天然草地与改良草地地上生物量空间分布及其群落结构特征
2014-01-07刘书娟高宁宁汪丽平王文颖程积民
刘书娟,陈 俊*,高宁宁,汪丽平,王文颖,程积民
(1.西北农林科技大学 动 物科技学院,陕西 杨 凌712100;2.中国科学院 水 利部水土保持研究所,陕西 杨 凌712100)
草地是世界上分布最广的植被类型之一,是陆地生态系统的重要组成部分,草地生态系统在调节气候、涵养水源、保持水土等方面具有十分重要的作用[1-2]。草地生物量是衡量草地生产力高低的重要参数,也是描述第一性生产的重要概念。草地植被生物量的空间分布格局与异质化程度,可以反映草地植物群落格局的变化,也可以综合体现群落结构和功能的变化[3-6]。因此研究草地生物量的空间分布规律,对草地改良和实际生产具有重要的学术价值。
空间异质性是指生态学变量在空间上的不均匀性和复杂性,是草地生态系统的重要特征[7-8]。经研究发现,人为干扰、土壤养分的不均一性以及植物自身的生物学特性等均可以影响植物群落的空间异质性,进而反映出草地群落结构的发展变化[9-12]。植物群落的空间异质性对生物量和生态系统中物质循环与能量流动存在着一定的影响,草地植被生物量可以反映该草地的生产情况[13-15],因此,探明生物量与空间异质性的关系是十分必要的。本研究对彬县退化天然草地和补播苜蓿的改良草地进行调查,运用γ分布模型的数理解析方法,探讨草地植物空间分布的异质性规律及其与生物量之间的相关性,从空间异质性的角度揭示退化天然草地和补播苜蓿的改良草地植物群落的动态变化,探讨草地改良对群落生物量、物种数、物种多样性以及空间异质性的影响程度,为退化天然草地生产量的提高及其可持续利用提供理论依据。
1 材料与方法
1.1 调查地概况
调查地位于陕西省彬县永乐镇高辉村(E108°06′13″;N35°16′26″;海拔1 096 m)。该地区退化的天然草地群落主要由黄背草、赖草、本氏针茅,铁杆蒿、茭蒿,多裂委陵菜,草木樨状黄芪、达乌里胡枝子等植物组成。彬县属渭北旱塬塬梁沟壑区,全县分为南北两塬一道川,泾河自西而东斜贯其中,境内流长104 km[16],总面积1183 km2,平均海拔1 108 m,处于半湿润和半干旱过渡气候带,对气候的变化较为敏感,是一个生态脆弱区[17]。年平均日照时数2298.8 h,年平均气温9.7℃,昼夜温差11.7℃,1月份平均气温-4.8℃,7月份平均气温22.3℃。年均降水量579 mm,且57%的降水集中在7~9月,无霜期180 d,属典型的大陆性暖温带半干旱气候。土壤类型为黑垆土,耕层质地较轻,疏松易耕,适耕期长,富含腐殖质,供肥性能好。
1.2 调查时间和方法
野外调查于2012年6月1日至10日在彬县永乐镇高辉村的退化天然草地和补播苜蓿的改良草地内进行。首先分别在退化的天然草地和补播苜蓿的改良草地内选择有代表性的地段各拉一条50 m长的样线,在每条样线上依次摆放100个50 cm×50 cm的样方;然后将样方内的所有植物齐地面刈割,带回实验室后按种分类装入纸袋;最后在65℃恒温干燥箱干燥48 h至恒重,将干燥后的植物样本称重,得到样方内所有植物种的地上生物量数据。
1.3 数据解析方法
1.3.1 γ分布模型 如果用xi表示单位面积内的生物量,则生物量的空间分布符合γ分布模型,γ分布模型可用公式(1)表示[21-22]。

式中:μ表示单位面积的平均生物量,α为生物量频度分布形状的参数。μ和α可按公式(2)表示。

式中:N为调查的总样方数(本试验中N=100),v是N个生物量数据的方差,xi为各个样方内植物种i的生物量。
1.3.2 空间异质性指数 组成群落的各植物种群的空间异质性指数值(Ii)可以用公式(3)表示[21]。

当α=1时,μ2=v,Ii=1,说明群落内的植物种群在空间上处于随机分布状态;式(1)则变为下列的指数函数形式。

当α→∞时,Ii→0,即v趋近于0,此时群落内的植物种群在草地上均匀分布;当α→0时,Ii→∞,说明组成群落的植物种群具有较强的空间异质性,且Ii值越大,异质性程度越强。
群落的空间异质性程度是在综合考虑组成群落的每种植物种群生物量大小与其异质性程度的基础上得出的,用公式(5)表示。

1.3.3 物种多样性指数 采用Shannon-Wiener的H'指数,用公式(6)表示。


1.3.4 数据处理 采用SPSS16.0统计软件对数据进行方差分析,使用Microsoft Excel 2003软件绘图。
2 结果与分析
2.1 草地主要植物种群对γ分布模型的吻合性
由表1可知,31种植物实测生物量的频度分布与γ分布模型的理论频度之间差异都不显著(P>0.05),即它们均与γ分布模型相吻合,说明这2种草地中的主要组成植物种生物量的分布符合γ分布模型,故草地生物量的空间分布格局可以利用γ分布模型进行解析。由图1知,4种代表性植物紫花苜蓿、茭蒿、本氏针茅、达乌里胡枝子的生物量频度分布,与γ分布模型具有很好的吻合性。

表1 γ分布模型的吻合性检验结果Table 1 Fitness of the gamma distribution by chi-square test
2.2 天然草地与改良草地的群落特征
由表2可知,改良草地的总生物量高于然草地,但总物种数和群落的空间异质性指数均低于天然草地,改良草地的平均生物量极显著的高于天然草地(P<0.001),但平均物种数与平均物种多样性指数均极显著的低于天然草地(P<0.001)。
2.3 空间异质性指数与生物量之间的关系
由图2可知,不然草地与改良草地之间都存在着负相关关系,决定系数分别为0.7312和0.7460,随着空间异质性指数的降低平均生物量增加,因此,草地管理中控制空间异质性的产生可提高生产力。
2.4 物种数与生物量之间的关系
从图3可知,天然草地平均物种数与生物量之间存在着正相关关系(R2=0.3945),说明天然草地
的生物量随着物种数的增加而增大,但改良草地物种数与生物量之间存在着负相关关系(R2=0.9245),即随着物种数的增加,生物量逐渐减少。
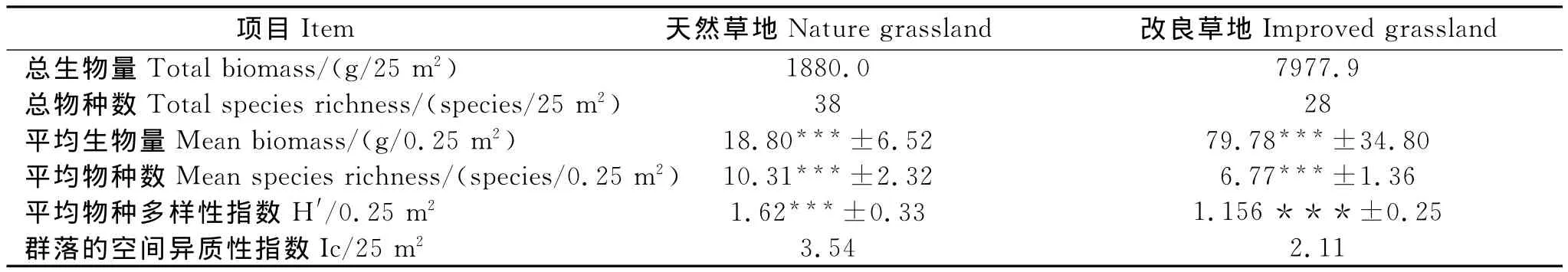
表2 草地群落总体特征Table 2 Community characteristics of grasslands

图1 γ分布模型的吻合性实例Fig.1 Instances fitted to gamma distribution model

图2 平均生物量与空间异质性指数之间的关系Fig.2 Relationship between mean biomass and spatial heterogeneity index
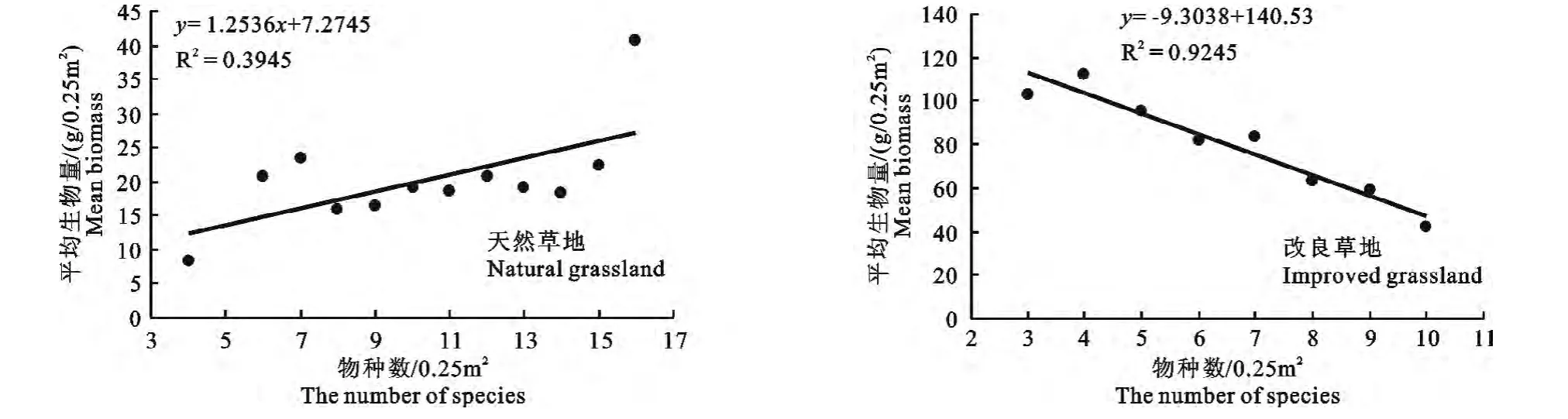
图3 物种数与平均生物量之间的关系Fig.3 Relationship between the number of species and mean biomass
3 讨 论
3.1 γ分布模型解析方法
γ分布模型最早由日本草地生态学专家Shiyomi等[18]应用到半天然草地地上生物量数据的解析中。草地植物群落生物量的频度分布与γ分布模型非常吻合,且经过多次草地植被调查研究证明利用该模型能够准确推测草地生物量的空间分布规律[19-20]。本研究表明,γ分布模型与该地区退化天然草地和补播苜蓿的改良草地地上生物量空间分布也具有很好的吻合性,这与Shiyomi等[18-20]、陈俊等[22]、Tsutsumi等[23-25]、Itano等[26]对日本半 天 然草地的研究结果一致,说明γ分布模型解析法可应用于我国的天然草地生物量空间分布格局的解析。同时利用γ分布模型的解析方法,探明了退化天然草地和补播苜蓿的改良草地植被群落的物种组成、物种多样性、空间分布格局及其与生物量之间的关系,为退化天然草地的改良提供理论依据。
3.2 草地改良对退化草地群落结构特征的影响
天然草地的退化受多种因素影响,补播是退化草地生态系统管理中常用的植被恢复措施,可以提高草地生产力[27-28],本研究中,补播苜蓿后的改良草地平均生物量显著增加(P<0.001),而平均物种数、物种多样性指数极显著的低于退化天然草地(P<0.001),天然草地平均物种数与生物量之间存在着正相关关系,而改良草地物种数与生物量之间存在着负相关关系,与葛庆征等[29]的研究结果相反,其主要原因可能是天然草地的立地条件并没有被现有的植物种完全占据,营养物质也没有完全被利用,所以若有新物种增加就会产生类似于协同的作用,随着物种数的增多其生物量增加[30]。但对于补播苜蓿的改良草地而言,因为苜蓿在群落中占据绝对优势,几乎将有限的生境空间占满,营养物质也得到合理利用,如果再有新物种的侵入,就会与苜蓿之间发生种间竞争,使得生物量减少[31-32]。
3.3 生物量与空间异质性
本研究结果表明,退化天然草地的空间异质性高于改良草地,2种草地植被群落中的所有植物种的空间异质性与它们的平均生物量之间呈现负相关关系(决定系数分别为0.7312和0.746),即随着空间异质性的增大草地植物的生物量逐渐降低,陈俊等[21]对日本半天然草地的研究也得到类似结论。因此,在草地管理与利用中,控制适当的放牧强度、合理刈割,使草地维持在较低的异质性水平,就能提高草地生产力。
[1] 刘兴元,牟月亭.草地生态系统服务功能及其价值评估研究进展[J].草业学报,2012,21(6):286-295.
[2] 刘兴元,龙瑞军,尚占环.青藏高原高寒草地生态系统服务功能的互作机制[J].生态学报,2012,32(24):7 688-7 697.
[3] 董晓玉,傅 华,李旭东,等.放牧与围封对黄土高原典型草原植物生物量及其碳氮磷贮量的影响[J].草业学报,2010,19(2):175-182.
[4] 周宇庭,付 刚,沈振西,等.藏北典型高寒草甸地上生物量的遥感估算模型[J].草业学报,2013,22(1):120-129.
[5] 邓 蕾,上官周平.陕西省天然草地生物量空间分布格局及其影响因素[J].草地学报,2012,20(5):825-835.
[6] Peter A G,Donald L W,Craig C S,et al.Biomass production potential of grasslands in the oak savanna region of Minnesota,USA[J].Bioenergy Research,2013,6(1):131-141.
[7] 邴龙飞,邵全琴,王军邦,等.大样地循环采样的草地生物量空间异质性及误差分析[J].草地学报,2012,20(2):257-267.
[8] 颜增霞,张鹏莉,吕 杰,等.青海湟源冷、暖季放牧草地群落结构特征研究[J].西北农林科技大学:自然科学版,2011,39(8):39-50.
[9] Shiyomi M,Gaborcík J,Koizumi H.Spatial patterns and species diversity of plant communities in sown,oversown,and semi-natural grasslands in Banská Bystrica,Slovakia[J].Grassland Science,2004,50(1):1-8.
[10] Bonham C D,Reich R M,Leader K K.Spatial cross-correlation of bouteloua gracilis with site factors[J].Grassland Science,1995,41:196-201.
[11] 左小安,赵学勇,赵哈林,等.沙地退化植被恢复过程中植被的空间异质性[J].生态环境学报,2010,19(7):1 513-1 518.
[12] 刘金鑫,宫渊波,左 琴,等.山地牧道与植被空间异质性研究评述[J].草业学报,2012,21(1):254-261.
[13] 杨秀云,韩有志,张芸香,等.采伐干扰对华北落叶松细根生物量空间异质性的影响[J].生态学报,2012,32(1):64-73.
[14] 李根柱,王贺新,陈英敏,等.长白山落叶松林下笃斯越桔群落生物量的空间分布[J].生态学杂志,2012,31(6):1 440-1 410.
[15] 戴 诚,康慕谊,纪文瑶,等.内蒙古中部草原地下生物量分配对环境因子的响应关系[J].草地学报,2012,20(2):268-274.
[16] 刘 戎,史兴民,曹继亮.陕西彬县矿区居民人居环境满意度测评[J].西北大学学报:自然科学版,2011,41(4):709-714.
[17] 梁灿盛,殷淑燕,李美荣,等.陕西省彬县近53年来气温及降水变化对白梨物候的影响[J].干旱地区农业研究,2012,30(4):190-194.
[18] Shiyomi M,Akiyama T,Takahashi S.A spatial pattern model of plant biomass in grazing pasture[J].Japan Grassland Science,1983,28:373-382.
[19] Shiyomi M,Okada M,Takahashi S.Spatial pattern changes in aboveground plant biomass in a grazing pasture[J].Ecological Research,1998,13(3):313-322.
[20] Shiyomi M,Akiyama T,Takahashi S.A spatial pattern model of plant biomass in grazing pastureⅡ[J].Journal of Japanese Grassland Science,1984,30:40-48.
[21] 李 强,周冀衡,杨荣生,等.马龙县植烟土壤养分空间变异特征及适宜性评价[J].土壤学报,2011,22(4):950-956.
[22] 陳 俊,堀良通,山村靖夫,等.小面積当り出現回数、被度、バイオマスで測った種数、種構成、空間的不均一性の関係[J].日本草地学会誌,2008,54(2):115-122.
[23] Tsutsumi M,Itano S.Variant of estimation method of aboveground plant biomass in grassland with gamma.Model 1.Use of an electronic capacitance probe[J].Japanese Society of Grassland Science,2005,5:275-279.
[24] Tsutsumi M,Shiyomi M,Wang Y S,et al.Species diversity of grassland vegetation under three different grazing intensities in the Heilongjiang steppe of China[J].Japanese Society of Grassland Science,2003,48(6):510-516.
[25] Tsutsumi M,Itano S,Shiyomi M.Number of samples required for estimating herbaceous biomass[J].Rangeland E-cology,2007,60(4):447-452.
[26] Itano S,Tsutsumi M,Sakanoue S,et al.Variant of estimation method of aboveground plant biomass in grassland with the gamma model.2.Proposal of a modified method with visual observation and tests in Zoysia pastures[J].Japanese Society of Grassland Science,2006,52:53-59.
[27] 冯忠心,周娟娟,王欣荣,等.补播和划破草皮对退化亚高山草甸植被恢复的影响[J].草业科学,2013,30(9):1 313-1 319.
[28] 张永超,牛得草,韩 潼,等.补播对高寒草甸生产力和植物多样性的影响[J].草业学报,2012,21(2):305-309.
[29] 葛庆征,魏 斌,张灵菲,等.草地恢复措施对高寒草甸植物群落的影响[J].草业科学,2012,29(10):1 517-1 520.
[30] 张灵菲,魏 斌,郝 敏,等.物种多样性对豆科牧草功能特征的影响[J].草业科学,2012,29(1):25-29.
[31] 侯扶江,杨中艺.放牧对草地的作用[J].生态学报,2006,26(1):244-264.
[32] 韩德梁,何胜江,陈 超,等.豆禾混播草地群落稳定性的比较[J].生态环境,2008,17(5):1 974-1 979.
