅克斯K-1杂交酸模的品种审定及特性[J].中国食品与营养,1999(增):25.
[6]薛颖,宋曙辉.鲁梅克斯K-1杂交酸模营养成分分析报告[J].中国食物与营养,1999(3):26-27.
[7]熊军功.鲁梅克斯绿色产业的形成与发展[J].中国食物与营养,1999(2):23-24,37.
[8]李水龙.鲁梅克斯绿色产业─中国21世纪巨龙产业[J].中国食物与营养,1999(1):14-16.
[9]张磊.大有开发前途的一种植物资源─杂种酸模[J].天津农林科技,1999(1):28-29.
Reproductive characteristics ofRumexlongleaf-20
CHANG Ying,AN Yuting,LI Yan,XU Yongqing,LI Fenglan,HU Baozhong
(School of Life Sciences,Northeast Agricultural University,Harbin 150030,China)
Using the paraffin section technique,light microscope and scanning electron microscope (sem)technique to studyRumexflower bud morphological differentiation,megasporogenesis and microsporogenesis,male and female gametophyte development,and post-embryonic development.The results showed that the whole flower bud differentiation process could be divided into inflorescence primordia differentiation stage,initial differentiation stage,initial sepal primordium differentiation stage,petal primordium differentiation stage,stamen primordium formation differentiation stage,and pistil primordium formation differentiation stage;the tetrad formed by meiosis of microspore mother cell belonged to tetrahedral shape type;with three or four germination grooves,most mature pollen grains were 2-cell type,few were 3-cell type;the tetrad from meiosis of megaspore mother cell was linear;the chalazal megaspore developed into mononuclear embryo sac which further developed into eight nuclear embryo sac of Polygonum type by three successive mitotic;after fertilization,the primary endosperm nucleus division preceded zygote division to provide nutrition for embryo development;after proembryo,globular embryo,and heart-shaped embryo stage,a mature embryo with complete cotyledons was formed.
Rumex;flower bud differentiation;megasporogenesis and microsporogenesis;male and female gametophyte development
S812;Q944.4
A
1005-9369(2014)11-0043-09
2014-05-16
中俄合作项目(2013DFR30270)
常缨(1970-),女,教授,博士,博士生导师,研究方向为植物资源学与植物分子生物学。E-mail:yingchang1970@yahoo.com
时间2014-11-21 16:33:52[URL]http://www.cnki.net/kcms/detail/23.1391.S.20141121.1633.003.html
常缨,安玉婷,李艳,等.引种鲁梅克斯长叶-20的生殖特征[J].东北农业大学学报,2014,45(11):43-51.
Chang Ying,An Yuting,Li Yan,et al.Reproductive characteristics ofRumexlongleaf-20[J].Journal of Northeast Agricultural University,2014,45(11):43-51.(in Chinese with English abstract)
