遮阴对大豆幼苗光合和荧光特性的影响
2014-01-02李瑞文涛唐艳萍孙歆夏超
李瑞,文涛,唐艳萍,孙歆,夏超
(四川农业大学农学院,四川 成都611130)
大豆(Glycinemax),古称菽,起源于我国,作为我国主要的粮油兼用型作物,在国民经济和人民生活中占有重要地位。目前,在我国大豆进口量逐年递增的情况下,稳定和发展国内大豆生产,对于保障我国食用大豆需求,保持一定的市场调节能力十分重要。在东北大豆面积饱和的情况下,大力发展南方间套作大豆是振兴我国大豆产业的重要途径之一[1-2]。与高秆作物间套作是南方大豆的主要种植方式[3-4],在间套作模式中,遮阴是制约大豆生长发育、产量和品质形成最主要的因子之一[5-8]。
遮阴对大豆生理形态及光合特性的影响已倍受关注,许多学者从遮阴程度[9-10]、遮阴时期[11-13]和不同耐阴性品种[14-15]等方面进行了一系列研究。王竹等[16]研究发现,田间光照不足导致大豆叶片光合速率和叶绿素a/b值降低,且阴蔽越严重,光合能力越弱。宋艳霞等[17]研究发现,套作遮阴提高了大豆叶片初始荧光(F0)和实际光化学效率(ΦPSⅡ),降低了PSⅡ最大光化学量子产量(Fv/Fm)、光化学荧光淬灭系数(qP)、非光化学荧光淬灭系数(NPQ)。
目前,为适应机械化生产,提高劳动力水平,西南地区玉/豆套作中玉米(Zeamays)采用宽窄行种植模式(窄行种植2行玉米,宽行套作2行大豆),这种玉米宽窄行种植模式下大豆行两侧的光环境有明显差异,邻近玉米行一侧受到高位作物玉米的遮阴,处于弱光环境;邻近大豆行一侧,光照环境充足。目前,关于玉米宽窄行种植模式下,大豆两侧处于不同光环境中的大豆叶片光合特性变化的研究鲜见报道。因此,本研究模拟西南地区玉/豆套作模式,选取强耐阴“南豆12号”和弱耐阴“桂夏3号”2个大豆品种,利用遮阳网盆栽试验,分析遮阴对2个大豆品种幼苗不同时期遮阴及复光后大豆幼苗叶片色素含量、光合特性和叶绿素荧光动力学参数的影响。从新的角度了解套作模式中大豆叶片光合特性变化规律,以期为套作大豆栽培和选育适宜套作模式的高光效大豆等研究提供理论依据。
1 材料与方法
1.1 试验材料
供试大豆品种为强耐阴“南豆12号”和弱耐阴“桂夏3号”,由四川农业大学作物栽培学与耕作学大豆课题组提供。
1.2 试验设计
试验于2013年4月到10月在四川农业大学进行。采用二因素完全随机试验,因素A为品种:南豆12号、桂夏3号;因素B为遮阴:不遮阴(CK)、半侧遮阴30%(B3)、半侧遮阴50%(B5)、半侧遮阴70%(B7)、全遮阴50%(Q5)(半侧遮阴:大豆单侧叶片遮阴处理,另一侧正常光照;全遮阴:对整株大豆遮阴处理)。每个品种5个处理,每个处理5盆,随机排列,每盆定苗5株,3次重复,共150盆。试验设计如图1。
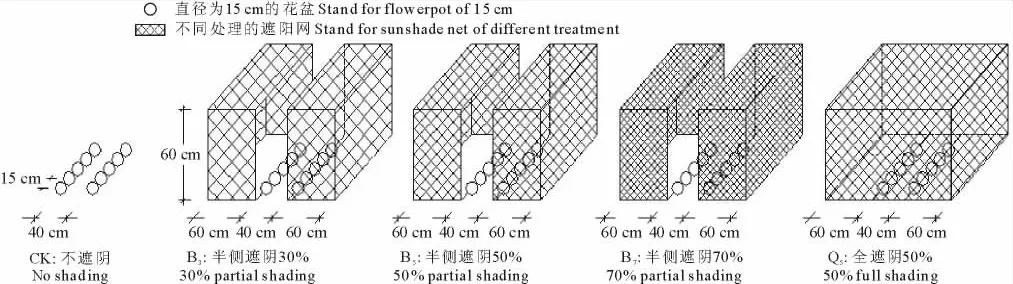
图1 不同遮阴处理设计Fig.1 Different shading treatments designed
试验采用盆栽(12cm×15cm)方式,以腐殖土为基质,挑选饱满均匀的大豆种子,直接播入基质中,各盆栽间行距40cm,窝距15cm。遮阴棚采用半封闭式,呈南北向排列,保证大豆苗全方位遮阴,遮阳网宽度根据田间玉米幅宽设置为60cm,高60cm,各盆栽处理以不互相影响为准。随着大豆的生长逐步抬高遮阴棚,保证模拟的准确性。大豆整个生长期间,正常供应水分,每周浇1次Hoagland营养液,保证大豆生长期间的营养。
1.3 测定项目与方法
于大豆第三复叶期(V3:大豆第三复叶完全展开,第四复叶开始长出时)、第四复叶期(V4)、第五复叶期(V5)和复光7d后(V6)4个时期选取大豆功能叶,动态测定大豆幼苗生长期各处理的叶绿素含量、净光合速率和叶绿素荧光参数。
参照邹琦[18]的方法测定叶绿素a、叶绿素b和叶绿素a/b值。每一处理选取有代表性的大豆幼苗5株,取功能叶剪碎,混合称取0.2g,丙酮乙醇水混合液浸提后用分光光度计在663,645,470nm下分别测定吸光度值并计算叶绿素含量。
选取晴朗天气,在早上9:00-11:00,用美国LI-COR公司生产的LI-6400光合测定系统,使用开放式气路,测定不同遮阴处理下的大豆幼苗叶片的光合参数。CO2浓度为350μmol/(m2·s)左右。选择红蓝光源叶室,设定光量子密度(PAR)为1000μmol/(m2·s)。每处理选取有代表性的大豆幼苗3株,测定其功能叶片中部上表面净光合速率(photosynthetic rate,Pn),取平均值。
用英国Technologica公司生产的CF Imager叶绿素荧光仪测定各处理的叶绿素荧光参数。每一处理选择有代表性的大豆幼苗6株,将功能叶充分暗适应30min后测定初始荧光强度(original fluorescence intensity,F0);然后测定光适应下的PSⅡ最大光化学量子产量(maximum photochemical efficiency,Fv/Fm)、PSⅡ实际光化学效率(actual photochemical efficiency,ΦPSⅡ)、光化学淬灭系数(photochemical quenching,qP)、非光化学淬灭系数(Non-photochemical quenching,NPQ)。
1.4 统计分析
采用Excel 2007整理数据,Sigmaplot 12.5作图,DPS 7.05软件进行方差分析。
2 结果与分析
2.1 遮阴对大豆幼苗叶绿素含量的影响
由表1可知,随着遮阴度的增加,“南豆12号”和“桂夏3号”2个大豆品种的叶绿素a和叶绿素b的含量均呈显著增加趋势。叶绿素a/b值(chla/b)则随遮阴度的增加逐渐降低,“南豆12号”各处理下,高度遮阴Q5与CK差异显著,其余处理间差异总体呈不显著水平;“桂夏3号”各处理下,中度遮阴B7和高度遮阴Q5与CK差异显著,轻度遮阴B3和适度遮阴B5与CK差异不显著。4个时期的变化规律大体一致。分析本试验结果发现,随着遮阴度的增加,2个大豆品种幼苗叶片叶绿素含量的增加主要体现在叶绿素b含量的增长上。2个品种间比较发现,“南豆12号”叶绿素a和叶绿素b的含量显著高于“桂夏3号”,叶绿素a/b值低于“桂夏3号”,但差异不显著,不同处理下“南豆12号”叶绿素b含量的增长幅度高于“桂夏3号”。表明在遮阴下具有较高叶绿素含量和较低叶绿素a/b值的“南豆12号”比“桂夏3号”有更强的光吸收能力,更适应在弱光环境下进行光合作用。

表1 遮阴对不同大豆幼苗叶片叶绿素含量及比值的影响Table 1 Effect of shading on chlorophyll content and proportion of different soybean seedling leaves
2.2 遮阴对大豆幼苗光合速率的影响
由表2可知,遮阴处理下2个大豆品种幼苗叶片的净光合速率(Pn)变化趋势一致,均随遮阴度的增加呈先升高后下降的趋势,B5处理下最高,Q5处理下最低。但2个品种之间也存在差异,“南豆12号”表现为B5>B3>B7>CK>Q5,“桂夏3号”表现为B5>B3>CK>B7>Q5,4个时期的变化趋势一致。4个时期间比较,南豆12号和桂夏3号均在V4期的光合速率显著性最高,表明遮阴对2个大豆品种幼苗在V4期的影响高于其他3个时期。
以不同时期的平均值对2个品种进行比较,“南豆12号”的净光合速率高于“桂夏3号”,中高度遮阴B7和Q5处理下2个品种差异显著,其他各处理下差异不显著。适度遮阴B5处理下“南豆12号”和“桂夏3号”的Pn明显升高,分别比CK高出33.6%和30.6%,升幅差异不大;高度遮阴Q5处理下“南豆12号”和“桂夏3号”的Pn明显下降,分别比CK下降9.9%和29.1%,“南豆12号”降幅较小,“桂夏3号”降幅较大。表明适度的遮阴对2个大豆品种Pn的影响差异不大,高度遮阴对“桂夏3号”Pn的影响明显大于“南豆12号”。
表2 遮阴对不同大豆幼苗光合速率的影响Table 2 Effect of shading on photosynthetic rate of different soybean seedling leaves
2.3 遮阴对大豆幼苗叶绿素荧光动力学参数的影响
F0代表暗适应下叶片的最小荧光,表示PSⅡ反应中心全部开放时的荧光水平,F0降低表示PSⅡ天线色素热耗散增加,F0增加表明PSⅡ反应中心的破坏或可逆失活[19]。由图2可知,不遮阴处理下,2个大豆品种幼苗叶片的F0处于同一水平,但随着遮阴度的增加,“南豆12号”和“桂夏3号”幼苗叶片的F0均呈上升趋势,但2个品种的增加幅度不同。Q5处理下,“桂夏3号”较CK高出24%,“南豆12号”较CK高出18%,可见“桂夏3号”的增加幅度更大。方差分析表明,在B3和B5处理下2个大豆品种的F0均呈缓慢增长,与对照差异不显著,B7和Q5处理下与CK差异达显著水平。4个时期的变化趋势一致。分析可能在B7和Q5处理下,大豆幼苗PSⅡ反应中心可逆失活或被破坏的缘故。
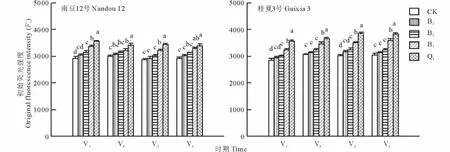
图2 遮阴对大豆幼苗初始荧光强度(F0)的影响Fig.2 Effect of shading on original fluorescence intensity(F0)of soybean seedling leaves
Fv/Fm指PSⅡ原初光能转化效率,正常生理状态下该参数极少变化,绝大多数植物的Fv/Fm在0.75~0.85之间,但逆境条件下该荧光参数会发生变化,可以作为衡量光抑制的指标。ΦPSⅡ指PSⅡ实际光化学效率,常用来表示植物光合作用电子传递的量子产额[20]。图3和图4表明,随着遮阴度的增加,“南豆12号”和“桂夏3号”的Fv/Fm和ΦPSⅡ均呈先升后降趋势,“南豆12号”表现为B5>B7>B3>CK>Q5,“桂夏3号”表现为B5>B3>CK>B7>Q5,4个时期的变化趋势一致。
图3表明,V6期“南豆12号”和“桂夏3号”的Fv/Fm在Q5处理下均小于0.75,其余处理在不同时期的Fv/Fm值均在0.75~0.80之间,与CK差异不显著,表明长时间高度遮阴导致“南豆12号”和“桂夏3号”在复光后受到强光胁迫,Fv/Fm显著下降,品种间受遮阴胁迫后Fv/Fm变化差异不大。图4表明,B7处理下“南豆12号”的ΦPSⅡ高于CK,“桂夏3号”的低于CK,差异均不显著;Q5处理下“南豆12号”和 “桂夏3号”的ΦPSⅡ均低于CK,但“桂夏3号”下降幅度大于“南豆12号”。说明适度遮阴对2个品种ΦPSⅡ的影响差异不大,但中高度遮阴则显著降低“桂夏3号”的PSⅡ实际光化学效率。

图3 遮阴对大豆幼苗最大光化学效率(Fv/ Fm)的影响Fig.3 Effect of shading on maximum photochemical efficiency(Fv/ Fm)of soybean seedling leaves
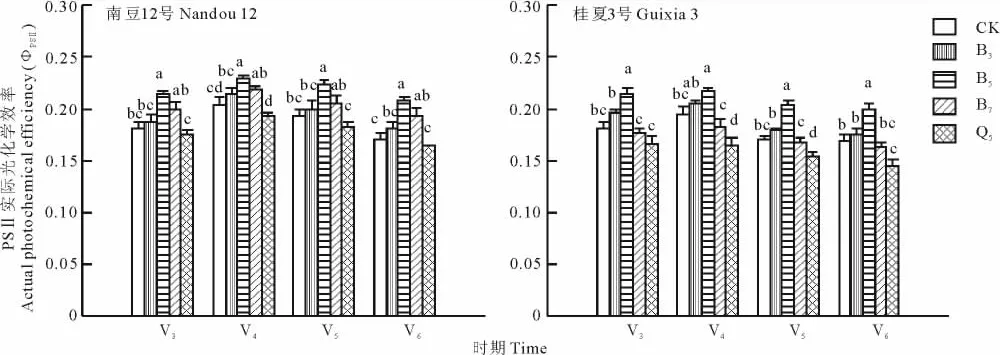
图4 遮阴对大豆幼苗PSⅡ实际光化学效率(ΦPSⅡ)的影响Fig.4 Effect of shading on actual photochemical efficiency(ΦPSⅡ )of soybean seedling leaves
荧光淬灭是植物体内光合量子效率调节的一个重要方面,分为光化学淬灭(qP)和非光化学淬灭(NPQ)两类。qP反映的是PSⅡ天线色素吸收的光能用于光化学传递的份额,qP愈大,PSⅡ的电子传递活性愈大;NPQ反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的光能部分[21]。图5可知,随着遮阴程度的增加,2个大豆品种幼苗叶片的qP呈先升后降趋势,“南豆12号”表现为B5>B7>B3>CK>Q5,“桂夏3号”表现为B5>B3>CK>B7>Q5,说明中高度遮阴对“桂夏3号”qP的影响大于“南豆12号”,不同时期变化趋势一致,V4期的qP高于其他3个时期。试验表明适度的遮阴有利于提高PSⅡ反应中心开放部分的比例,将更多的光能用于推动光合电子传递,从而提高光能潜力的利用。
随着遮阴度的增加,“南豆12号”的NPQ表现为先降后升趋势,B3、B5处理下低于CK,差异不显著,B7、Q5处理下显著高于CK(图6)。“桂夏3号”在B3处理下的NPQ显著低于CK,其余处理则显著高于CK,Q5处理下的NPQ不再增加,和B7处理维持在一定的水平,差异不显著,表明中高度遮阴下“桂夏3号”用于过剩光能耗散的比例相差不大,可能是高度遮阴下“桂夏3号”PSⅡ反应中心光能过剩,来不及以热能形式耗散。不同时期的变化趋势一致,2个大豆品种在V6期的NPQ均高于其他3个时期,说明复光后大豆过剩光能急剧上升,大豆通过增加热耗散保护其光合机构免受伤害。

图5 遮阴对大豆幼苗叶绿素荧光光化学淬灭(qP)的影响Fig.5 Effect of shading on photochemical quenching of chlorophyll fluorescence(qP)of soybean seedling leaves

图6 遮阴对大豆幼苗叶绿素荧光非光化学淬灭(NPQ)的影响Fig.6 Effect of shading on non-photochemical quenching of chlorophyll fluorescence(NPQ)of soybean seedling leaves
3 讨论和结论
遮阴是西南地区大豆产量的主要限制因子,合适的种植密度可以改善群体结构,提高光能利用率,使品种的优良特性得到最大限度的发挥[22]。而光照不足则会限制植物的光合作用,导致碳获取的减少,使生长受阻[23]。因此植物光合作用的运转状况决定着其生产力水平,其光合性能可以通过叶绿素含量的比例变化、光合速率得到衡量[24],而叶绿素荧光作为光合作用机理研究的有力探针,可以更直接地反映不同光照条件下光合作用反应中心的开放程度及植物热耗散等情况,是一种反应灵敏、操作简便且对植物无伤害的方法。
遮阴对光合作用的影响,因作物品种、生育期和遮阴时间的不同,结果有差异,许多研究表明,遮阴通常降低单叶的净光合速率[25]。但本研究结果却发现大豆幼苗光合速率随着遮阴度的增加呈先升后降趋势,这与谭春燕等[26]的研究结果一致。B5处理下大豆幼苗光合速率显著高于CK,分析原因是因为南豆12号和桂夏3号均为耐阴性品种,为满足生存需要,在生长发育过程中对环境中的光信息能不断作出可塑性变化,形成了耐阴机制,通过自我机体的调节,促使大豆幼苗的光暗反应适应环境中光强和光质的改变,最终使植株适应了弱光环境。但中高度遮阴B7和Q5处理下“桂夏3号”Pn大幅度下降,显著低于CK,可能是强遮阴限制了叶绿体光能潜力的发挥或对大豆PSⅡ光系统中心造成破坏或可逆失活的缘故。
植物在一定范围内对光环境有很强的自我适应与调节能力,一些植物在弱光下会通过合成大量的叶绿素,去捕获更多的光能,但不同光反应类型的品种间存在差异[27]。本试验研究结果与梁镇林等[28]的研究结果一致,即随着遮阴度的增加,大豆幼苗叶片的叶绿素含量均显著增加,主要体现在叶绿素b含量的增长上,且“南豆12号”的增加幅度大于“桂夏3号”。这是由于遮阴在降低光强度的同时也改变了光谱组成,挡住了直射光源从而导致红光比例降低,蓝光更容易散射而大量聚集在遮阴处,使蓝光比例增高。且叶绿素a有较宽的红光区吸收带,叶绿素b有较宽的蓝光区吸收带[29]。因此耐阴性品种主要通过增加叶绿素b含量的方式来吸收更多的光能,以适应弱光环境,试验表明在弱光环境“南豆12号”比“桂夏3号”有更强的光吸收能力。
叶绿素荧光动力学技术在测定叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等方面具有独特的作用,与“表观性”的气体交换指标相比,叶绿素荧光参数更具有反映“内在性”的特点[20]。通过本试验遮阴引起的“南豆12号”和“桂夏3号”大豆幼苗功能叶F0、Fv/Fm、ΦPSⅡ、qP和NPQ等荧光参数的变化,结果表明适度遮阴处理下2个品种幼苗叶片的变化趋势一致,但中高度遮阴处理下2个品种间存在差异。适度的遮阴下大豆幼苗叶片的F0、Fv/Fm、ΦPSⅡ和qP增加,而NPQ下降,表明“南豆12号”和“桂夏3号”大豆幼苗叶片在适度遮阴条件下,通过降低热耗散,提高光化学反应效率,进而维持较高的能量代谢水平[30],以适应光环境的变化。中高度遮阴下,大豆幼苗叶片的Fv/Fm、ΦPSⅡ和qP降低,而F0和NPQ升高,桂夏3号”变化幅度明显大于“南豆12号”,且Q5处理下“桂夏3号”的NPQ与B7处理维持在一定的水平不再增加,表明高度遮阴导致“南豆12号”和“桂夏3号”大豆幼苗叶片PSⅡ反应中心的开放比例降低,天线色素捕获的光能流向光化学的部分减少,光合电子传递速率下降,使累积在PSⅡ反应中心的光能过剩,“南豆12号”通过提高NPQ及时耗散了过剩的光能,保护了光合机构免遭破坏,而“桂夏3号”在Q5处理下不能及时耗散过剩光能,受到光氧化的伤害,导致其PSⅡ反应中心受到损伤,是光合速率下降的主要原因,这也许是2个品种在耐阴性上表现出差异的原因之一。
本研究还发现在50%遮阴下,全遮阴处理的2个大豆品种光合性能的增降幅度明显大于半侧遮阴处理。这与何维明和董鸣[31]对旱柳局部遮阴和全遮阴处理下的研究结果一致,即部分遮阴能够通过改变叶片光合性能来提高阳生枝的光合产物积累,降低阴生枝的光合产物积累,从而将更多的资源分配到强光环境中,使整个植株获得更多的光合产物而有利于其生长。
综上所述,在玉米窄行大豆宽行种植模式下,半遮阴50%左右的遮阴度较适于耐阴大豆的营养生长,而常规的全遮阴50%则对大豆的营养生长造成一定的损失,且长期高遮阴下的大豆幼苗在复光后会受到光胁迫。“南豆12号”在遮阴胁迫下的各参数平均值均高于“桂夏3号”,且变化幅度小于“桂夏3号”,具有更强的耐阴性。适度遮阴后,大豆幼苗通过改变色素含量及组分比例,吸收更多的光能,来提高PSⅡ反应中心的开放比例,推动光合电子的传递,将吸收的能量用来增加光化学反应,进而提高光能潜力的利用以适应弱光环境。长期过度遮阴后,“南豆12号”可以及时清除过剩光能,保护其光合机构免受破坏,而“桂夏3号”不能及时清除过剩光能,造成大豆幼苗光合机构的破坏,是光合速率下降的主要原因。
[1]常汝镇,韩天富.关于发展南方间套作大豆生产的建议[J].大豆科技,2008,(4):7.
[2]杨文钰,雍太文,任万军,等.发展套作大豆,振兴大豆产业[J].大豆科学,2008,(1):1-7.
[3]吴迅,张明荣,吴海英,等.我国南方耐荫大豆的现状及前景[J].黑龙江农业科学,2009,(5):148-150.
[4]陈远学,陈晓辉,唐义琴,等.不同氮用量下小麦/玉米/大豆周年体系的干物质积累和产量变化[J].草业学报,2014,23(1):73-83.
[5]陈怀珠,孙祖东,杨守臻,等.荫蔽对大豆主要性状的影响及大豆耐荫性鉴定方法研究初报[J].中国油料作物学报,2003,25(4):78-82.
[6]黄其椿,李初英,吴建明,等.不同遮光处理对菜用大豆产量的影响[J].大豆科学,2012,31(1):81-84.
[7]吴其林,王竹,杨文钰.苗期遮阴对大豆茎秆形态和物质积累的影响[J].大豆科学,2007,26(6):868-872.
[8]刘庆超,刘庆华,马宗骧,等.三桠乌药耐阴性研究[J].草业学报,2013,22(6):93-99.
[9]王秀领,闫旭寿,徐玉鹏,等.玉米-大豆间作复合体系光合特性研究[J].河北农业科学,2012,16(4):33-35,59.
[10]林绍森,唐永金.玉米密度、行距和穴距对间作大豆光合速率的效应分析[J].大豆科学,2007,26(2):149-153.
[11]王瑞,王宏富,陈新霞,等.玉米大豆不同间作模式下大豆鼓粒期的光学特性[J].东北农业大学学报,2012,43(4):16-20.
[12]王一,杨文钰,张霞,等.不同生育时期遮阴对大豆形态性状和产量的影响[J].作物学报,2013,39(10):1871-1879.
[13]伍晓燕,王竹,张含彬,等.玉/豆套作对大豆开花后光合生产的影响[J].作物杂志,2006,(3):30-33.
[14]梁慕勤,潘世元,梁镇林.大豆耐荫性的研究Ⅱ:不同生态区的品种、植株形态和籽粒特性与大豆耐荫性的关系[J].贵州农业科学,1986,(3):5-8.
[15]于晓波,张明荣,吴海英,等.净套作下不同耐荫性大豆品种农艺性状及产量分布的研究[J].大豆科学,2012,31(5):757-761.
[16]王竹,杨文钰,吴其林.玉/豆套作荫蔽对大豆光合特性与产量的影响[J].作物学报,2007,33(9):1502-1507.
[17]宋艳霞,杨文钰,李卓玺,等.不同大豆品种幼苗叶片光合及叶绿素荧光特性对套作遮阴的响应[J].中国油料作物学报,2009,31(4):474-479.
[18]邹琦.现代植物生理学实验指南[M].北京:中国农业出版社,2000:72-75.
[19]许大全,张玉忠,张荣铣.植物光合作用的光抑制[J].植物生理学通讯,1992,28(4):237-243.
[20]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[21]郭春芳,孙云,唐玉海,等.水分胁迫对茶树叶片叶绿素荧光特性的影响[J].中国生态农业学报,2009,17(3):560-564.
[22]詹杰,罗旭辉,苏小珍,等.不同留株密度对圆叶决明生产性能及光合特性的影响[J].草业学报,2011,20(5):66-71.
[23]Hu Y B,Sun G Y,Wang X C.Induction characteristics and response of photosynthetic quantum conversion to changes in irradiance in mulberry plants[J].Journal of Plant Physiology,2006,164:959-968.
[24]闫瑞瑞,卫智军,运向军,等.放牧制度对短花针茅荒漠草原主要植物种光合特性日变化影响的研究[J].草业学报,2009,18(5):160-167.
[25]刘贤赵,康绍忠,周吉福.遮阴对作物生长影响的研究进展[J].干旱地区农业研究,2001,19(4):65-73.
[26]谭春燕,刘作易,朱星陶,等.不同密度间作大豆的光合生理响应[J].贵州农业科学,2012,40(8):49-52.
[27]许岳飞,金晶炜,陈浩,等.草坪植物耐弱光机理研究进展[J].草地学报,2011,19(6):1064-1069.
[28]梁镇林,梁慕勤,潘世元,等.大豆耐阴性研究Ⅹ:不同耐阴性大豆叶片叶绿素含量和比叶重研究[J].贵州农学院学报,1992,11(2):16-22.
[29]孙小玲,许岳飞,马鲁沂,等.植株叶片的光合色素构成对遮阴的响应[J].植物生态学报,2010,34(8):989-999.
[30]张哲,杨姝,杜桂娟,等.遮阴对三种豆科牧草光合特性和叶绿素荧光参数的影响[J].草业学报,2013,22(5):212-219.
[31]何维明,董鸣.毛乌素沙地旱柳生长和生理特征对遮阴的反应[J].应用生态学报,2003,14(2):175-178.
